HAL Id: tel-00154576
https://tel.archives-ouvertes.fr/tel-00154576
Submitted on 14 Jun 2007HAL is a multi-disciplinary open access archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
DE PEPTIDES N-FORMYLES
Emilie Huet Moulard
To cite this version:
Emilie Huet Moulard. ETUDE DE LA REGULATION DES RECEPTEURS DE PEPTIDES N-FORMYLES. Biologie cellulaire. Université Joseph-Fourier - Grenoble I, 2007. Français. �tel-00154576�
présentée par
Emilie HUET MOULARD
pour obtenir le grade de
DOCTEUR
DE L’UNIVERSITE JOSEPH FOURIER-GRENOBLE I
Discipline : Biologie Cellulaire soutenue le 5 juin 2007
ETUDE DE LA REGULATION
DES RECEPTEURS DE PEPTIDES N-FORMYLES
Directrice de thèse : Dr. Marie-Josèphe RABIET
JURY
Pr. Françoise Morel Présidente
Pr. Oliver Nüße Rapporteur
Dr. Ralf Jockers Rapporteur
Dr. Eric Reiter Examinateur
Dr. Marie-Josèphe Rabiet Examinateur
Thèse préparée au Laboratoire de Biochimie et Biophysique des Systèmes Intégrés iRTSV / CEA-Grenoble
RESUME
Les cellules phagocytaires (neutrophiles, monocytes et macrophages) constituent la première ligne de défense contre les pathogènes. Leur migration dirigée vers le site infectieux et la libération de leur arsenal antimicrobien et cytotoxique sont l'aboutissement de voies de signalisation intracellulaires sollicitées par la stimulation de récepteurs appelés récepteurs de chimioattractants. Ces récepteurs appartiennent à la famille des récepteurs couplés aux protéines G hétérotrimériques (RCPGs). Après fixation du ligand et transmission du signal par la protéine G hétérotrimérique, les RCPGs sont phosphorylés et interagissent avec les β-arrestines, protéines d’échafaudage qui concourent au processus d’internalisation des récepteurs par la voie des puits mantelés de clathrine. Plusieurs exemples récents suggèrent qu’une signalisation indépendante des protéines G pourrait également être transmise par les β-arrestines et font émerger le concept de macro-complexes de signalisation capables d’assurer la spécificité des réponses cellulaires.
Le travail présenté dans ce mémoire concerne les récepteurs de la famille FPR (Formyl Peptide Receptor) et plus spécialement le récepteur FPRL1 (FPR-like 1).
Des peptides N-formylés dérivant de protéines du pathogène Listeria monocytogenes ou de protéines mitochondriales humaines ont été identifiés comme nouveaux agonistes de ces récepteurs. Les sélectivités relatives des différents récepteurs pour les peptides bactériens ou mitochondriaux assignent un rôle prédominant pour le récepteur FPR dans la défense antibactérienne et suggèrent un rôle des récepteurs de la famille FPR dans les phénomènes inflammatoires associés aux dommages tissulaires et aux maladies dégénératives.
La régulation des récepteurs dépend essentiellement de leur phosphorylation et de leur internalisation. Dans cette étude, il a été montré que le récepteur FPRL1 était phosphorylé sur des résidus sérines et thréonines. Pour cartographier les sites de phosphorylation, des mutants de remplacement Ser /Thr en alanine ont été construits par mutagenèse dirigée de la région C-terminale du récepteur. L’étude de leur phoshorylation a permis d’identifier les sites majeurs de phosphorylation. Il a été montré que les β-arrestines interagissent avec le récepteur FPRL1 phosphorylé et qu’elles sont indispensables à son internalisation. Ceci différencie le récepteur FPRL1 du récepteur FPR qui peut être internalisé même en l’absence des β-arrestines.
Par ailleurs, il a été montré par diverses approches (surexpression de formes sauvages ou de "dominants négatifs" des β-arrestines, utilisation de cellules déficientes en β-arrestines et inhibition par la toxine pertusssique de la signalisation par les protéines G de type Gi) que l’activation rapide et transitoire des MAP kinases ERK1/2, enclenchée par la stimulation du récepteur FPRL1, est majoritairement dépendante de la protéine G héterotrimérique et qu’il n’y pas de signalisation transmise par les β-arrestines. Cette propriété, que le récepteur FPRL1 partage avec le récepteur FPR, pourrait être une particularité des récepteurs de chimioattractants qui sont essentiellement couplés à des protéines G de type Gi.
Enfin, une analyse protéomique des complexes multi-protéiques bâtis autour du couple FPRL1/β-arrestine a été menée. Après purification de la β-arrestine et des protéines associées par la méthode TAP (Tandem Affinity Purification), l’identification par spectrométrie de masse des protéines présentes a permis d’identifier un nouveau partenaire des β-arrestines qui interagirait avec celles-ci après stimulation du récepteur FPRL1. Il s’agit du complexe adaptateur AP3. Son homologue AP2 est impliqué dans l’internalisation par les puits mantelés de clathrine, mais peu de données existent concernant AP3. Une implication dans le trafic endocytaire ou post-endocytaire du récepteur peut être envisagée.
TABLE DES MATIERES
INTRODUCTIONA/ Les phagocytes, première ligne de défense de l’organisme contre les pathogènes... 1
I- Les phagocytes ... 1
II- Les fonctions cellulaires du neutrophile... 3
II-1 Le chimiotactisme ou migration cellulaire dirigée... 3
II-2 La phagocytose... 5
II-3 La mobilisation des granules... 5
II-4 L’explosion respiratoire, la NADPH oxydase... 7
B/ Les récepteurs de chimioattractants... 9
I- Les récepteurs couplés aux protéines G hétérotrimériques ... 10
II- Les récepteurs chimiotactiques ... 12
II-1 Description générale des récepteurs de la famille FPR... 12
II-2 Ligands des récepteurs de la famille FPR ... 13
II-3 Localisation cellulaire et tissulaire des récepteurs de la famille FPR ... 15
II-4 Signalisation intracellulaire en aval des récepteurs de chimioattractants ... 16
II-5 Régulation de l’activité des récepteurs de chimioattractants ... 19
C/ Les β-arrestines ... 24
I- Historique ... 24
I-1 L’arrestine visuelle ... 24
I-2 Découverte des β-arrestines... 24
II- Description des β-arrestines... 25
II-1 Description générale... 25
II-2 Caractéristiques des deux isoformes de β-arrestine ... 27
II-3 Données structurales sur les β-arrestines ... 29
II-4 Etat oligomérique des β-arrestines... 31
III- Fonctions des β-arrestines... 32
III-1 Découplage du récepteur à la protéine G ... 33
III-2 Implication dans l’internalisation des récepteurs... 36
III-3 Implication dans la signalisation, protéines d’échafaudage... 40
D/ But du travail... 43
MATERIELS ET METHODES A/ Biologie Cellulaire et Biochimie ... 44
I- Lignées et cultures cellulaires... 44
II- Transfection des cellules ... 45
II-1 Transfection transitoire ... 45
II-2 Transfection stable ... 46
III- Analyses menées sur cellules vivantes ... 46
III-1 Mobilisation du calcium intracellulaire ... 46
III-2 Test de chimiotactisme ... 47
III-3 Dosage de la production d’ions superoxyde ... 47
III-4 Cytométrie de flux ... 48
IV- Techniques utilisant l’immunoreconnaissance par des anticorps ... 48
IV-2 Immunoprécipitation... 50
IV-3 Immunodétection après électrophorèse et électrotransfert ... 50
V- Préparation d’extraits cellulaires... 51
V-1 Cinétique d’activation des MAP-kinases ERK1/2... 51
V-2 Purification de complexes par affinité en tandem... 52
V-3 Préparation membrane/cytosol... 53
VI- Manipulations effectuées avec du matériel radioactif ... 53
VI-1 Phosphorylation du récepteur après marquage métabolique avec l’acide orthophosphorique [32P] ... 53
VI-2 Analyse des acides aminés phosphorylés ... 53
VI-3 Iodation du peptide WKYMVm ... 54
