T
T
H
H
È
È
S
S
E
E
En vue de l'obtention du
D
D
O
O
C
C
T
T
O
O
R
R
A
A
T
T
D
D
E
E
L
L
’
’
U
U
N
N
I
I
V
V
E
E
R
R
S
S
I
I
T
T
É
É
D
D
E
E
T
T
O
O
U
U
L
L
O
O
U
U
S
S
E
E
Délivré par l'Université Toulouse III - Paul SabatierDiscipline ou spécialité : Neurosciences
JURY
Pr. Jacques Haiech, Professeur des Universités (Rapporteur) Dr. Serge Birman, Directeur de Recherche (Rapporteur)
Dr.Yves Le Conte, Directeur de Recherche (Examinateur) Pr. Martin Giurfa, Professeur des Universités (Président du jury) Dr. Valérie Raymond-Delpech, Maître de Conférence (Directrice de thèse)
et Dr. Marc Moreau, Directeur de Recherche (Directeur de thèse)
Ecole doctorale : CLESCO Unité de recherche : UMR5169
Directeur(s) de Thèse : Dr. Valérie Raymond-Delpech et Dr. Marc Moreau Présentée et soutenue par Emmanuel Perisse
Le 20 novembre 2009
Titre : Le calcium, un déclencheur moléculaire de la formation
de la mémoire olfactive à long terme chez l’abeille : approches comportementale et transcriptomique.
Avant-propos
« Le savant n’est pas l’homme qui fournit les vraies réponses, c’est celui qui pose les vraies questions »
Claude Levi-Strauss (1908-2009)
Je souhaite remercier Valérie Raymond-Delpech, Marc Moreau, Jean-Christophe Sandoz mais aussi Isabelle Néant et Catherine Leclerc pour m’avoir choisi et avoir cru en moi pour travailler sur ce projet ambitieux et très intéressant financé grâce à la bourse du président de l’université.
Je tiens à remercier le Professeur Martin Giurfa de m’avoir accepté au sein de son laboratoire déjà en maîtrise, en Master 2 Recherche, puis enfin en thèse, et d’avoir accepté de présider ce jury. Mais surtout, merci Martin pour l’influence scientifique que vous avez eue sur moi : vos enseignements en Licence, en Maîtrise puis en Master 2 Recherche m’ont donné le goût de la cognition. Merci aussi pour votre aide pour la suite de ma carrière scientifique.
Je tiens à remercier Jacques Haiech et Serge Birman pour avoir accepté d’être rapporteurs de mon travail de thèse. Je remercie également Yves Le Conte pour avoir accepté d’examiner ce travail de thèse.
Je remercie très tendrement Stéphanie pour tout ce qu’elle m’a apporté au quotidien depuis ces 3 dernières années. Merci pour ton soutien, ton aide, ta gentillesse, ta générosité et ton amour qui ont fait de moi aujourd’hui le docteur que je suis.
Je remercie très chaleureusement Valérie pour sa gentillesse et sa disponibilité, qui m’ont permis d’avancer sur ce projet et d’évoluer dans mon travail scientifique. Je te remercie pour toutes ces discussions (au travers du plexiglas) autour d’interrogations sur les expériences qui ne marchent pas, ou sur les choix à entreprendre pour avancer sur ce travail. Merci également d’avoir passé du temps en dehors de tes heures de travail pour m’aider à travailler sur différents projets de thèse ou post-thèse.
Je veux exprimer toute ma gratitude et ma reconnaissance à Jean-Christophe pour avoir dirigé mon stage de M2R et mon travail de recherche de thèse pendant ces 4 dernières années. Je te suis très reconnaissant de m’avoir fait partager tes connaissances, ton savoir-faire et ton expérience. Tes qualités ont été selon moi déterminantes et à l’origine de l’avancement rapide de mes travaux de recherche. Je te remercie pour tes conseils, ta patience notamment pour t’avoir dérangé, de temps en temps le soir, 2 minutes et finalement resté 45 minutes pour des discussions scientifiques mémorables.
Je souhaiterais mantenant passer du coté moléculaire de la science et remercier Marc pour tout ce qu’il m’apporté, sur ces connaissances (scientifiques et autres..) qui m’ont permis de grandir scientifiquement. Un grand merci à Isa pour m’avoir appris les secrets de la biomol et pour ta disponibilité. Merci aussi à Catherine pour avoir suppléer Isa dans cette lourde tâche. Merci aussi à tous les trois pour votre bonne humeur, votre disponibilité, votre aide et vos conseils.
Je veux remercier tout particulièrement Jean-Marc avec qui tout à commener en maîtrise. Tu m’as permis de découvrir le monde de la recherche en France mais aussi au travers de voyage vers différents continents. Merci de m’avoir aidé et formé pour que je puisse aujourd’hui, obtenir le statut de docteur. Je remercie également Jean-Marc d’avoir partagé ma découverte de l’escalade, et oui, 7a en un an, c’est grâce à toi.
Un énorme merci à Benoît pour son amitié, son soutien et son aide depuis ces 4 dernières années. Merci pour ta bonne humeur au quotidien et pour ta générosité. Je ne l’oublierai pas. Merci aussi de m’avoir fait découvrir la capoeira.
Un grand merci à Claire pour ses conseils précieux, pour m’avoir aidé pour ma thèse et surtout pour la suite de ma carrière. Je te suis très reconnaissant.
Je ne peux pas oublier non plus les personnes qui m’ont aidé pour une partie des expériences menées dans cette thèse : Kevin, Yuki, Lisa, Pascal, Lidwine, Laure, Quentin et Fred.
Je veux finir remercier toutes les personnes qui ont partagé mon quotidien et que j’apprécie vraiment beaucoup : Laure, Alex, Edith, Julien, Julie, Meike, Lionel, Théo, Ambre, Audrey, Medhi, Abel, Geoffrey (même si t’es parti, on a partagé beaucoup de choses depuis la première année de fac), Monique, Hélène, Isa, Gérard, Patrick, Stéphane, Cathy, Bernard, Michel et Sandrine.
Je veux dédier cette thèse à ma famille qui m’a aidé, supporté et encouragé pendant ces longues années. Merci pour tout ce que vous m’avez apporté.
Publications et communications
Travaux de thèse publiés (revues internationales à comité de lecture):
Perisse E, Raymond-Delpech V, Neant I, Matsumoto Y, Leclerc C, Moreau M and Sandoz JC (2009)
Early calcium increase triggers the formation of the olfactory long-term memory in honeybees. BMC
Biol. 16;7(1):30.
Travaux de thèse en cours de publication :
Perisse E, Neant I, Sandoz JC, Moreau M, Leclerc C and Raymond-Delpech V. Calcium-regulated
gene expression involved in the olfactory long-term memory formation in honeybees.
