Metabolism under the spotlight in senescence
Texte intégral
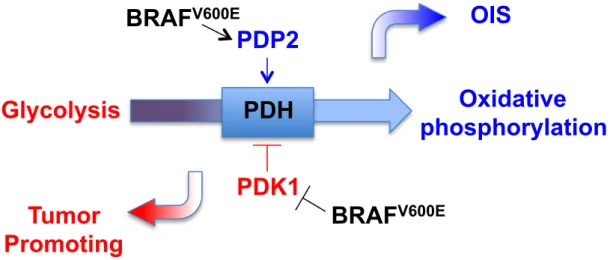
Figure
Documents relatifs
CF= correlation factor between a mitochondrial probe (MitoTracker green) and Bak or Bcl-2, 100 cells were used in each experiment. Bak is cytosolic in siMtx2 treated cells, but not
As we provide molecularly oriented de fi nitions of terms including intrinsic apoptosis, extrinsic apoptosis, mitochondrial permeability transition (MPT)-driven necrosis,
This model parameterizes the BSC with four parameters: the mean scatterer radius a, the gamma width factor ζ, the total volume fraction φ and the relative acoustic impedance contrast.
This paper presents an automatic elbow control method based on natural coordinations between shoulder and elbow kinematics: as healthy individuals would use their upper
3 ) obtained after 1 h of kinetic study in stoichiometric conditions and (b) Structures detected on the spectrum; Figure S10: (a) 1 H NMR spectrum (in CDCl 3 ) obtained after 10 h
Eu égard à leur mode d’action et à leurs effets pluriels (glycémiques/extraglycémiques), les analogues du GLP-1 (GLP-1 RAs pour Glucagon-like-Peptide-1 Receptor Agonists)
• Un triangle est constructible lorsque la longueur de son plus grand côté est strictement inférieure à la somme des longueurs des deux autres côtés. • Si ce n'est pas
In this review, we discuss our current understanding of the molecular machinery of each of the main types of non-apoptotic RCD, including necroptosis, pyroptosis, ferroptosis,