VI-4 Liaison de ligand radioactif ... 54
VI-5 Séquestration de ligand radioactif ... 55
B/ Biologie Moléculaire... 55
I- Méthodologies ... 55
I-1 Préparation d’ADN plasmidique ... 55
I-2 Réaction en chaîne par polymérase, PCR... 56
I-3 Purification des fragments d’ADN ... 56
I-4 Mutagenèse dirigée (Kit QuickChange® de Stratagene) ... 56
I-5 Hybridation des oligonucléotides ... 57
I-6 Ligation et transformation par choc thermique ... 57
II- Matériels... 57
III- Constructions plasmidiques ... 59
III-1 Vecteurs d’expression de la β-arrestine 1 mutée sur le site polyproline ... 59
III-2 Vecteurs d’expression du récepteur FPRL1 et de ses mutants de phosphorylation ... 59
III-3 Vecteurs d’expression basés sur le système TAP ... 61
RESULTATS ET DISCUSSIONS A/ Identification de nouveaux agonistes des récepteurs de la famille FPR... 65
I- Identification de nouveaux peptides N-formylés induisant la signalisation intracellulaire par stimulation des récepteurs de la famille FPR ... 67
I-1 Mobilisation du calcium intracellulaire ... 67
I-2 Signalisation vers les MAP kinases... 70
II- Capacité des agonistes nouvellement identifiés à induire des réponses cellulaires telles que le chimiotactisme et la génération d’ions superoxyde... 72
II-1 Capacité du peptide d’origine mitochondriale f-MMYALF à induire le chimiotactisme des cellules HL-60 exprimant FPRL1 ou FPRL2 ... 72
II-2 Capacité des agonistes f-MMYALF et f-MIVIL à entraîner la génération d’ions superoxyde par la NADPH oxydase dans les cellules HL-60 différenciées ... 73
III- Discussion et conclusions ... 75
B/ Rôle des β-arrestines dans les voies de signalisation initiées par les récepteurs de chimioattractants... 78
I- Etude du rôle de l’interaction entre la β-arrestine 1 et la tyrosine kinase c-Src dans les cellules myéloïdes HL-60... 79
I-1 Etablissement d’une lignée HL60 exprimant de façon inductible la β-arrestine 1 sauvage ou mutée sur les motifs impliqués dans son interaction avec la tyrosine kinase c-Src ... 79
I-2 Effet de l’expression du double mutant P91G-P121E de la β-arrestine 1 sur la production d’ions superoxyde ... 82
II- Etude du rôle des β-arrestines dans la signalisation vers les MAP kinases suite à la
stimulation du récepteur FPRL1 ... 84
II-1 Effet de la surexpression des β-arrestines... 84
II-2 Effet de l’absence des β-arrestines... 86
II-3 Effet de l’inhibition de la voie dépendante des protéines Gi ... 88
II-4 Discussions... 91
C/ Etude de la régulation des récepteurs des peptides N-formylés : phosphorylation et internalisation ... 92
I- Phosphorylation ... 92
I-1 Etude des sites de phosphorylation du récepteur FPRL1 ... 92
I-2 Etude de la phosphorylation de chimères des récepteurs FPRL1 et FPRL2 ... 98
II- Etude du rôle des β-arrestines dans l’internalisation du récepteur FPRL1... 101
II-1 Etude de l’association des β-arrestines avec le récepteur FPRL1... 101
II-2 Etude du rôle des β-arrestines dans l’internalisation du récepteur FPRL1... 102
II-3 Discussion ... 104
D/ Etude protéomique globale des complexes formés autour du couple "récepteur FPRL1 / β-arrestine" ... 106
I- Purification des complexes par la méthode TAP (Tandem Affinity Purification) ... 106
II- Vecteurs d’expression ... 109
III- La purification par affinité en tandem appliquée aux β-arrestines ... 110
III-1 Mise au point de la purification par la méthode TAP ... 110
III-2 Etablissement des lignées cellulaires stables exprimant les β-arrestines fusionnées à l’étiquette TAP en vue de l’étude protéomique ... 111
III-3 Résultats de l’étude protéomique menée sur les β-arrestines étiquetées TAP... 113
III-4 Etude de l’interaction entre les β-arrestines et AP3... 120
III-5 Discussions et perspectives sur l’étude protéomique menée sur les β-arrestines... 125
IV- La méthode TAP appliquée au récepteur FPRL1 ... 127
IV-1 Etablissement de lignées cellulaires stables exprimant FPRL1-TAP... 127
IV-2 Résultats... 128
IV-3 Perspectives ... 132
DISCUSSION GENERALE ... 135
BIBLIOGRAPHIE ... 138
ANNEXES Annexe A : Inhibition sélective et inductible de l’expression d’une protéine ... 156
Annexe B : Résultats complets des analyses protéomiques……….………162 Annexe C : Article Rabiet et al. (2005) Human mitochondria-derived N-formylated peptides are novel agonists equally active on FPR and FPRL1, while Listeria monocytogenes-derived peptides preferentially activate FPR. Eur. J. Immunol., 35, 2486-2495
Annexe D : Revue Rabiet et al. (2007) The N-formyl peptide receptors and the anaphylatoxin C5a receptors : an overview. Biochimie, In Press
Annexe E : Article Huet et al. (2007) The role of beta-arrestins in the formyl peptide receptor-like 1 internalization and signaling, Cellular Signaling, In Press
TABLE DES FIGURES
Figure 1 : Polynucléaire neutrophile entouré de globules rouges après coloration au
May-Grünwald-Giesma ... 1
Figure 2 : Schématisation de l’hématopoïèse... 2
Figure 3 : Migration du neutrophile vers le site inflammatoire ... 3
Figure 4 : Polarisation d’un neutrophile... 4
Figure 5 : Phagocytose de candida albicans par un polynucléaire neutrophile... 5
Figure 6 : Schématisation de la dégranulation du neutrophile ... 6
Figure 7 : Schématisation de l’assemblage du complexe NADPH oxydase phagocytaire ... 7
Figure 8 : Schématisation de la notion de récepteur membranaire spécifique... 9
Figure 9 : Représentation schématique d’un RCPG dans la membrane plasmique ... 10
Figure 10 : Cycle d’activation des protéines G hétérotrimériques... 11
Figure 11 : Représentation schématique des voies de signalisation enclenchées par les récepteurs de chimioattractants ... 17
Figure 12 : Représentation schématique du trafic régulé des récepteurs de chimioattractants 20 Figure 13 : Alignement des domaines C-terminaux de récepteurs de chimioattractants ... 21
Figure 14 : Alignement de protéines de la famille "arrestine" ... 26
Figure 15 : Identité des membres de la famille des arrestines humaines ... 27
Figure 16 : Alignement des deux isoformes de β-arrestines humaines... 27
Figure 17 : Structure de la β-arrestine 1 bovine, diagramme en ruban... 29
Figure 18 : Carte de densité électronique du corps polaire de la β-arrestine 1 bovine ... 29
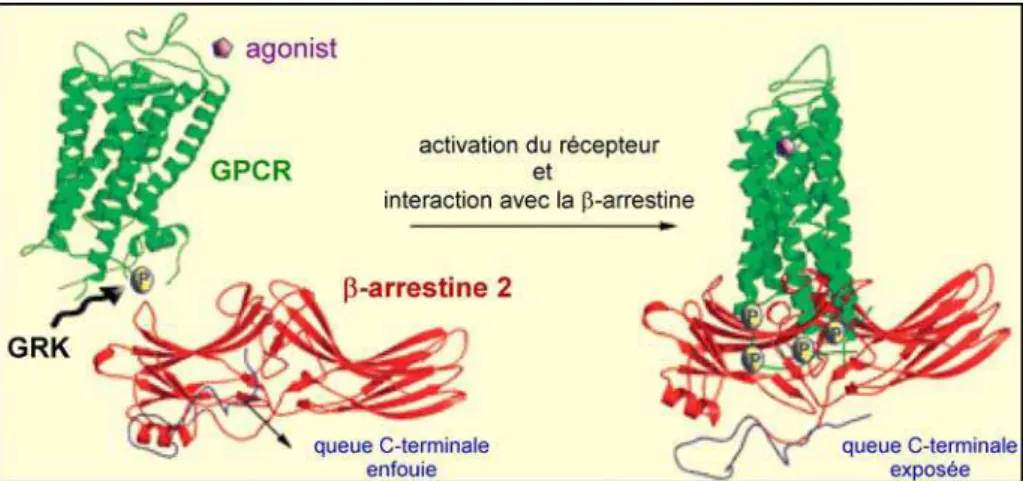
Figure 19 : Modèle structural des changements conformationnels de la β-arrestine 2 suite à l’activation par un RCPG phosphorylé ... 31
Figure 20 : Homologies entre les arrestines et les protéines G ... 34
Figure 21 : Structure de la β-arrestine 2 présentant quelques uns de ses sites d’interaction ... 37
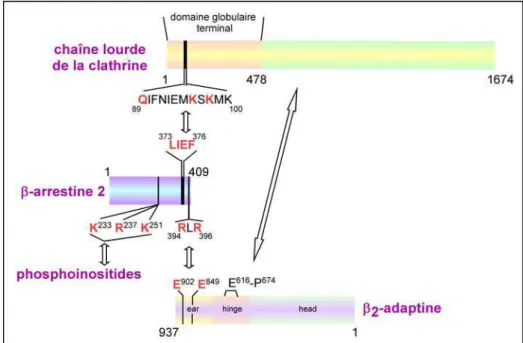
Figure 22 : Schématisation des interactions protéiques impliquant la β-arrestine et des composants de la machinerie d’endocytose ... 39
Figure 23 : Stratégie de biologie moléculaire pour l’obtention des mutants de FPRL1 ... 60
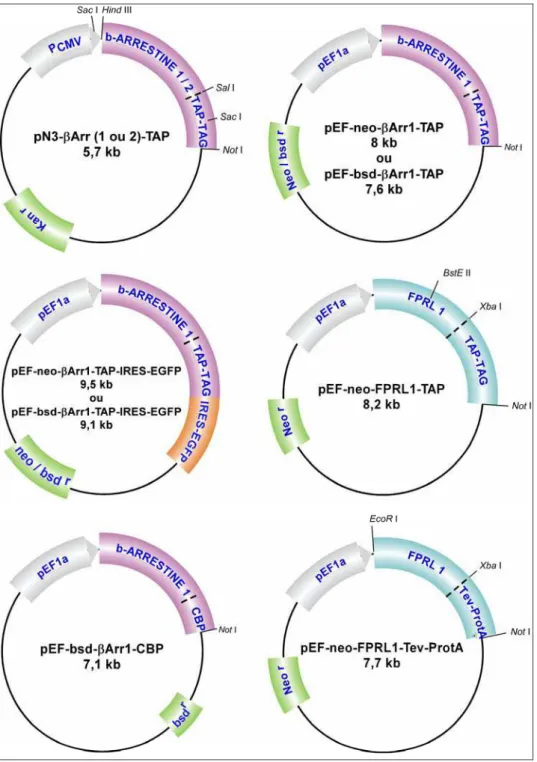
Figure 24 : Schématisation des vecteurs d’expression basés sur la méthode TAP ... 63
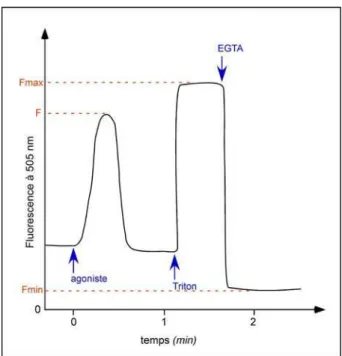
Figure 25 : Suivi de fluorescence type lors d’un test de mobilisation calcique au Fura2 ... 68
Figure 26 : Courbes dose-réponse de la mobilisation de calcium intracellulaire dans les cellules HL-60 exprimant FPR, FPRL1 ou FPRL2... 69
Figure 27 : Activation des MAP kinases dans les cellules HL-60 exprimant FPR, FPRL1 ou FPRL2 ... 71
Figure 28 : Chambre de chimiotactisme ... 72
Figure 29 : Activité chimiotactique du peptide mitochondrial f-MMYALF dans les cellules HL-60 non différenciées exprimant FPRL1 ou FPRL2 ... 73