Travaux de thèse présentés lors de congrès scientifiques nationaux ou internationaux:
Perisse E, Sandoz JC, Neant I, Leclerc C, Moreau M and Raymond-Delpech V. (Juin 2009) What can
we learn from honeybees? The role of calcium in long-term memory formation in honeybees: a new model to study neurodegenerative diseases. ECS workshop: Annexin target and calcium binding protein in pathology. Smolenice, Slovaquie.
Perisse E. (Juillet 2009). Long-term memory and plasticity in honeybee: structural and molecular
approaches. Invité par Scott Waddell, Departement de Neurobiologie, Université du Massachussets, Worcester, USA.
Perisse E. (Mai 2008). Calcium and gene expression involved in the olfactory long-term memory in
the honeybee Apis mellifera. 9ème rencontre du Club de Neurobiologie des Invertébrés, Toulouse, France.
Perisse E. (Mars 2008). Role of calcium in the olfactory long-term memory in the honeybee Apis
mellifera. Conférences Alpha-T, Toulouse, France.
Perisse E. (Septembre 2007).Calcium and gene expression involved in the olfactory long-term
Table des matières
I.
Introduction Générale ... 15
I.1.
Partie I : la formation de la mémoire à long terme... 15
I.1.1. La mémoire : un processus psycho-biologique complexe. ...15
a. Mémoire à court terme et mémoire à long terme : le mécanisme de consolidation mnésique...15
b. Le conditionnement classique comme modèle d’apprentissage pour l’étude des bases cellulaires et moléculaires de la formation de la mémoire à long terme ...17
c. Les modèles animaux utilisés pour l’étude des bases cellulaires et moléculaires de la formation de la mémoire à long terme...18
I.1.2. Les bases cellulaires et moléculaires sous-tendant la formation de la mémoire à long terme...19
a. Les bases moléculaires de la formation de la mémoire à long terme ...20
b. Synthèse protéique et mémoire à long terme...26
c. Mémoire à long terme et modifications de l’efficacité synaptique : le phénomène de potentialisation à long terme...28
d. Mémoire à long terme et modifications de l’architecture synaptique ...30
I.2.
Partie II. Le calcium, un acteur clé des fonctions nerveuses... 32
I.2.1. Le calcium et les fonctions neuronales...32
I.2.2. Quelles sont les sources de calcium ? ...33
a. L’entrée de calcium extracellulaire ...33
• Les CCVD (canaux calciques voltage dépendants)...33
• Les canaux calciques chimio-dépendant (CCCD) ou ROC (« receptor operated channel ») ...34
• Les canaux SOC, CRAC et SMOC ...35
• Les canaux TRP (« transient receptor potential ») ...35
b. La libération de calcium à partir des stocks intracellulaires...38
• Le réticulum endoplasmique ...38
Les récepteurs à l’IP3 (IP3R)...39
Les récepteurs à la ryanodine (RyR) ...40
• Le réticulum endoplasmique et la génération de signaux calciques complexes 40 • Les mitochondries ...41
• L’enveloppe nucléaire ...42
I.2.3. Régulation de l’homéostasie calcique ...42
b. Les échangeurs sodium/calcium (Na+/Ca2+)...43
c. Le calcium, un acteur clé de la plasticité synaptique ...43
d. Calcium et régulation de la transcription...44
• Le calcium régule l’activation de facteurs de transcription...44
• Les cibles de l’activation de facteurs de transcription par le calcium ...48
La transcription régulée par CREB ...48
La transcription régulée par NF-κB...49
La transcription régulée par NFAT ...49
La transcription régulée par DREAM ...50
La transcription régulée par SRF...51
I.2.4. Le dialogue synapto-nucléaire par le calcium...52
I.2.5. Calcium et formation de la mémoire à long terme ...53
I.3.
Partie III. L’abeille Apis mellifera, modèle neurobiologique des
bases comportementales, cellulaires et moléculaires de la formation de la
mémoire. ... 55
I.3.1. Le conditionnement classique appétitif Pavlovien du réflexe d’extension du proboscis. ...56
I.3.2. Les différentes phases de mémoire chez l’abeille ...57
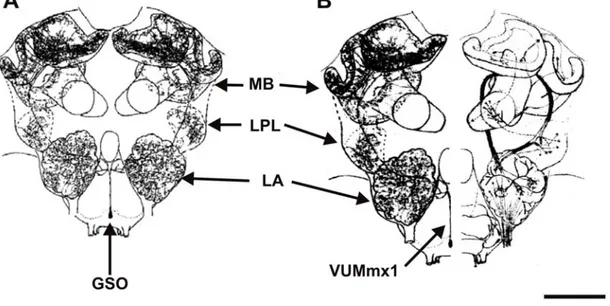
I.3.3. Les bases neurales de la formation de la mémoire olfactive chez l’abeille...60
a. Architecture du cerveau de l’abeille...60
b. Traitement de l’information olfactive...61
c. Traitement de l’information gustative : la voie du renforcement lors du conditionnement du PER ...64
I.3.4. Le cerveau de l’abeille, siège d’une plasticité associée à l’expérience ou à l’apprentissage ...65
I.3.5. Les bases moléculaires de la formation de la mémoire chez l’abeille. ...68
II.
Objectifs des travaux de thèse ... 69
III.
Matériels et Méthodes ... 71
III.1.
Partie 1 : Etudes comportementales... 71
III.1.1. Animaux ...71
III.1.2. Conditionnement olfactif ...72
a. Dispositif ...72
c. Test de rétention ...73
d. Odeurs utilisées pour le conditionnement ...74
III.1.3. Agents pharmacologiques utilisés pour le comportement ...74
a. Modes d’injection des agents pharmacologiques ...74
b. Contrôle de l’inhibition de l’augmentation de la [Ca2+]i...75
c. Contrôle de l’augmentation de la [Ca2+]i ...75
d. Inhibition de la synthèse protéique de novo ...77
e. Préparation des solutions ...77
f. Composés possédants un groupement acétoxy-méthyl (AM) ...77
III.1.4. Photolyse du produit encagé...77
a. Préparation des abeilles ...77
b. Le dispositif ...79
c. La photolyse ...79
III.1.5. Imagerie Calcique...80
a. Préparation des abeilles ...80
b. Procédure d’enregistrement des signaux calciques ...82
c. Analyse des données d’imagerie calcique ...84
III.1.6. Analyse statistique des données...84
III.2.
Partie 2 : Etude transcriptomique... 85
III.2.1. Animaux et procédure expérimentale...85
III.2.2. Sélection des abeilles...87
III.2.3. Expérience utilisant les puces à ADN ...88
a. Extraction des ARN totaux...89
b. Marquage des ARNm (ARN messagers)...90
c. Hybridation...91
d. Scan des puces à ADN hybridées ...92
e. Analyse d’images des scans de puces à ADN ...92
f. Analyse statistique des résultats de puces à ADN...93
III.2.4. Transcription inverse et réaction de polymérisation en chaîne (RT-PCR) quantitative ...93
a. Sélection des gènes candidats...93
b. Choix des gènes de références...94
c. Choix des amorces...94
d. Efficacité des couples d’amorces choisis ...95
e. Quantification relative des gènes candidats...97
IV.
Résultats ... 99
IV.1.
Chapitre 1 : Rôle précoce du calcium dans la formation de la
mémoire olfactive à long terme chez l’abeille. ... 99
IV.1.1. Introduction ...100
IV.1.2. Résultats ...101
a. L’augmentation du niveau de calcium intracellulaire pendant l’apprentissage est nécessaire pour la formation de la mémoire olfactive à long terme. ...101
b. L’augmentation du niveau de calcium intracellulaire pendant l’apprentissage est suffisante pour la formation d’une mémoire olfactive à long terme dépendante de la synthèse protéique. ...106
IV.2.
Chapitre 1 : points importants à retenir... 117
IV.3.
Chapitre 2 : Criblage des gènes dont l’expression est dépendante
du calcium et impliqués dans la formation de la mémoire olfactive à long
terme chez l’abeille. ... 119
IV.3.1. Introduction ...120
IV.3.2. Résultats ...120
a. Criblage transcriptomique ...120
b. RT-PCR quantitative ...125
IV.4.
Chapitre 2 : points importants à retenir... 131
V.
Discussion ... 133
V.1.
Le rôle du calcium dans les étapes précoces de la consolidation ..
133
V.2.
La spécificité de l’effet du calcium sur la mémoire à long terme..
138
V.3.
La spécificité de la mémoire à long terme formée
... 138
V.4.
Les sources de calcium impliquées dans la formation de la mémoire
à long terme
... 139
V.6.
Les gènes impliqués dans la formation de la mémoire à long
terme ……….144
V.7.
L’expression de gènes dépendant du calcium et impliqués dans la
formation de la mémoire à long terme... 146
VI.
Conclusions et perspectives ... 149
VII.
Références ... 155
VIII. Liste des figures ... 179
Abbréviations
AC : adénylate cyclase
ADP-RT : ADP-ribosyl-tranferase AM : acétoxy-méthyl
AMPA : α-amino-3-hydroxy-5-méthylisoazol-4-propionate AMPc : adénosine monophosphate cyclique
ARCs : arachidonate-activated channels ARNm: ARN messagers
Atf-1: activating transcription factor-1 ATP : adénosine tri-phosphate
BAPTA-AM : 1,2 - bis - (o - Aminophenoxy) - ethane - N,N,N',N' - tetraacetic acid,
tetraacetoxymethyl ester
BDNF: brain-derived neurotrophic factor BLAST: basic local alignment sequence tool Ca2+: calcium
[Ca2+]i : concentration intracellulaire de calcium Ca2+/CaM: Calcium/Calmoduline
CaBPs : calcium-binding protein
CaMKs : calmoduline-dependent protein kinases
CaV: canal d’ion calcium physiologiquement régulé par le voltage
CBP: CRE Binding Protein
CCAT : calcium channel-associated transcription CCCD : canaux calciques chimio-dépendant CICR: calcium-induce calcium release CCVD : canaux calciques voltage dépendant CIRB: calmoduline/IP3 binding domain CRAC: calcium release-activated calcium
CREB : cAMP responses element binding Crem : cAMP response element modulator DAG : diacylglycérol
DLT : dépression à long terme DMSO : diméthylsulfoxyde
DREAM: downstream response element (DRE) antagonist modulator ERK: extracellular-signal regulated kinases
GC : guanylate cyclase
HATs : histones acétyl-transférases IEGs: immediatly early genes IκB : inhibitory κB
IP3R : récepteur à l’inositol tri-phosphate
JNK/SAPK: jun N-terminal kinase/stress-activated protein kinase KChIP3 : potassium-channel-interacting protein 3
l-ACT : lateral antenno-cerebral tract m-ACT: medial antenno-cerebral tract MAPK: mitogenes-activated protein kinases mGluR : récepteurs métabotropiques au glutamate MCT : mémoire court terme
MLT : mémoire à long terme MMT : mémoire à moyen terme NFAT : nuclear factor activated T-cells NF-κB: nuclear factor-kappa B
NMDA : N-methyl-D-aspartate NO : monoxyde d’azote
NP-EGTA-AM
:O-Nitrophényle-éthylèneglycol-bis(β-aminoéthyle)-N,N,N’,N’-tétraacétoxyméthyle-ester
NS: non significatif
PBS : Phosphate Buffer Saline
PCMA : plasmic membrane calcium ATPases PIP2 : phosphatidylinositol 4,5 biphosphate PKA : protéine kinase A
PLC : phospholipase C
PLT : potentialisation à long terme RC : Réponse Conditionnée
RCPG : récepteurs couplés aux protéines G RE : réticulum endoplasmique
ROC: receptor operated channel
REB : cAMP responses element binding REP : réflexe d’extension du proboscis RNAsin : RiboLockTM RNase Inhibitor
RS : réponse spécifique
RyR : récepteurs à la ryanodine SC : stimulus conditionnel
SERCA : sarco-endoplasmic reticulum calcium-ATPases SI : stimulus inconditionnel
SMOCs: second messenger operated channels SOCs : store operated channels
SRF : serum response factor TCF : ternary complex factor
TRAP: Translating Ribosome Affinity Purification TRP: transient receptor potential
UV : rayonnement ultraviolet VOC: voltage operated channel
I.
Introduction Générale
I.1.
Partie I : la formation de la mémoire à long terme
I.1.1.
La mémoire : un processus psycho-biologique complexe.
La mémoire peut être définie comme un processus nous permettant d’acquérir et de conserver certaines informations sensorielles perçues, puis de les restituer ultérieurement. Elle permet à l’individu de survivre en adaptant son comportement aux modifications de l’environnement, et détient ainsi une valeur adaptative importante. L’une des plus extraordinaires propriétés du cerveau réside dans sa plasticité liée à l’expérience. Les études menées aussi bien sur les invertébrés que sur les vertébrés ont grandement contribué à l’amélioration de la compréhension des mécanismes qui sous-tendent cette plasticité cérébrale (Bailey et kandel, 1993 ; Kolb et Whishaw, 1998 ; Kandel, 2001 ; Dubnau et al., 2003a). Depuis plusieurs siècles et au travers de différentes disciplines (de la philosophie à la neurobiologie), la compréhension du fonctionnement de la mémoire n’a de cesse de fasciner les chercheurs. Les principales avancées sont essentiellement issues des travaux de neurobiologie combinant les approches comportementales venues de la psychologie expérimentale avec les approches d’anatomie, d’électrophysiologie, de pharmacologie, de biologie moléculaire, de biochimie et de génétique (Rudy, 2008).