Figure 30 : Activité NADPH oxydase dans des cellules HL-60 différenciées ... 74
Figure 31 : Schématisation d’une voie de signalisation initiée par les chimioattractants, implication éventuelle des β-arrestines... 78
Figure 32 : Système inductible régulé par la tétracycline ... 80
Figure 33 : Plasmides permettant l’expression de la protéine de fusion β-arrestine 1-EGFP . 81 Figure 34 : Analyse de l’expression et de la répression des protéines de fusion par cytométrie de flux... 81
Figure 35 : Analyse de la production d’ions superoxyde dans les cellules HL-60-tTA
exprimant ou non la protéine de fusion β-arrestine 1-EGFP ou le mutant ... 82
Figure 36 : Cinétique d’activation des MAP kinases dans les cellules HEK-293-FPRL1 surexprimant ou non les β-arrestines ... 85
Figure 37 : Effet de petits ARN interférents ciblant les β-arrestines sur la cinétique d’activation des MAP kinases dans les cellules HEK-293-FPRL1... 86
Figure 38 : Caractérisation des lignées MEF-FPRL1 et MEF-βarr1&2-/--FPRL1... 87
Figure 39 : Cinétique d’activation des MAP kinases dans les cellules MEF-FPRL1 déficientes ou non en β-arrestine... 88
Figure 40 : Effet de la toxine pertussique sur l’activation des MAP kinases ERK1/2 dans les cellules HEK-293-FPRL1 ... 89
Figure 41 : Effet de la toxine pertussique sur l’activation des MAP kinases ERK1/2 dans les cellules MEF-FPRL1 déficientes ou non en β-arrestine ... 90
Figure 42 : Alignement des domaines C-terminaux des récepteurs FPR et FPRL1 ... 93
Figure 43 : Cartographie des acides aminés phosphorylés du récepteur FPRL1 ... 93
Figure 44 : Schématisation du récepteur FPRL1 avec ses sites potentiels de phosphorylation regroupés en clusters ... 94
Figure 45 : Phosphorylation des mutants du récepteur FPRL1... 96
Figure 46 : Schématisation des chimères entre les récepteurs FPRL1 et FPRL2 ... 98
Figure 47 : Séquences des queues cytoplasmiques de FPRL1 et FPRL2 ... 99
Figure 48 : Phosphorylation basale des récepteurs FPRL1 et 2 et de leurs chimères ... 100
Figure 49 : Co-localisation de la β-arrestine 2-EGFP et du récepteur FPRL1 ... 102
Figure 50 : Internalisation du récepteur FPRL1 révélée par immunofluorescence dans les cellules MEF déficientes ou non en β-arrestine... 103
Figure 51 : Séquestration de ligand radioactif dans les cellules MEF déficientes ou non en β-arrestine ... 103
Figure 52 : Schématisation de l’étiquetage TAP en C-terminal... 107
Figure 53 : Schéma de la méthode TAP de purification et d’analyse de complexes protéiques ... 108
Figure 54 : Séquence du TAP-TAG du vecteur pZome1C®... 109
Figure 55 : Les étapes de la purification TAP vues par immunodétection de la β-arrestine 1 étiquetée ... 110
Figure 56 : Expression stable des β-arrestines étiquetées dans le modèle cellulaire HEK-293 ... 112
Figure 57 : Translocation des β-arrestines 2 étiquetée ou endogène à la membrane après stimulation... 113
Figure 58 : Contenu protéique des différentes étapes de la purification TAP de la β-arrestine 1 étiquetée TAP dans le clone HEK-293-βArr1-TAP ... 114
Figure 59 : Représentation graphique des résultats des analyses protéomiques menées sur les complexes formés autour des β-arrestines ... 117
Figure 60 : Co-immunoprécipitation de l’adaptine β2 par la β-arrestine 2-TAP ... 120
Figure 61 : Schématisation de la structure des complexes AP, exemple de AP1 ... 121
Figure 62 : Caractérisation de l’interaction entre les β-arrestines et la sous-unité β3A du complexe AP3 ... 122
Figure 64 : Mobilisation du calcium intracellulaire dans les cellules HL-60 exprimant le
récepteur FPRL1 étiqueté TAP ... 128
Figure 65 : Images d’immunofluorescence relatives à l’internalisation du récepteur FPRL1-TAP dans les cellules HEK-293... 130
Figure 66 : Purification par affinité en tandem du récepteur FPRL1-TAP exprimé dans les cellules HEK-293 marquées métaboliquement au 32P ... 132
Figure 67 : Schématisation d’une variante de la méthode TAP : le "split-TAG" ... 134
TABLE DES TABLEAUX
Tableau 1 : Lignées stables disponibles ou établies durant la thèse... 45Tableau 2 : Liste des anticorps utilisés... 49
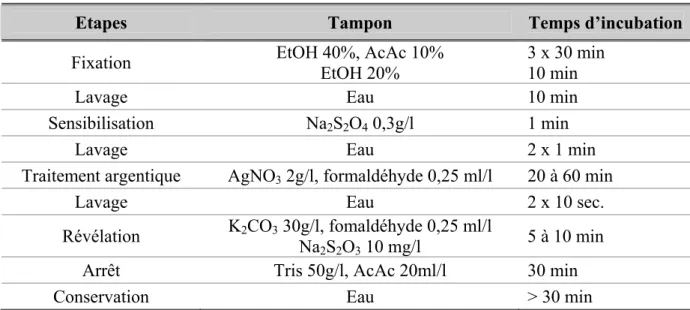
Tableau 3 : Etapes successives de la coloration à l’argent des protéines contenues dans un gel SDS-PAGE ... 51
Tableau 4 : Vecteurs des différents vecteurs utilisés ... 57
Tableau 5 : Liste des oligonucléotides utilisés... 58
Tableau 6 : Liste des peptides synthétiques testés ... 66
Tableau 7 : Activités biologiques des peptides (EC50 du test de mobilisation du calcium) dans les cellules exprimant les récepteurs de la famille FPR... 69
Tableau 8 : Séquences C-terminales des mutants de phosphorylation de FPRL1 ... 95
Tableau 9 : Compilation des résultats des analyses protéomiques menées sur les β-arrestines 1&2 étiquetées TAP ... 116
ABBREVIATIONS
17-AAG 17-(Allylamino)-17-demethoxyGeldanamycin ADN Acide DésoxyriboNucléique
ADP Adenosine DiPhosphate
AP Adaptor Protein (protéine adaptatrice) ARF6 ADP-Ribosylation Factor 6
ARN Acide RiboNucléique
ARNO ARF Nucleotide binding site Opener ASK1 Apoptosis Signaling Kinase 1
AT1AR Récepteur de l’Angiotensine II Type 1A ATCC American Type Culture Collection β2AR Récepteur β2-Adrénergique
Aβ42 fragment de 42 acides aminés de la protéine β-amyloïde A
BSA Bovine Serum Albumine
BOC ButOxyCarbonyl
Bt2-AMPc diButyryl-Adénosine MonoPhosphate cyclique C5aR Récepteur de l’anaphylatoxine C5a
CBP Calmodulin Binding Peptide (peptide de liaison à la calmoduline) CCV Clathrin Coated Vesicles (vésicules mantelées de clathrine)
CGD Chronic Granulomatous Desease (Granulomatose septique chronique)
COX Cytochrome c OXydase
cpm coups par minute
CsH Cyclosporine H
DABCO 1,4-DiAzaBiCyclo[2.2.2]Octane
DAG diacylglycérol
DiI 1,1’-Dioctadecyl-3,3,3’,3’-tetramethylIndolcarbocyanine perchlorate
DMSO DiMethylSulfOxide
dNTP désoxy-Nucleotide TriPhosphate
DUOX OXydase DUale
DTT DiThioThreitol
ECL Enhanced ChemiLuminescence
EDTA Ethylene Diamine Tetraacetic Acid EEA1 Early Endosome Antigen 1
EGFP Enhanced Green Fluorescent Protein EGTA Ethylene Glycol Tetraacetic Acid
Elk-1 Ets domain-containing Transcription Factor 1 ERK Extracellular signal-Regulated Kinase
FITC Fluorescein IsoThioCyanate FPR N-Formyl Peptide Receptor FPRL1/2 Formyl Peptide Receptor-Like ½ FSH Follicle-Stimulating Hormone Fura2-AM ester AcetoMethoxyl de Fura2
GEF Guanine-nucleotide Exchange Factor G-CSF Granulocyte Colony Stimulating Factor
GM-CSF Granulocyte-Monocyte Colony Stimulating Factor
GRK GPCR Kinase
GST Glutathion S-Transférase
GTP, GDP ou GMP Guanosine TriPhosphate, DiPhosphate ou MonoPhosphate HEK Human Embryonic Kidney fibroblast-like
HIV Human Immunodeficiency Virus
HRP HorseRadish Peroxidase
Hsp Heat shock protein
IB, IF, IP ImmunoBlot, ImmunoFluorescence, ImmunoPrécipitation
Ig Immunoglobuline
IGF-1 Insulin-like Growth Factor 1 IP3 Inositol triPhosphate
JNK c-Jun N-terminal Kinase
kDa kiloDalton
KRAB KRueppel-Associated Box
LAD Leucocyte Adhesion Deficiency (déficit d’adhésion des leucocytes)
LB Luria-Broth
LPS LipoPolySaccharide
LXA4 LipoXine A4
MAPK Mitogen-Activated Protein Kinase MEF Mouse Embryonic Fibroblast NFκB Necrosis Factor κB
MKK MAP Kinase Kinase
MKP7 MAP Kinase Phosphatase 7
NDx sous-unité x de la NADH déshydrogénase
NADPH forme réduite du Nicotinamide Adénine Dinucléotide Phosphate NES Nuclear Export Signal (signal d’export nucléaire)
NOX NADPH OXydase
PAF Platelet Activating Factor PAR Protease-Activated Receptor
PBS Phosphate Buffered Saline (tampon phosphate salin)