L’ensemble des travaux relatifs à l’étude des mécanismes de la mémoire a conduit à la mise en évidence de deux types de mémoires distinctes au niveau temporel, aussi bien chez l’Homme que chez l’animal, qui sont la mémoire à court terme et la mémoire à long terme.
Les deux objectifs majeurs de ce manuscrit de thèse sont de déchiffrer et de comprendre, (1) quel pourrait être le déclencheur moléculaire de la formation de la mémoire à long terme, et (2) comment pourrait être régulée, via ce déclencheur moléculaire, l’expression de gènes requis pour la formation de cette mémoire.
a. Mémoire à court terme et mémoire à long terme : le mécanisme de consolidation mnésique
Au début du vingtième siècle, les travaux d’Hermann Ebbinghaus ont permis de mettre en évidence les deux principales caractéristiques du stockage mnésique. Il montra que les souvenirs ont des durées de vie différentes et que le processus de répétition rend les souvenirs plus durables (Ebbinghaus, 1913). En effet, certains souvenirs, de courtes durées, sont retenus pendant quelques heures ; d’autres, de longue durée, peuvent persister pendant des jours, des mois voire des années. Quelques années plus tard, les
psychologues allemands Georg Müller et Alfons Pilzeckzer suggéraient que cette mémoire, qui peut durer des jours voire des semaines, se consolide avec le temps, et ont donc évoqué pour la première fois le processus de consolidation mnésique, qui correspond à la période de stockage des informations à long terme (Müller et Pilzecker, 1900 ; Lechner et al., 1999). Pendant cette période, la mémoire est sensible aux perturbations, mais une fois que le souvenir est consolidé, il apparaît alors robuste et insensible aux interférences. Grâce à ces travaux, le philosophe et physiologiste William James (1890) proposa une distinction qualitative pertinente entre la mémoire primaire (mémoire à court terme) et la mémoire secondaire (mémoire à long terme). Il montra que la mémoire à court terme dure quelques minutes et est essentiellement une extension du moment présent. A l’inverse, la mémoire à long terme intervient dans la recherche du passé. Cette distinction s’est avérée fondamentale pour la compréhension des processus mnésiques.
Ainsi, on peut définir le processus de mémorisation en trois étapes clés (figure 1) : la première est la phase d’acquisition, qui permet la détection et l’encodage de stimuli informatifs pertinents. Elle relève de différentes modalités sensorielles, comme la vision, l’audition, l’olfaction, etc. Ces informations sont ensuite assemblées et transformées, via le processus d’encodage, en une représentation mentale interne cohérente. Ensuite vient la phase de stockage ou de consolidation, permettant la conservation de ces informations au sein du système nerveux central. La durée de cette phase est très variable, de l’ordre de la seconde à toute une vie, et dépend du type d’apprentissage (expérience unique ou répétée) et de l’impact émotionnel de l’information acquise. Enfin, la phase de rappel correspond à la restitution différée de ces informations, permettant ainsi l’expression d’un comportement modifié par une expérience antérieure.
A l’heure actuelle, on ne peut affirmer que les différents processus permettant la formation d’une mémoire à long terme se déroulent de façon strictement séquentielle, comme cela a été décrit dans le modèle d’Atkinson et Shiffrin (1968). Il n’est pas exclu que ces processus se déroulent de façon parallèle ou une combinaison « séquentielle-parallèle » (figure 1). Même s’il existe des limites au modèle d’Atkinson et Shiffrin (Van der Linden, 1995), ce modèle demeure toujours d’actualité et permet aujourd’hui encore d’appréhender les différentes phases du processus de mémorisation, à savoir l’acquisition, le stockage et le rappel. Une question cruciale demeure cependant encore en suspens et concerne la nature et la séquence des mécanismes permettant la conservation des informations perçues.
Figure 1. Modèle des bases comportementales, cellulaires et moléculaires de la formation de la mémoire à long terme. On peut noter la possibilité d’un traitement séquentiel et/ou parallèle de la formation de la mémoire à long
terme – par l’intermédiaire ou non de la formation de la mémoire à court terme. Les trois principales étapes de la formation de la mémoire à long terme sont : l’acquisition, le stockage et le rappel de l’information. Chacune de ces étapes est supportée par des événements cellulaires et moléculaires différents. La stabilisation à long terme d’une information au sein du cerveau requiert l’étape de consolidation. Ainsi, l’information stockée et stabilisée devient insensible à l’interférence.
b. Le conditionnement classique comme modèle d’apprentissage pour l’étude des bases cellulaires et moléculaires de la formation de la mémoire à long terme
La formation et le stockage à long terme d’une information nécessitent au préalable un apprentissage. Il existe différents types d’apprentissages plus ou moins complexes au cours desquels la quantité et la qualité des informations acquises sont différentes. Avec l’essor des recherches en neurobiologie, les travaux pionniers concernant l’étude des bases cellulaires et moléculaires de la formation de la mémoire ont été entrepris de manière réductionniste, basés sur des modèles d’apprentissage simple (apprentissage associatif, comme le conditionnement classique, et apprentissage non associatif, comme l’habituation et la sensibilisation), et sur des animaux dont le système nerveux était peu complexe comme celui de l’aplysie, une limace de mer (pour revue, Kandel, 2001).
Le conditionnement associatif classique permet un apprentissage élémentaire simple, robuste et précis, que l’on peut étudier sur différents modèles animaux, vertébrés ou invertébrés. Au niveau comportemental, ce type de conditionnement a été mis en évidence pour la première fois par Ivan Pavlov (1927), qui étudiait les réflexes digestifs chez le chien. Ce conditionnement consiste en l’association de deux stimuli, un stimulus
neutre qui devient conditionnel (SC) et un stimulus inconditionnel (SI) (figure 2). Le stimulus neutre n’entraîne à lui seul aucune réponse comportementale de la part de l’animal. En revanche, s’il est associé selon un appariement antérograde (le SC précède le SI), le SC peut ensuite déclencher une réponse comportementale appelée réponse conditionnée (RC).
Figure 2. Le conditionnement classique pavlovien. 1. Avant le conditionnement, la nourriture (le
stimulus inconditionnel) entraîne une réponse réflexe (réponse inconditionnée) de salivation du chien. 2. En revanche, la présentation du son par le diapason (le stimulus neutre) ne déclenche aucune réponse de la part de l’animal. 3. Pendant le conditionnement, on présente tout d’abord le son (devenant stimulus conditionnel) qui va être rapidement associé à la présentation de la nourriture. Cette association entraîne alors une réponse de salivation du chien, le chien apprenant l’appariement temporel entre ces deux stimuli.