pEF promoteur du Facteur d’Elongation humain
PFA ParaFormAldéhyde
PI3K PhosphoInositide 3 Kinase
PIP2 PhosphoInositol-4,5-bisPhosphate PIP3 PhosphoInositol-3,4,5-triPhosphate PKA, PKC Protéine Kinase A ou C
PLA, C ou D PhosphoLipase A, C ou D PP1 PyrazolPyrimidine 1 PP2A Protein Phosphatase 2A
PTX ToXine Pertussique
ProtA répétition de deux domaines de la Protéine A liant les IgG RCPG Récepteurs Couplés aux Protéines G
RGS Regulator of G protein Signaling (régulateur de la signalisation par les protéines G)
Rho-GDI Rho-GDP Dissociation Inhibitor
ROS Reactiv Oxygen Species (espèces réactives de l’oxygène) SDS DodécylSulfate de Sodium
SDS-PAGE Electrophorèse sur Gel de PolyAcrylamide en présence de SDS
SH3 Src Homology domain 3
siRNA small interfering RNA (petits ARN interférents) snRNP small nuclear RiboNucleoProtein
SRAS Syndrome Respiratoire Aigü Sévère
TAP Tandem Affinity Purification (purification par affinité en tandem) TCA Acide TriChloroacetique
TetO Opérateur Tetracycline bactérien TetR Répresseur Tetracycline bactérien
TEV Tobacco Etch Virus
TGF-β Transforming Growth Factor TNF-α Tumor Necrosis Factor α
Tris Tris[hydroxymethyl]aminomethane WASP Wiskott-Aldrich Syndrome Protein
A/ Les phagocytes, première ligne de défense de l’organisme
contre les pathogènes
I- Les phagocytes
Les phagocytes (neutrophiles, monocytes et macrophages) constituent la première ligne de défense de l’organisme contre les micro-organismes pathogènes. Ces cellules appartiennent à la famille des leucocytes (ou globules blancs) qui comprend les granulocytes (polynucléaires neutrophiles, éosinophiles et basophiles), les lymphocytes et les monocytes.
Les polynucléaires neutrophiles représentent plus de la moitié des leucocytes (~65%), les lymphocytes comptant pour le quart, alors que les autres cellules sont très minoritaires. Comme les autres granulocytes, ces cellules présentent un noyau multilobé (et non pas plusieurs noyaux comme le laisserait supposer leur nom) et de nombreux granules dans leur cytoplasme (Figure 1). Ce sont les cellules leucocytaires possédant la plus grande capacité de défense contre les microorganismes pathogènes. Les fonctions cellulaires du neutrophile seront détaillées dans le chapitre II.
Figure 1 : Polynucléaire neutrophile entouré de globules rouges après coloration au May-Grünwald-Giesma
Image du Laboratoire d’Hématologie du C.H.U. d’Angers
Le monocyte est la plus grande cellule du sang. Il se caractérise par son noyau encoché et par son cytoplasme étendu parsemé de fines granulations. Il séjourne quelques jours dans le sang avant de gagner les tissus, dans lesquels il se transforme en macrophage (cellule ayant la capacité d’ingérer des particules solides). Sa fonction est proche de celle des polynucléaires neutrophiles. Le monocyte intervient dans l’élimination des bactéries, des particules étrangères ou des globules rouges âgés grâce à une activité de phagocytose moins spécifiquement antibactérienne que celle des neutrophiles. Il intervient également dans certaines étapes des réactions immunologiques, en particulier en assurant la transformation des protéines étrangères en peptides antigéniques.
Les macrophages, comme les polynucléaires neutrophiles, participent à la réaction inflammatoire, au nettoyage et à la remise en état des tissus altérés par l’agression de germes
étrangers. La capture des germes est facilitée par des molécules appelées opsonines (anticorps et complément). En exposant à leur surface, après les avoir digérées, les protéines étrangères, les macrophages jouent un rôle essentiel dans le développement de la réponse immunitaire en présentant les antigènes aux lymphocytes T.
Tous ces types leucocytaires, ainsi que les érythrocytes (globules rouges) et les plaquettes, dérivent de cellules souches hématopoïétiques pluripotentes dont la différenciation et la maturation ont lieu principalement dans la moëlle osseuse et sont contrôlées par des facteurs de croissance spécifiques. L’ensemble des processus conduisant aux cellules sanguines matures constitue l’hématopoïèse (Figure 2).
Figure 2 : Schématisation de l’hématopoïèse
Les neutrophiles, monocytes et macrophages dérivent de cellules progénitrices myéloïdes. Les cellules de la lignée HL-60 sont à un stade de différenciation situé en amont des progéniteurs de type granulocyte-monocyte. Les cellules HL-60 peuvent être différenciées vers un phénotype neutrophile ou monocyte-macrophage (d’après Goldsby et al., 2000).
La différenciation des cellules souches hématopoïétiques vers les cellules progénitrices myéloïdes et finalement vers le neutrophile mature est sous le contrôle successif de facteurs protéiques, notamment l’interleukine 3, et les facteurs de croissance GM-CSF (Granulocyte-Monocyte Colony Stimulating Factor) et G-CSF (Granulocyte Colony Stimulating Factor).
II- Les fonctions cellulaires du neutrophile
Les principales fonctions des polynucléaires neutrophiles se déclinent ainsi : le chimiotactisme permettant à la cellule de se rendre rapidement sur le site infectieux, la phagocytose visant à l’internalisation des bactéries, la dégranulation d’enzymes lytiques et l’explosion respiratoire génératrice de dérivés microbicides de l’oxygène.
II-1 Le chimiotactisme ou migration cellulaire dirigée
Lors d’un processus infectieux, ou après un dommage tissulaire ou une inflammation, des cytokines sont produites et vont modifier localement les propriétés adhésives et la perméabilité de l’endothélium. Les leucocytes qui circulent dans le sang adhèrent à l’endothélium par des mécanismes de margination, c'est-à-dire qu’ils s’accolent et roulent le long de la paroi du vaisseau sanguin. Sous l’effet des facteurs chimiotactiques établis en gradient de concentration décroissante à partir du foyer infectieux, les neutrophiles s’insinuent entre les cellules endothéliales et traversent la paroi vasculaire (diapédèse ou extravasation), puis migrent dans les tissus.
Figure 3 : Migration du neutrophile vers le site inflammatoire vaisseau
sanguin
chimioattractants
La contribution des protéines d’adhérence (sélectines, intégrines et membres de la superfamille des immunoglobulines), ainsi qu’un perpétuel remodelage du cytosquelette d’actine avec la participation de la myosine et des protéines d’ancrage de l’actine à la membrane plasmique (ezrine, radixine, moésine), sont les éléments majeurs générant les forces motrices de la chemotaxie. Ainsi, l’avant de la cellule émet de fins lamellipodes alors qu’un uropode contractile se forme à l’arrière de la cellule (Figure 4). Ces modifications ont lieu sous le contrôle de kinases, notamment la PI3 kinase (PhosphoInositide 3 kinase), et de petites GTPases monomériques telles que Rho, Rac et Cdc42 (Ridley et al., 2003). Il est remarquable de noter que, bien que la variation de concentration dans le gradient entre l’avant et l’arrière de la cellule soit infime, le neutrophile est tout de même capable de percevoir cette variation grâce à une compartimentation des signaux générés finement régulée dans le temps et dans l’espace.
Figure 4 : Polarisation d’un neutrophile
En présence de chimioattractant, le neutrophile change de morphologie et passe d’une cellule assez ronde et lisse à une cellule allongée et plissée et présentant des pseudopodes pointant vers les concentrations les plus fortes de chimioattractants, la cellule est alors dite "polarisée" (d’après
Weiner et al., 1999).