4. Ainsi, après le conditionnement, et si le chien a bien appris l’association temporelle (le son précède
l’arrivée de nourriture), la présentation du stimulus conditionnel (le son du diapason) entraîne cette fois-ci une réponse conditionnée du chien : il salive.
c. Les modèles animaux utilisés pour l’étude des bases cellulaires et moléculaires de la formation de la mémoire à long terme
Pendant les années 50 et 60, de nombreux biologistes et psychologues considéraient que l’apprentissage, et donc le comportement, était le seul domaine de la biologie pour lequel l’utilisation d’animaux, tels les modèles invertébrés, n’avait que peu de chance d’apporter des résultats concluants. Cependant, malgré le développement de nouvelles techniques (comme la biologie moléculaire, l’électrophysiologie et la génétique), l’étude des bases cellulaires et moléculaires de la formation de la mémoire au sein de système nerveux aussi complexe que celui du mammifère, s’est révélée particulièrement difficile.
Sachant que des formes élémentaires et non élémentaires d’apprentissage sont communes à tout animal possédant un système nerveux (pour revue, Kandel, 2001), il doit exister, au sein des mécanismes d’apprentissage, des caractéristiques cellulaire et moléculaire communes et conservées entre les vertébrés et les invertébrés. Ainsi, Eric Kandel s’est intéressé à l’aplysie Aplysia californica, pour entreprendre l’étude des bases cellulaires et moléculaires de la formation de la mémoire. La relative simplicité du système nerveux de ce modèle, par rapport à celui des mammifères, a permis à Kandel et ses collaborateurs de faire des découvertes majeures sur l’identification des réseaux neuronaux et le rôle de certaines molécules impliquées dans la formation de la mémoire (pour revue, Kandel, 2001). Le modèle de la drosophile a quant à lui été utilisé pour démontrer génétiquement le rôle nécessaire et suffisant de certaines structures de son cerveau et de certaines molécules dans les processus mnésiques (Davis, 2005 ; McGuire et al., 2005 ; Keene et Waddell, 2007). Ce modèle présente cependant des inconvénients importants à considérer, tels que la petite taille de son cerveau qui le rend difficile d’accès, ainsi que les paradigmes comportementaux utilisés pour l’étude de la formation de la mémoire qui ne sont pas réalisés de façon individuelle. Dans ce contexte, depuis une vingtaine d’années, l’abeille domestique Apis mellifera s’est révélée être un modèle approprié pour l’étude des bases comportementales, cellulaires et moléculaires de l’apprentissage et de la mémoire (Menzel et Müller, 1996 ; Menzel, 2001 ; Giurfa, 2007). En effet, son cerveau est facilement accessible et bien décrit au niveau anatomique. De plus, le génome de l’abeille a été récemment séquencé (The Honeybee Genome Sequencing Consortium, 2006) et les paradigmes comportementaux utilisés pour étudier la formation de la mémoire sont réalisés au niveau individuel (voir I.3 partieIII).
I.1.2.
Les bases cellulaires et moléculaires sous-tendant la formation
de la mémoire à long terme
D’un point de vue neurobiologique, il est généralement admis que la formation et le stockage à long terme des souvenirs s’accompagnent de modifications durables de l’efficacité de la transmission synaptique et de l’architecture des réseaux neuronaux activés au cours de l’apprentissage (pour revues, Bailey et Kandel, 1993 ; McGaugh, 2000 ; Kandel, 2001 ; Dudai, 2004 ; Lynch, 2004). Cette relation entre les processus mnésiques et la plasticité des structures cérébrales est connue depuis la fin du XIXème siècle grâce aux travaux de Ramon y Cajal (1894) et Camillo Golgi (1873). Ces auteurs ont émis l’hypothèse selon laquelle l’apprentissage conduirait à des modifications structurales des synapses permettant de stocker l’information. Ces travaux constituent les fondements des neurosciences modernes et ont permis à Donald Hebb de formuler les concepts élémentaires appelés aujourd’hui les « lois de Hebb » (Hebb, 1949). Selon lui, l’activité neuronale pourrait induire un renforcement des connexions synaptiques
pré-existantes ainsi qu’une synaptogenèse. Jerzy Konorski (1948) proposa une hypothèse complémentaire : la plasticité synaptique, induite par l’association de stimuli répétés, pourrait transformer les connexions synaptiques potentielles en connexions synaptiques fonctionnelles. Aujourd’hui, les théories de « Hebb-Konorski » constituent les fondements essentiels de la plupart des modèles neurobiologiques de la mémoire.
Ces modifications synaptiques seraient donc le siège du stockage de l’information à long terme et mettraient donc un certain temps à se mettre en place (Bailey et Kandel, 1993). En effet, les changements synaptiques à court terme mettent en jeu des modifications de protéines pré-existantes conduisant à des modifications de connexions synaptiques elles-aussi pré-existantes. La phase de mise en place de ces modifications est assez courte, mais au cours de celle-ci, les connexions sont sensibles à l’interférence : on parle alors de trace mnésique fragile. Cependant, afin qu’une information soit maintenue à long terme et pour éviter que le souvenir s’efface, des changements synaptiques à long terme semblent nécessaires. Ces changements impliquent l’expression de nouveaux gènes, une synthèse protéique de novo, et la formation de nouvelles connexions. Ces changements, qui nécessitent du temps, vont permettre de stabiliser les connexions d’un réseau précis de neurones au sein duquel l’information va être stockée à long terme. La trace mnésique sera donc stabilisée au sein de ce réseau de neurones (pour revue, Kandel, 2001 ; Laroche, 2006).
a. Les bases moléculaires de la formation de la mémoire à long terme
Dans les années 70, Kandel et ses collaborateurs ont étudié la formation de la mémoire à court terme et à long terme au niveau comportemental, puis cellulaire et enfin moléculaire. Ces études ont été réalisées sur un modèle simple, l’aplysie, en utlisant des apprentissages simples tels que l’habituation, la sensibilisation ou le conditionnement classique. Par la suite, les études sur la plasticité synaptique chez l’aplysie et chez le mammifère, ainsi que des études génétiques chez la drosophile, ont permis de nombreuses avancées sur le rôle des molécules impliquées dans la formation de la mémoire. Nous essaierons dans cette partie d’en récapituler les caractéristiques essentielles et les travaux majeurs qui ont permis d’arriver aujourd’hui au concept suivant: la consolidation mnésique repose sur des modifications durables de la connectique et de l’efficacité des synapses (plasticité neuronale à long terme). Les modifications cellulaires sous-jacentes dépendent de régulations géniques, de modifications post-traductionnelles et d’une néo-synthèse de protéines. Tout ceci serait lié à l’activation de nombreuses molécules et de voies moléculaires, de façon conservée chez différents modèles allant des vertébrés aux invertébrés (pour revues, Bailey et al., 1996 ; Kandel, 2001 ; Wang et al., 2006 ; Hawkins et al., 2006 ; Izquierdo et al., 2006 ; Keene et Waddell, 2007) (figure 3).
L’adénylate cyclase (AC)
Parmi ces molécules, la plus étudiée est l’adénylate cyclase (AC) associée à la voie adénosine monophosphate cyclique - protéine kinase A (AMPc-PKA). En effet, elle est l’une des premières voies de seconds messagers à avoir été étudiée par Kandel. Un grand nombre de travaux chez différents modèles animaux, a montré l’importance des molécules de cette voie dans la formation de la mémoire (Livigstone et al., 1984 ; Schacher et al., 1988 ; Skoulakis et al., 1993 ; Mons et al., 1999 ; Müller, 2000 ; Sutton et Carew, 2000 ; Schwärzel et Müller, 2006 ; Matsumoto et al., 2006 ; Michel et al., 2008).
L’AC catalyse la conversion de l’adénosine tri-phosphate (ATP) en AMPc, un second messager qui présente divers rôles régulateurs au sein du système nerveux, et qui est nécessaire à l’activation de la PKA. L’activité de l’AC est dépendante du niveau de calcium intracellulaire (Ferguson et Storm, 2004). L’AC est aussi un détecteur de coïncidence capable d’intégrer des informations de deux sources indépendantes (figure
3) comme lors d’un apprentissage Pavlovien associant deux stimuli (Davis et al., 1995).
Chez les mammifères, il existe deux types d’adénylate cyclase fortement impliquées dans des processus neuronaux complexes comme la formation de la mémoire à long terme, l’AC1 et l’AC8 (pour revue, Ferguson et Storm, 2004). Cela est lié à son produit d’activation, l’AMPc, qui active différentes molécules comme la PKA. La PKA est composée de deux sous-unités, une régulatrice et une catalytique. L’AMPc produite par l’activation de l’AC se fixe sur la sous-unité régulatrice de la PKA. La sous-unité catalytique est alors libérée et transloquée dans le noyau afin de phosphoryler des facteurs de transcription (Poser et Storm, 2001) (figure 3). Chez la drosophile et chez l’aplysie, un rôle similaire a aussi été montré, indiquant une remarquable conservation de ces processus au cours de l’évolution (pour revue, Bailey et al., 1996). En effet, des mutants chez la drosophile comme rutabaga, dunce ou DCO, des souris Knock-out (génétiquement modifié pour inactiver un gène), ou des animaux injectés avec des inhibiteurs pharmacologiques, ont permis de montrer que des perturbations de l’AC ou de la voie AMPc-PKA entraînent des déficits mnésiques importants (Tully et Quinn, 1985 ; Zhao et al., 1995 ; Müller, 2000 ; Zars et al., 2000 ; Ferguson et Storm, 2004 ; Hawkins
et al., 2006 ; Michel et al, 2008). Enfin, l’activation de la voie AC-AMPc-PKA peut être
amplifiée par le complexe calcium/calmoduline qui va interagir avec l’AC (figure 3).
Les calmodulines Kinases (CaMK)
Une autre voie très étudiée est la voie des calmodulines kinases
(calmoduline-dependent protein kinases, CaMKs) avec notamment le rôle de la CaMKII (Mayford et al., 1996 ; Micheau et Riedel, 1999 ; Irvine et al., 2006 ; Cammarota et al., 2008 ;
post-synaptique (Kennedy et al., 1983). Cette enzyme, qui s’autophosphoryle grâce au complexe calcium/Calmoduline (Ca2+/CaM) suite à une élévation de la concentration intracellulaire de calcium [Ca2+]i, est capable, au moins in vitro, de décoder les oscillations calciques intracellulaires par modifications de l’affinité de ses sites de phosphorylations. Ces caractéristiques suggèrent que cette enzyme serait responsable de l'amplification des signaux d’activation cellulaire (Dupont et Goldbeter, 1998 ; Dupont et
al., 2003 ; Cammarota et al., 2008). La CaMKII, activée suite à une augmentation de la
[Ca2+]i et une activation du complexe Ca2+/CaM, va pouvoir interagir avec certains récepteurs glutamatergiques chez le rongeur (comme les récepteurs NMDA
(N-methyl-D-aspartate) et AMPA (α-amino-3-hydroxy-5-méthylisoazol-4-propionate)). Cette interaction va permettre d’augmenter le flux ionique à travers ces canaux et ainsi d’amplifier le signal calcique (Soderling, 1993 ; Derkach et al., 1999). La CaMKII peut aussi être transloquée dans le noyau, après son activation, pour ensuite activer des facteurs de transcription et permettre la synthèse de nouvelles protéines nécessaires à la formation de la mémoire (figure 3) (Matthews et al., 1994 ; Greer et Greenberg, 2008).
Figure 3. Représentation schématique des principales voies de signalisation impliquées dans la formation de la mémoire. L’arrivée d’un potentiel d’action au niveau de la terminaison pré-synaptique (1)
entraîne l’entrée de calcium (Ca2+) qui permet l’exocytose de neurotransmetteurs qui vont activer des récepteurs spécifiques au niveau de l’élément post-synaptique (3). Les récepteurs activés vont permettre l’entrée de calcium et d’autres ions qui entraîneront la dépolarisation de la membrane post-synaptique et l’ouverture de canaux dépendant du voltage menant à une plus grande entrée de calcium. Ce calcium va activer différentes voies de signalisation moléculaire comme la voie des MAPK (Raf1-MEK, ERK), la voie des CamKs (Calcium/Calmoduline (Ca2+/CaM), CaMKII), la voie adénosine monophosphate-protéine kinase A (AMPc-PKA) par l’intermédiaire de l’activation de l’adénylate cyclase (AC), et la voie du monoxyde d’azote (NO). Le NO formé sous l’action de la NO synthase (NOS) activée par la Ca2+/CaM, diffuse dans l’élément pré-synaptique (1) active la guanylate cyclase (GC) et l’ADP-ribosyl-tranférase (ADP-RT) entraînant une augmentation de l’exocytose des vésicules de neurotransmetteurs dans la fente synaptique. Un neurone modulateur (2) va quant à lui activer, par libération de neurotransmetteurs, des récepteurs couplés aux protéines G pour activer à son tour l’AC. Ces différentes voies de signalisation vont activer des facteurs de transcription comme CREB (cAMP responses element binding) permettant l’expression de gènes cibles. Le calcium peut aussi activer la transcription plus directement en agissant sur l’activation de co-facteur transcriptionnel comme CBP (CRE binding protein). Cette activation est due à une augmentation de la [Ca2+] nucléaire par l’intermédiaire d’une libération du calcium des stocks du réticulum endoplasmique (RE) et/ou par un autre mécanisme encore mal connu. L’activation de CREB va permettre la transcription de gènes cibles et la synthèse de nouvelles protéines requises pour la formation de la mémoire à long terme.