La migration dirigée des neutrophiles vers le site infectieux est donc un processus intégré de plusieurs étapes hautement complexes. Elle est initiée par des molécules appelées chimioattractants. Ces molécules sont libérées par les microorganismes invasifs ou générées de manière endogène par l’hôte. Elles diffusent en formant un gradient continu de concentration décroissante à partir du site inflammatoire. Le mouvement dirigé des neutrophiles vers le site infectieux est initié par la fixation de ces molécules chimiotactiques sur des récepteurs membranaires spécifiques, appelés récepteurs de chimioattractants. L’activation de ces récepteurs déclenche des cascades de signalisation intracellulaires dont les étapes seront plus détaillées dans un autre chapitre de cette thèse (cf. § B/II-4).
II-2 La phagocytose (pour revue : Verhoef et R. Visser, 1993)
La plupart des microorganismes ne peuvent être reconnus et phagocytés par les polynucléaires neutrophiles qu’après avoir été opsonisés, c'est-à-dire recouverts de facteurs du complément et d’anticorps pouvant être reconnus par les récepteurs des neutrophiles. La paroi bactérienne peut être chargée en facteur activé C3b du complément qui est reconnu par les récepteurs CR1 et CR3 du neutrophile. Ce type d’opsonisation suffit à l’attachement mais pas à l’ingestion des microorganismes qui requiert des interactions plus fortes de type antigène-anticorps. Ainsi, des immunoglobulines (Ig) G1 et G3 se fixent à des antigènes spécifiques à la surface des bactéries et servent de ligands aux neutrophiles qui présentent des récepteurs aux fragments Fc de ces IgGs. D’autres opsonines plus spécifiques du type de microorganismes sont capables d’initier l’interaction hôte-pathogène, c’est le cas de la "lipopolysaccharide (LPS) binding protein" qui se lie au LPS des bactéries Gram-négatif et au récepteur CD14 des cellules phagocytaires.
Figure 5 : Phagocytose de candida albicans par un polynucléaire neutrophile
Comme l’illustre la Figure 5, après opsonisation, les pathogènes sont littéralement ingérés par les neutrophiles grâce à l’émission de pseudopodes qui entourent les microorganismes opsonisés et vont permettre leur engloutissement par fusion membranaire dans une vacuole appelée le phagosome. C’est dans ce compartiment isolé du reste de la cellule que le pathogène va être tué et digéré.
II-3 La mobilisation des granules
Les neutrophiles matures ne synthétisent que peu de protéines de novo. Ils possèdent au contraire des stocks de protéines qu’ils peuvent mobiliser très rapidement suivant les besoins. Ces protéines sont compartimentées dans des organelles de stockage intracellulaires appelées
granules. Par ailleurs, les granules peuvent fusionner avec le phagosome contenant un pathogène. Ce processus, appelé dégranulation, permet le déversement dans les vacuoles de phagocytose de nombreuses enzymes lysosomales, donnant ainsi naissance au phago-lysosome. Les enzymes lysosomales interviennent dans la digestion des pathogènes phagocytés. D’autre part, la fusion des granules peut avoir lieu avec la membrane plasmique, ce qui permet de compenser la surface membranaire perdue lors de la phagocytose mais aussi d’augmenter rapidement la quantité de protéines membranaires, notamment les récepteurs de surface ("up-regulation"). Le contenu des granules peut aussi être déversé dans le milieu extérieur.
Figure 6 : Schématisation de la dégranulation du neutrophile
Les polynucléaires contiennent plusieurs types de granules (pour revue : Segal, 2005). Dans les granules primaires (ou granules denses azurophiles) sont contenus des peptides et des protéines visant principalement à la destruction des bactéries, comme la myéloperoxydase, le lysozyme (hydrolyse des parois bactériennes), la cathepsine G et l’élastase. Dans les granules secondaires (ou granules spécifiques), on note, entre autres, la présence de lysozyme, lactoferrine, gélatinase et de protéines membranaires comme le flavocytochrome b558 de la NADPH oxydase. Les granules tertiaires (ou granules gélatinase) contiennent de la gélatinase mais pas de lactoferrine. Les vésicules sécrétrices sont les vésicules les plus rapidement mobilisables et contiennent, entre autres, la phosphatase alcaline et le flavocytochrome b558 de la NADPH oxydase. Elle présentent également dans leur membrane une réserve de récepteurs, notamment de récepteurs de chimioattractants, qui peut être mobilisée rapidement (Sengelov et al., 1994). Ces granules fusionnent avec la membrane plasmique des neutrophiles activés pour augmenter le nombre de récepteurs en surface et amplifier la réponse aux chimioattractants.
II-4 L’explosion respiratoire, la NADPH oxydase (pour revues : Babior, 2004; Vignais, 2002 )
La principale fonction microbicide des neutrophiles consiste en la libération d’espèces réactives de l’oxygène (ROS) dans l’environnement du site infectieux et dans les vésicules phago-lysosomales. Leur production est assurée par la NADPH oxydase phagocytaire qui est une enzyme présente dans les cellules phagocytaires et également dans d’autres types cellulaires, comme les lymphocytes B (Morel et al., 1993), les cellules endothéliales (Gorlach et al., 2000) et les ostéoclastes (Yang et al., 2001). Elle catalyse la production d’ions superoxyde (O2-) à partir de l’oxygène et du NADPH (forme réduite du Nicotinamide Adénine Dinucléotide Phosphate) selon la réaction :
NADPH + 2 O2→ NADP+ + H+ + 2 O2-.
La NADPH oxydase est un complexe protéique composé de protéines membranaires et cytosoliques qui s’assemblent lors de l’activation (Figure 7). Il comprend :
- deux sous-unités membranaires, formant le cytochrome b558 : gp91phox et p22phox, - trois facteurs cytosoliques : p67phox, p47phox et p40phox,
- et une petite protéine G monomérique (Rac).
Figure 7 : Schématisation de l’assemblage du complexe NADPH oxydase phagocytaire L’activité oxydase ne s’exprime que si tous les éléments sont assemblés de manière coordonnée au niveau de la membrane. L’assemblage de la forme active de la NADPH oxydase implique des processus de phosphorylation qui sont essentiels à l’activation et de multiples interactions entre les composants de la NADPH oxydase. Les interactions moléculaires entre ces composants relèvent principalement d’interactions de type SH3 (Src Homology domain 3) et des domaines polyproline.
Les sous-unités gp91phox et p22phox forment le flavocytochrome b558. La sous-unité gp91phox comporte tous les groupes prosthétiques nécessaires au transfert d’électrons du NADPH vers l’oxygène (le FAD, Flavine Adénine Dinucléotide, deux hèmes, et un site de liaison pour le NADPH).
Dans les neutrophiles au repos, les protéines p67phox, p47phox et p40phox sont présentes sous forme d’un complexe cytosolique de 250 kDa. Lors de l’activation de la NAPDH oxydase, ces protéines, qualifiées de sous-unités régulatrices, migrent à la membrane plasmique pour se fixer au flavocytochrome b558 et l’activer.
L’activation met également en jeu la protéine Rac. Au repos, Rac liée au GDP (Guanosine DiPhosphate) est majoritairement présente dans le cytosol, sous forme complexée à la protéine inhibitrice Rho-GDI (Rho-GDP Dissociation Inhibitor). Dans le complexe oxydase actif, Rac, dissociée de Rho-GDI, est liée au GTP (Guanosine TriPhosphate). La migration de Rac à la membrane s’effectue de manière indépendante par rapport aux autres facteurs cytosoliques cités précédemment.
Le rôle physiologique de la NADPH oxydase phagocytaire est primordial dans la réponse de l’hôte aux infections. Un dysfonctionnement de cette enzyme, causé par des mutations affectant les composants de ce complexe protéique, est à l’origine de la granulomatose septique chronique (CGD), qui est une maladie héréditaire caractérisée par des infections graves et récidivantes.
Il est à noter que d’autres oxydases ont récemment été mises en évidence dans d’autres types cellulaires que les phagocytes, il s’agit d’homologues de gp91phox appelés NOXs pour NADPH oxydases ou DUOXs pour oxydases duales. Dans cette nouvelle nomenclature, gp91phox a été dénommée NOX2. Les NOXs autres que gp91phox sont exprimées principalement dans l’épithélium, les cellules musculaires lisses et l’endothélium, elles génèrent de petites quantités de ROS, apparemment à des fins de transduction du signal dans les mécanismes de croissance, d’angiogenèse et d’apoptose (Lambeth, 2004). Il a été montré plus récemment que les NOXs peuvent être ciblées dans des compartiments cellulaires discrets pour la production localisée de ROS ; ce serait le cas, par exemple, au sein des complexes focaux des lamellipodes, où les ROS seraient requis pour la migration cellulaire dirigée (Ushio-Fukai, 2006).
B/ Les récepteurs de chimioattractants
Tout système biologique se doit de pouvoir "ressentir" son environnement et de réagir en conséquence.
La notion de "récepteur membranaire spécifique" a été introduite par les travaux de l’immunologiste Paul Ehrlich dans le cadre de l’étude des toxines, travaux pour lesquels il a reçu le prix Nobel de physiologie ou médecine en 1908.
Figure 8 : Schématisation de la notion de récepteur membranaire spécifique dessin suggestif, réalisé par Paul Ehrlich lui-même (Ehrlich, 1900).