Les MAPK
La voie des MAPK (mitogenes-activated protein kinases) pourrait être impliquée dans la formation de la mémoire chez les vertébrés (Atkins et al., 1998 ; Micheau et Riedel, 1999 ; Schafe et al., 1999 ; Sweatt, 2004 ; Cammarota et al., 2008) et les invertébrés (Sharma et Carew, 2004 ; Feld et al., 2005 ; Moressis et al., 2009 ; Shobe et al., 2009). La voie des MAPK est généralement activée par la liaison de ligands au niveau de divers récepteurs (figure 3). L’activation de ces récepteurs conduit à l’activation de la protéine ERK (extracellular-signal regulated kinases) qui va être transloquée dans le noyau pour réguler l’expression de gènes cibles. Lors d’un apprentissage, ces gènes pourraient être nécessaires aux modifications synaptiques permettant le stockage à long terme des souvenirs, comme la stabilisation des épines dendritiques, la modulation de récepteurs ioniques ou l’insertion de récepteurs à la membrane (pour revue, Sweatt, 2004).
Le monoxyde d’azote (NO)
Le monoxyde d’azote (nitric oxide, NO) est un gaz dont le rôle dans la formation de la mémoire et de la plasticité synaptique a été montré chez de nombreux modèles animaux (Garthwaite et Boulton 1995 ; Müller, 1996 ; Kendrick et al., 1997 ; Antonov et al., 2007). Une entrée de calcium massive au niveau post-synaptique va permettre l’activation de l’enzyme de synthèse du NO (la NO synthase, NOS) et par conséquent la production du NO. Le NO est un gaz de très courte durée de vie (quelques secondes) qui va pouvoir traverser la membrane plasmique et passer de l’élément post-synaptique à l’élément pré-synaptique. A ce niveau, il va interagir avec la guanylate cyclase et l’ADP-ribosyl-tranferase entraînant une augmentation de l’exocytose des vésicules de neurotransmetteurs dans la fente synaptique (figure 3). Le signal sera donc potentialisé et amplifié par le NO permettant une importante activation de l’élément post-synaptique, et donc des voies de signalisations qui vont mener à la régulation de l’expression de gènes et de la synthèse protéique (pour revue, Prast et Philippu, 2001).
Les cascades d’activation de protéines en chaînes citées précédemment peuvent durer plusieurs dizaines de minutes ; elles constituent une première phase temporelle dans les processus de conversion de la mémoire à court terme en mémoire à long terme. Le stockage à long terme va nécessiter la régulation d’une nouvelle transcription et d’une synthèse protéique par l’activation de facteurs de transcription. Cette deuxième phase nécessitera quant à elle plusieurs heures au cours desquelles l’information se stabilisera par diverses modifications synaptiques au sein d’un réseau de neurones.
CREB («cAMP responses element binding »)
Le facteur de transcription CREB possède un rôle charnière dans le passage d’une mémoire à court terme vers une mémoire à long terme (pour revues, Bailey et al., 1996 ;
Silva et al., 1998 ; Perazzona et al., 2004 ; Alberini, 2009). La molécule CREB est présente de façon constitutive dans tous les types cellulaires du cerveau (Carlezon et al., 2005). Elle est activée par différents types de signaux extracellulaires, et est impliquée dans un nombre important et varié de fonctions cellulaires (Carlezon et al., 2005). Néanmoins, CREB est requis pour la transcription nécessaire à la formation de la mémoire à long terme. Il peut être activé par les différentes molécules des différentes voies citées précédemment (figure 3) (à l’exception du NO). Par exemple, la CaMKII et la CaMKIV (non citée précédemment) activées par le calcium nucléaire et cytoplasmique vont phosphoryler CREB et permettre la transcription de gènes cibles (Silva et al., 1998 ; Alberini, 2009). CREB a donc un rôle central d’intégration des différents signaux d’activation. Il peut donc y répondre en régulant la transcription de gènes cibles et, par exemple, dans le cas de la formation de la mémoire à long terme, permettre la synthèse de nouvelles protéines nécessaires à des modifications synaptiques fonctionnelles et structurales. Suite à des mutations chez la drosophile comme dCREB2, de knock-out chez le rongeur, ou suite à l’utilisation de divers inhibiteurs de CREB chez l’aplysie, la mémoire à long terme est sévèrement perturbée (pour revues, Bailey et al., 1996 ; Silva et
al., 1998 ; Bozon et al., 2003 ; Perazzona et al., 2004 ; Alberini, 2009). De plus,
l’activation de la transcription dépendante de CREB mettrait en jeu l’activation d’un CREB activateur et d’un CREB répresseur chez l’aplysie (Bailey et al., 1996 ; Alberini, 2009). Ce phénomène est encore soumis à discussion chez la drosophile (Perazzona et al., 2004).
Les autres voies moléculaires
Il existe d’autres molécules, moins étudiées, mais pourtant impliquées dans la formation de la mémoire à long terme : la calcineurine, une phosphatase calcium-dépendante permet de réguler l’activation de certaines kinases (Mansuy et al., 1998 ; Mansuy et Shenolikar, 2006), certaines molécules d’adhésion cellulaire nécessaires pour le remodelage de l’architecture synaptique (Cheng et al., 2001 ; Bisaz et al., 2009), ou encore divers types de récepteurs et canaux comme les récepteurs NMDA, les récepteurs nicotiniques, les canaux calciques ou les récepteurs aux amines biogènes (Castellano et
al., 1997 ; Rampon et al., 2000 ; Shimizu et al., 2000 ; Woodside et al., 2004 ; Bitner et al., 2007 ; Dacher et Gauthier, 2008 ; González-Burgos et Feria-Velasco, 2008 ; Seoane et al., 2009).
Ces « molécules de la mémoire », dont l’activation est liée directement ou indirectement au calcium, permettent l’expression de gènes cibles et donc la synthèse de nouvelles protéines, une étape majeure et primordiale dans la formation de la mémoire à long terme.
b. Synthèse protéique et mémoire à long terme
L’une des plus importantes découvertes réalisée au cours du siècle dernier, sur les bases cellulaires et moléculaires de la formation de la mémoire à long terme, concerne la nécessité d’une synthèse de nouvelles protéines (pour revue, Davis et Squire, 1984). En effet, contrairement à la mémoire à court terme, la formation de la mémoire à long terme peut être altérée par des inhibiteurs de synthèse protéique, aussi bien chez les vertébrés que les invertébrés (Stäubli et al., 1985 ; Tully et al., 1994 ; Freeman et al., 1995 ; Martin
et al., 1997 ; Meiri et Rosenblum, 1998 ; Menzel et al., 2001). Ce phénomène est donc
conservé au fil de l’évolution et pourrait être sous-tendu par des processus cellulaires et moléculaires communs.