Le récepteur est enchâssé dans la membrane plasmique et sa spécificité est apportée par sa structure qui présente une poche de liaison parfaitement adaptée à la reconnaissance de la molécule signal, cette molécule est appelée agoniste. La fixation de l’agoniste sur son récepteur membranaire à l’extérieur de la cellule entraîne un changement conformationnel du récepteur qui conduit à la mise en place d’interactions protéiques à l’intérieur de la cellule qui aboutissent finalement à une cascade de signaux intracellulaires menant à terme à l’exécution de fonctions cellulaires adaptées.
Les récepteurs membranaires peuvent être classés selon deux types majeurs de transduction du signal : les récepteurs transduisant eux-mêmes le signal (récepteurs canaux, récepteurs enzymes) et les récepteurs couplés à une protéine effectrice intermédiaire, dont font partie les récepteurs de chimioattractants qui sont couplés à une GTPase hétérotrimérique.
I- Les récepteurs couplés aux protéines G hétérotrimériques
(pour revue : Bockaert et al., 2002)La superfamille des récepteurs couplés aux protéines G (RCPGs) est la famille de récepteurs la plus importante, elle compte chez les mammifères pour près de 1000 gènes. Hormis le fait qu’ils soient tous couplés à une protéine G hétérotrimérique, la caractéristique majeure de ces récepteurs est leur structure secondaire faite de 7 hélices transmembranaires, c’est pourquoi on les nomme également récepteurs à 7 hélices ou à 7 domaines transmembranaires ou encore de façon plus imagée récepteurs serpentins. La partie N-terminale extracellulaire du récepteur diverge énormément entre les membres de la famille des RCPGs, c’est elle qui assure la reconnaissance des ligands spécifiques. La rhodopsine est le seul RCPG dont la structure cristallographique ait été résolue (Okada et al., 2000). La partie C-terminale cytoplasmique de ce récepteur est composée d’une huitième hélice, plateforme participant à la reconnaissance de la protéine G. Les RCPGs sont retrouvés dans la plupart des types cellulaires, et sont impliqués dans un spectre extrêmement large de fonctions biologiques. De nombreuses pathologies, comme les rétinites pigmentaires ou l’hyperactivité thyroïdienne, sont liées à un mauvais fonctionnement ou à une mauvaise régulation des RCPGs (pour revue : Spiegel, 1996).
Figure 9 : Représentation schématique d’un RCPG dans la membrane plasmique Les zones de fixation du ligand sont figurées en vert, celles de fixation de la protéine G en bleu et les zones qui participent à la régulation du récepteur par phosphorylation en rouge.
Fixation
du ligand
Sites de
phosphorylation
Fixation de la
protéine G
RCPG
Une fois stimulés, les récepteurs à 7 hélices subissent un changement de conformation qui induit l’activation de la protéine G hétérotrimérique à laquelle ils sont couplés. Ces protéines G ont été découvertes dans les années 1970. Martin Robdell et Alfred Gilman ont tous deux reçu le prix Nobel de physiologie ou médecine en 1994 pour leurs travaux séparés sur ces protéines G et leur rôle dans la transmission des signaux dans les cellules. Les protéines G hétérotrimériques sont composées de trois sous-unités : α qui fixe les nucléotides de type guanine et porte l’activité GTPase, et β et γ qui restent associées sous forme d'un dimère. Leur cycle d’activation commence par la dissociation de la sous-unité α du dimère βγ (Figure 10). La sous-unité α est alors le siège de l’échange du GDP pour du GTP et c’est sous sa forme associée au GTP qu’elle est active et peut agir sur des effecteurs de la signalisation (cf. § B/II-4). Les fonctions du dimère βγ sont moins bien connues mais engagent également des effecteurs de la signalisation. La désactivation implique une hydrolyse du GTP en GDP, catalysée par des protéines RGS (Regulator of G protein Signaling). En fin de cycle, la sous-unité α se retrouve sous forme inactive liée au GDP et se réassocie avec le dimère βγ pour reformer la protéine G hétérotrimérique inactive. Il existe de nombreuses isoformes des sous-unités β, γ et surtout α des protéines G. Les différents types de sous-unités Gα sont classés par familles : αi, αs, αq et α12.
II- Les récepteurs chimiotactiques
Les molécules chimioattractantes ont été définies comme telles car elles induisent le chimiotactisme des phagocytes qui reconnaissent ces molécules grâce à des récepteurs de surface de type RCPG appelés récepteurs chimiotactiques (pour revue : Ye et Boulay, 1997). Parmi ces récepteurs sont retrouvés :
- les récepteurs de chimioattractants dits classiques, comme le récepteur de l’anaphylatoxine C5a (C5aR), les récepteurs de la famille FPR (N-Formyl Peptide Receptor), les récepteurs du leukotriène B4 et du PAF (Platelet Activating Factor)
- et les récepteurs de chimiokines.
Ils présentent des caractéristiques communes les différenciant des autres sous-groupes de RCPGs (pour revue : Murphy, 1994) :
- leurs séquences sont similaires en taille avec environ 350 acides aminés. Ce sont les plus petits membres de la famille des RCPGs. Cette petite taille est due à une séquence N-terminale de longueur moyenne, à une 3ème boucle intracellulaire i3 très courte et à un domaine C-terminal relativement court.
- leurs différents membres présentent plus de 20 % d’identité entre eux.
- la très courte boucle i3 (de 16 à 22 acides aminés) est enrichie en acides aminés basiques. Dans la plupart des cas, cette boucle i3 peut être modélisée sous la forme d’une hélice α qui présente un côté hydrophobe et l’autre chargé positivement.
- le segment N-terminal extracellulaire est le plus souvent acide. - les gènes codant ces récepteurs sont en général sans intron.
II-1 Description générale des récepteurs de la famille FPR
Le récepteur humain FPR (N-Formyl Peptide Receptor) est le premier récepteur de chimioattractants présent à la surface des neutrophiles à avoir été caractérisé biochimiquement (Showell et al., 1976). Il doit son nom au fait qu’il se lie avec une haute affinité à des peptides N-formylés, et notamment le f-MLF (N-formyl-methionyl-leucyl-phénylalanine). Sa structure primaire a été élucidée au début des années 90 grâce au clonage de deux formes allèliques à partir d’une banque d’ADNc issue de cellules HL-60 différenciées (Boulay et al., 1990a; Boulay et al., 1990b). Depuis, la famille des récepteurs de chimioattractants des cellules
myéloïdes s’est agrandie (pour revue : Ye et Boulay, 1997). Deux récepteurs homologues à FPR, FPRL1 et 2 (FPR-Like 1 et 2), ont été clonés (Bao et al., 1992; Murphy et al., 1992). Les trois gènes correspondant à la famille des récepteurs FPR forment un cluster sur le chromosome 19. Malgré un fort degré d’identité avec FPR (69%), FPRL1 ne se lie au f-MLF qu’avec une faible affinité. FPRL1 est également appelé récepteur de la lipoxine A4 (LXA4), car ce médiateur lipidique a été le premier ligand endogène de haute affinité découvert pour FPRL1 (Fiore et al., 1994). Le troisième membre de la famille FPR, FPRL2, partage 56% d’identité avec FPR, mais il ne se lie pas, ni ne répond au peptide f-MLF. Les deux homologues de FPR montrent une forte identité avec celui-ci au niveau des parties cytoplasmiques, alors que les boucles extracellulaires sont plus divergentes.
Une étude de l’occurrence de polymorphismes pour les gènes codant FPR et FPRL1 a révélé qu’il n’existe pas de polymorphisme pour FPRL1, alors que plusieurs isoformes de FPR ont pu être mises en évidence (Sahagun-Ruiz et al., 2001). Certaines de ces isoformes sont associées à des phénotypes plus ou moins marqués. Les formes R190W et N192K sont par exemple associées à la periodontite aggressive (inflammation des gencives) (Zhang et al., 2003). Les isoformes FPR98 et FPR-G6 pourraient, quant à elles, résulter en un défaut de couplage du récepteur à la protéine G (Wenzel-Seifert et Seifert, 2003).
II-2 Ligands des récepteurs de la famille FPR (pour revues : Migeotte et al., 2006 et Rabiet et al., 2007, Biochimie, sous presse)
Les zones de fixation des ligands sur les récepteurs de chimioattractants impliquent le domaine N-terminal et les boucles extracellulaires e1 et e2, toutes deux reliées par un pont disulfure. De nombreux agonistes et antagonistes des récepteurs de la famille FPR sont désormais connus. FPR et FPRL1 sont qualifiés de récepteurs infidèles qui peuvent être activés par un spectre assez large d’agonistes peptidiques ne présentant pas de caractéristiques structurales communes. Ces agonistes peuvent être synthétiques, dérivés de l’hôte ou des pathogènes. FPRL2 est activé avec une faible capacité par quelques peptides chimiotactiques non-formylés qui sont également des agonistes de FPRL1 (Betten et al., 2001; Christophe et al., 2001).
II-2-1 Agonistes
Des criblages aléatoires de banques de peptides ont permis de découvrir des agonistes des récepteurs de la famille FPR. Le peptide WKYMVM et le peptide WKYMVm, où la méthionine finale présente une conformation dextrogyre, diffèrent dans leur capacité à activer les récepteurs de la famille FPR (Baek et al., 1996; Seo et al., 1997). Le peptide WKYMVm active les trois récepteurs FPR, FPRL1 et FPRL2, et présente une meilleure affinité pour FPRL1. Par contre, l’isomère WKYMVM n’a quasiment aucune capacité à activer FPR (Christophe et al., 2001; Dahlgren et al., 2000).
Les principaux agonistes naturels des récepteurs FPR dérivent des virus et bactéries. Des peptides dérivés de protéines de l’enveloppe du virus HIV1 ont été décrits comme agonistes des récepteurs FPR et FPRL1 (Su et al., 1999b). Le peptide Hp(2-20) de la bactérie Helicobacter pylori est capable d’activer la mobilisation du calcium et la NADPH oxydase chez le neutrophile par l’intermédiaire du récepteur FPRL1 (Bylund et al., 2001).
Il existe également des agonistes des récepteurs de la famille FPR qui proviennent de l’hôte même. Le premier à avoir été découvert a été la lipoxine A4 qui est un agoniste de haute affinité pour FPRL1 (Fiore et al., 1994), mais cet agoniste est un mauvais activateur des fonctions du neutrophile. Des protéines de type amyloïde, ou des fragments dérivés de telles protéines, peuvent activer les cellules myéloïdes via FPRL1 (Le et al., 2001; Su et al., 1999a). L’humanine, un peptide neuroprotecteur de 24 acides aminés retrouvé dans la région occipitale du cerveau, utilise également le récepteur FPRL1 de manière fonctionnelle. De manière intéressante, la cathepsine G, qui est libérée lors de la dégranulation du neutrophile, présente des propriétés chimiotactiques par l’intermédiaire du FPR (Sun et al., 2004).
D’autres agonistes N-formylés des récepteurs de la famille FPR ont pu être identifiés durant ce travail de thèse, certains provenant de Listeria monocytogenes et d’autres, de façon plus originale, de protéines mitochondriales humaines (cf. section A des Résultats et Discussions et Rabiet et al., 2005).
II-2-2 Antagonistes
Ces antagonistes sont des ligands des récepteurs, au même titre que les agonistes, à ceci près qu’ils n’entraînent pas d’activation du récepteur, c'est-à-dire ni changement de conformation, ni signalisation intracellulaire, ils se comportent donc comme des inhibiteurs.
Des composés antagonistes ont été identifiés pour FPR et FPRL1. Pendant de nombreuses années, le peptide t-BOC, forme N-ButOxyCarbonylée du peptide FLFLF, a été considéré comme l’antagoniste le plus puissant de l’action du f-MLF (Freer et al., 1980). En 1993, le décapeptide cyclique cyclosporine H (CsH), dérivé du champignon Tolypocladium inflatum, a été décrit comme un inhibiteur fort de FPR, environ dix fois plus puissant et sélectif que les antagonistes connus jusqu’alors (Wenzel-Seifert et Seifert, 1993). La CsH agit comme un agoniste inverse, faisant passer le récepteur FPR d’un état actif (R*) à un état inactif (R) (Wenzel-Seifert et al., 1998).
Il a été montré que certains pathogènes étaient capables de sécréter des protéines ou peptides avec des propriétés antagonistes. Ainsi les virus HIV 1 et 2, Ebola, le coronavirus du SRAS (Syndrome Respiratoire Aigü Sévère) possèdent des protéines de surface dont dérivent des peptides antagonistes pour FPR (Mills, 2006). La bactérie Staphylococcus aureus est capable de sécréter des petites protéines à activité antagoniste pour FPR (de Haas et al., 2004; Prat et al., 2006). Ces microorganismes semblent en cela avoir développé des stratagèmes pour échapper à la première ligne de défense de l’hôte constitué par les phagocytes.
Récemment, deux antagonistes du récepteur FPRL1 ont été identifiés, il s’agit du peptide synthétique WRWWWW (Bae et al., 2004) et de PBP10, un peptide dérivé de la gelsoline (Cunningham et al., 2001). Ce sont les rares composés à inhiber spécifiquement l’activité de FPRL1.
II-3 Localisation cellulaire et tissulaire des récepteurs de la famille FPR
Parallèlement au nombre grandissant de ligands de la famille FPR, la distribution cellulaire et tissulaire de ces récepteurs s’est élargie. FPR et FPRL1 sont exprimés dans macrophages et les neutrophiles alors que FPRL2 est présent dans les monocytes-macrophages mais pas dans les neutrophiles. L’expression des récepteurs des peptides N-formylés dans une variété de cellules autres que les cellules phagocytaires (Migeotte et al., 2006 et Rabiet et al., 2007, Biochimie, sous presse) suggère que ces récepteurs puissent avoir des rôles fonctionnels autres que la défense de l’hôte dans la réponse immunitaire innée. Un rôle précis du récepteur FPRL1, exprimé dans les cellules microgliales, a d’ailleurs été évoqué récemment dans les maladies neurodégénératives (pour revue : Iribarren et al., 2005). FPRL1 peut en effet être activé par un fragment de 42 acides aminés de la protéine β-amyloïde A (Aβ42), dont la surproduction et la précipitation sous forme de plaques ou de fibrilles sont
impliquées dans la maladie d’Alzheimer. Le rôle du complexe FPRL1/Aβ42 serait double puisqu’il participerait à la persistance d’une activité proinflammatoire par activation et recrutement des phagocytes, et que son internalisation entraînerait la formation de fibrilles d’Aβ42 dans ces phagocytes (Cui et al., 2002; Le et al., 2001).
II-4 Signalisation intracellulaire en aval des récepteurs de chimioattractants
La première étape des voies de signalisation enclenchées par la stimulation des RCPGs est l’activation de la protéine G qui leur est associée. L’interaction entre un grand nombre de RCPGs et leur protéine G est dépendante du motif consensus E/DRY situé sur la face intracellulaire du troisième pilier transmembranaire du récepteur (Oliveira et al., 1994). Plusieurs études ont montré que le couplage des récepteurs de chimioattractants avec la protéine Gi2 implique de multiples sites d’interaction, à la fois dans la boucle intracellulaire i1, sur la liaison entre les boucles intracellulaires i2 et i3 et au niveau de régions discrètes de la queue carboxyl-terminale (Bommakanti et al., 1995).
Les récepteurs de chimioattractants sont principalement couplés à une protéine G de type Gi2 (Gierschik et al., 1989) et les fonctions du polynucléaire neutrophile induites par les chimioattractants sont inhibées par un traitement avec la toxine pertussique (PTX) de Bordetella pertussis, qui provoque une ADP-ribosylation spécifique des sous-unités αi des protéines G hétérotrimériques de type Gi (Bokoch, 1995). Il a été montré que les récepteurs FPR et C5aR sont également capables de coupler avec les protéines Gi1, G0, et les protéines G insensibles à la PTX de type Gz et G16 (Tsu et al., 1995).
Les voies de signalisation stimulées en aval des récepteurs de chimioattractants sont hautement complexes, une vue schématique et non exhaustive de ces voies est présentée en Figure 11. Après la dissociation de la sous-unité α, le dimère βγ active la phospholipase Cβ2 (PLCβ2) (Camps et al., 1992) et la phosphoinositide 3 kinase γ (PI3Kγ) (Stoyanov et al., 1995). La PI3Kγ convertit le phosphoinositol-4,5-bisphosphate (PIP2) membranaire en phosphoinositol-3,4,5-triphosphate (PIP3). La délétion de la PI3Kγ chez la souris a révélé que cette enzyme est requise pour la migration dirigée des neutrophiles dans un gradient de peptide f-MLF (Hannigan et al., 2002) et pour la génération d’ions superoxyde initiée par la stimulation des récepteurs de chimioattractants (Hirsch et al., 2000). La PLCβ2 hydrolyse le
PIP2 membranaire en diacylglycérol (DAG) et inositol triphosphate (IP3). Ce dernier cause la libération du calcium des vésicules de stockage intracellulaires.
Le calcium et le DAG entraînent l’activation des isoformes de la protéine kinase C (PKC). Les neutrophiles et les cellules HL-60 différenciées vers un phénotype "neutrophile-like" expriment les isoformes classiques de la PKC, α, βI et βII, la nouvelle isoforme de PKC δ et l’isoforme atypique ζ (Tardif et al., 1998; Tsao et Wang, 1997). Les études pour déterminer quelles isoformes de PKC ont un rôle dans les fonctions du neutrophile sont assez contradictoires. Des études in vitro ou dans des systèmes reconstitués de génération d’ions superoxyde indiquent que certaines isoformes de PKC sont impliquées dans l’activation et la régulation de la NADPH oxydase.
Figure 11 : Représentation schématique des voies de signalisation enclenchées par les récepteurs de chimioattractants
L’activation d’autres kinases est également intimement liée à la signalisation relative aux chimioattractants. Dans le neutrophile, les Mitogen-Activated Protein (MAP) kinases ERK1/2 (Extracellular signal-Regulated Kinases 1 et 2) et p38 sont activées par les chimioattractants. Ces deux voies de signalisation participent aux phénomènes d’adhérence, de chimiotactisme et de production d’ions superoxyde. La voie menant à l’activation de p38 n’est pas encore complètement élucidée alors que celle vers l’activation de ERK1/2 a été reconstituée (Lopez-Ilasaca et al., 1997). Le dimère βγ de la protéine G recrute la PI3Kγ à la membrane plasmique, augmentant ainsi l’activité des tyrosine kinases de la famille Src, qui
phosphorylent des protéines d’échafaudage comme la protéine adaptatrice Shc. L’association fonctionnelle entre Shc, Grb2 et la protéine SOS intervient alors et mène à l’activation de la voie Ras-Raf-MEK-ERK.
Les effecteurs intracellulaires couplés aux cascades de signalisation en aval des récepteurs de chimioattractants incluent entre autres, la phospholipase A2-α, la phospholipase D, les MAP kinases et les kinases de la famille Src (Ma et al., 2000; Rane et al., 1997; Torres et al., 1993). La PLA2-α est phosphorylée par les MAP kinases et elle est transférée à la membrane par un mécanisme dépendant du calcium (Lin et al., 1993). La PLA2-α pourrait être requise pour l’activation de la NADPH oxydase car la production d’ions superoxyde est inhibée par traitement des cellules différenciées HL-60 avec des ARN antisens dirigés contre cette phospholipase. Le défaut résultant peut être compensé par l’addition d’acide arachidonique (Dana et al., 1998). Toutefois, ces résultats sont en contradiction avec une étude récente qui montre que la délétion du gène de la PLA2-α chez la souris supprime totalement la libération d’acide arachidonique mais n’affecte pas la production d’ions superoxyde (Rubin et al., 2005).
La stimulation des récepteurs de chimioattractants mène à l’activation de GTPases monomériques, appelées protéines G de petite masse moléculaire, de la famille Rho (Rho, Rac et Cdc42), par l’intermédiaire de l’activation de protéines GEFs (Guanine-nucleotide Exchange Factors) comme Vav1 et pRex1 (Kim et al., 2003; Welch et al., 2002). Les GTPases de type Rho sont des régulateurs primordiaux des fonctions des leucocytes (pour revue : Sanchez-Madrid et del Pozo, 1999). RhoA serait impliqué dans l’adhésion dépendante des intégrines induite par les chimioattractants. La Rho-GTPase Rac2, dont l’expression est restreinte aux cellules hématopoïétiques, joue un rôle central dans la formation du complexe actif de la NADPH oxydase (pour revue : Dinauer, 2003). Les Rho-GTPases Rac1, Rac2 et Cdc42 sont impliquées dans le remodelage du cytosquelette d’actine. L’activation de Cdc42 serait responsable de la relaxation de la conformation auto-inhibitrice de la protéine WASP (Wiskott-Aldrich Syndrome Protein), une protéine à domaines multiples qui est un activateur du complexe de nucléation Arp2/3 (Machesky et Gould, 1999; Symons et al., 1996). Il a été montré que la queue cytoplasmique C-terminale du récepteur C5aR interagit avec WASP lorsque la conformation auto-inhibitrice est relâchée (Tardif et al., 2003). Cette interaction pourrait être un moyen pour ce récepteur de contrôler spatialement les sites de polymérisation de l’actine pendant la migration dirigée des leucocytes le long d’un gradient de C5a.
II-5 Régulation de l’activité des récepteurs de chimioattractants
La restriction et la localisation de l’activité de signalisation sont d’un grand intérêt pour la régulation des fonctions leucocytaires. Une modulation rapide de la signalisation au niveau du récepteur est en effet essentielle à la cellule pour percevoir les modifications infimes de concentration de chimioattractants lors de la migration dirigée le long du gradient chimiotactique. De même, l’atténuation de réponses leucocytaires comme la dégranulation ou la production d’ions superoxyde est d’une importance majeure dans le contrôle de l’inflammation et pour empêcher des dommages tissulaires trop importants.
II-5-1 Modulation de l’activité des récepteurs
De nombreuses études biochimiques suggèrent que les RCPGs peuvent former des structures dimériques ou multimériques (pour revues : Bai, 2004; Park et al., 2004). L’oligomérisation joue un rôle important dans le trafic des récepteurs et la régulation des voies de signalisation. De plus, l’hétéro-oligomérisation entre des récepteurs différents peut engendrer des nouvelles spécificités vis-à-vis des agonistes et contribuer à la modulation des fonctions des récepteurs. L’oligomérisation du récepteur C5aR est constitutive et semblerait jouer un rôle dans la biosynthèse et le transport du récepteur (Floyd et al., 2003). Le récepteur FPR a, quant à lui, été décrit comme ne formant pas de dimères (Gripentrog et al., 2003). L’hétéro-oligomérisation est également un moyen utilisé par la cellule pour moduler l’activité de certains récepteurs suite à la stimulation d’autres récepteurs, ce processus est apparenté à la désensibilisation hétérologue. Le récepteur de chimiokine CCR5 est par exemple phosphorylé et internalisé suite à la stimulation du récepteur C5aR, avec lequel il forme des hétéro-dimères (Huttenrauch et al., 2005).
II-5-2 L’inactivation des récepteurs
La terminaison de la réponse aux chimioattractants rend les cellules réfractaires à une seconde stimulation du même récepteur. Plusieurs mécanismes mutuellement non exclusifs peuvent participer à cette perte d’activité des récepteurs : la phosphorylation des récepteurs, leur interaction avec des molécules accessoires comme les β-arrestines et finalement leur internalisation.
Figure 12 : Représentation schématique du trafic régulé des récepteurs de chimioattractants
Suite à leur phosphorylation, les récepteurs peuvent interagir avec les β-arrestines qui participent au processus d’endocytose par recrutement de la clathrine et de la protéine adaptatrice AP2. Après passage dans des vésicules d’endocytose, les récepteurs sont recyclés à la membrane plasmique dans un état fonctionnel.
II-5-2-1 Phosphorylation
La phosphorylation des récepteurs de chimioattractants est assurée majoritairement par des GRKs (GPCR Kinases) et par des protéines kinases de type PKC.
Le récepteur de l’anaphylatoxine C5a possède un site consensus de phosphorylation par la PKC (R-X-X-S-X-R-X) sur la troisième boucle intracellulaire. Sa phosphorylation est partiellement inhibée par des inhibiteurs de PKC (Tardif et al., 1993). Néanmoins, les sites majeurs de phosphorylation de ce récepteur sont restreints à sa région carboxyl-terminale (Giannini et al., 1995). La détermination de l’isoforme de GRK impliquée dans la phosphorylation du C5aR est encore controversée. Le C5aR est phosphorylé majoritairement sur des résidus sérines (Ser314, Ser317, Ser327, Ser332, Ser334 et Ser338) (Giannini et al., 1995). La phosphorylation de ce récepteur fait appel à un processus hiérarchique (Christophe et al., 2000; Naik et al., 1997). La phosphorylation de l’une des deux paires de sérines Ser332/Ser334 ou Ser334/Ser338 est un prérequis pour la phosphorylation des autres sérines et elle est suffisante pour permettre l’internalisation du récepteur par la formation d’un complexe stable avec les β-arrestines jusque dans des vésicules intracellulaires (Braun et al., 2003). Une forme
mutante du récepteur C5aR non phosphorylable n’est pas internalisée (Naik et al., 1997) et induit une signalisation ainsi qu’une production d’ions superoxyde prolongées (Christophe et al., 2000).
310 320 330 340
| * * | * | * * * | C5aR 296NPIIYVVAGQGFQGRLRKSLPSLLRNVLTE-ESVVRESKSFTRSTVDTMAQKTQAV350
310 320 330 340
| | **|** * * **|
FPR 297NPMLYVFMGQDFRERLIHALPASLERALTE--DSTQTSDTATNSTLPSAEVALQAK350
| | | | | |
FPRL1 298NPMLYVFVGQDFRERLIHSLPTSLERALSE--DSAPTNDTAANSASPPAETELQAM351
| || | | | | | | |
FPRL2 298NPILYVFMGRNFQERLIRSLPTSLERALTEVPDSAQTSNTDTTSASPPEETELQAM353 Figure 13 : Alignement des domaines C-terminaux de récepteurs de chimioattractants Les résidus sérines et thréonines sont indiqués en gras sur fond gris. Les astérisques rouges indiquent, pour C5aR et FPR, les résidus phosphorylés de manière agoniste-dépendante, parmi lesquels ceux qui sont primordiaux pour la phosphorylation des autres résidus sont surlignés en vert et jaune. Les deux sites putatifs de phosphorylation par les GRK sont soulignés en rouge et bleu, ils sont précédés d’un acide aminé de caractère acide surligné en noir.
La queue C-terminale du récepteur FPR contient 11 sérines et thréonines. Ces deux types de résidus sont phosphorylés (Prossnitz et al., 1995). L’utilisation de mutants présentant un défaut de phosphorylation a conclu que huit de ces résidus, localisés entre les sérine S328 et thréonine T339, sont critiques pour l’internalisation et la désensibilisation du récepteur FPR (Maestes et al., 1999). Ces résidus sont répartis en deux domaines, contenant 4 sérines ou thréonines et précédés d’un acide aminé à caractère acide. Ceci est une caractéristique des sites de phosphorylation de type GRK (Onorato et al., 1991). Des études par mutagenèse dirigée ont montré que les résidus glutamine 326, aspartate 327 et aspartate 333 sont critiques pour la phosphorylation de FPR (Prossnitz et al., 1995). La phosphorylation du récepteur FPR n’est pas dépendante des PKC, des PKA ou des tyrosine kinases. La queue C-terminale de FPR peut être phosphorylée par les kinases GRK2 et GRK3, mais pas par GRK5 ou 6 (Prossnitz et al., 1995). La phosphorylation de FPR est un processus hiérarchique dans lequel la phosphorylation de sérines et thréonines adjacentes (Ser328/Thr329 ou Thr331/Ser332) est requise pour la phosphorylation consécutive des autres sérines et thréonines. La phosphorylation des sérines 328, 332 et 338 est primordiale pour l’internalisation et l’interaction avec la β-arrestine 2 (Potter et al., 2006). Deux domaines de phosphorylation régulent de manière différente les affinités pour la β-arrestine et les agonistes (Key et al.,