La synthèse protéique requise pour la formation de la mémoire à long terme correspond non seulement à la traduction d’ARN messagers (ARNm) déjà présents au sein du réseau de neurones qui stocke l’information, mais aussi à la transcription de nouveaux gènes menant à la synthèse de nouvelles protéines.
La synthèse protéique correspondant à la traduction d’ARNm déjà présents dans le réseau de neurones, se situe préférentiellement au sein des dendrites et des épines dendritiques (supports anatomiques des connexions synaptiques). La majeure partie de cette synthèse protéique a lieu localement au niveau des dendrites, où se trouvent les composants majeurs de la machinerie traductionnelle, à savoir les ribosomes, les facteurs de traduction et bien sûr les ARNm (Steward et Schuman, 2001 ; Bramham et Wells, 2007). De plus, des travaux récents ont montré que cette synthèse protéique locale est nécessaire pour différentes formes de plasticité synaptique (Vickers et al., 2005), et pour la stabilisation de nouvelles synapses créées à la suite d’un apprentissage (Miniaci et al., 2008). Ainsi, cette synthèse protéique locale participerait aux modifications synaptiques nécessaires au stockage à court et à long terme d’une information.
La synthèse protéique impliquant la transcription de nouveaux gènes, est composée de deux vagues post-apprentissage, nécessaires pour la formation de la mémoire à long terme (Freeman et al., 1995 ; Artinian et al., 2007 ; Izquierdo et al., 2006 ; Laroche, 2006). Des expériences principalement réalisées chez les vertébrés suggèrent qu’une première vague se produit rapidement après l’apprentissage (jusqu’à 1 heure) et la seconde entre 3 et 6 heures après l’apprentissage (Grecksch et Matthies, 1980 ; Freeman
et al., 1995; Bourtchuladze et al., 1998; Quevedo et al., 1999; Igaz et al., 2002 ; Igaz et al., 2004 a, b ; Artinian et al., 2007). Ces vagues de synthèse protéique correspondent à la
transcription de gènes précoces ou « immediatly early genes » (IEGs) par l’activation de facteurs de transcription comme CREB, suivie de la transcription de gènes tardifs (figure
suivent les variations de la protéine CREB phosphorylée suite à un apprentissage (pour revues, Izquierdo et al., 2006 ; Alberini, 2009).
Cette représentation du décours temporel de la synthèse protéique qui suit l’apprentissage peut néanmoins varier en fonction des conditions d’apprentissage. En effet, chez le rongeur, un apprentissage « faible », lors d’un seul essai de conditionnement de peur, nécessiterait deux vagues de synthèse protéique alors qu’un apprentissage « fort », lors de trois essais de conditionnement de peur, ne nécessiterait que la première vague (Bourtchouladze et al., 1998).
Figure 4. Représentation schématique du décours temporel de l’expression des gènes après un apprentissage. La première phase de transcription correspond à celle des gènes précoces (ou IEGs) avec un
pic d’expression environ 30 minutes après l’apprentissage. Cette transcription est suivie de celle des gènes tardifs pendant une période beaucoup plus longue avec un pic d’expression entre 3 et 6 heures après l’apprentissage.
La première étape de transcription qui suit l’apprentissage est celle des IEGs
(Morgan et Curran, 1989 a, b ; Clayton, 2000 ; Guzowski, 2002 ; Miyashita, 2008 ; Alberini, 2009). Les IEGs codent pour des variétés différentes de protéines classées en deux catégories principales:
Il existe des IEGs effecteurs qui participent à une grande variété de fonctions cellulaires, telles que la croissance cellulaire (BDNF, Narp), la signalisation intracellulaire (RheB, RGS-2, Homer 1a), des modifications synaptiques au niveau structural (Arc, Homer 1a, Narp, TPA, BDNF), l’homéostasie synaptique (régulation du
nombre de synapses) (Arc, Homer 1a) et le métabolisme (COX-2) (Guzowski, 2002 ; Lanahan et Worley, 1998 ; Shepherd et al., 2006 ; Miyashita, 2007). Ces fonctions sont compatibles avec des modifications synaptiques sous-jacentes à une plasticité synaptique et à la formation de la mémoire (Dragunow, 1996). Par exemple, des inhibitions pharmacologiques des protéines ou des inhibitions de l’expression de gènes, comme Arc,
BDNF ou homer1a entraînent des déficits mnésiques (Bekinschtein et al., 2007 ;
Miyashita, 2007 ; Lu et al., 2008 ; Inoue et al., 2009).
La deuxième catégorie d’IEGs est celle des gènes régulateurs de transcription comme
c-fos, c-jun (aussi nommé, AP-1), zif268 (aussi nommé, Egr-1, Krox24) et C/EBP
(Greenberg et al., 1986 ; Morgan et Curran, 1989 a, b ; Clayton, 2000 ; Guzowski, 2002 ; Miyashita, 2007 ; Alberini, 2009). Ces gènes, comme leur nom l’indique, vont réguler la transcription de gènes cibles dit tardifs, par un recrutement de la machinerie transcriptionnelle, et sont clairement requis pour la formation de la mémoire à long terme (pour revues, Guzowski, 2002 ; Miyashita, 2007 ; Alberini, 2009).
La seconde étape de transcription qui a lieu 3 à 6 heures après un apprentissage,
correspond à celle des gènes cibles tardifs. Ces gènes sont impliqués dans divers mécanismes cellulaires permettant notamment les modifications synaptiques à long terme et donc la stabilisation de la trace mnésique (pour revues, Clayton, 2000 ; Miyashita, 2007 ; Laroche, 2006). Par exemple, durant cette période, des kinases, des récepteurs ou des enzymes sont exprimés dans l’hippocampe de rongeur après un conditionnement de peur (pour revue, Izquierdo et al., 2006). Depuis quelques années, des chercheurs tentent d’identifier et de comprendre le rôle des gènes impliqués dans la formation de la mémoire à long terme (Dubnau et al., 2003b ; Igaz et al., 2004 b ; Izquierdo et al., 2006). Ces études réalisées sur différents modèles animaux se révèlent très complexes et les résultats ne permettent pas d’établir de conclusions claires.
La synthèse de protéines détiendrait ainsi un rôle clé dans le renforcement des connexions synaptiques pré-existantes, ainsi que dans l’établissement de nouvelles connexions fonctionnelles afin de stocker l’information à long terme au sein d’un réseau spécifique de neurones. Ces deux principaux types de modifications synaptiques sont présentés en détail dans les paragraphes suivants.
c. Mémoire à long terme et modifications de l’efficacité synaptique : le phénomène de potentialisation à long terme
La formation de la mémoire repose sur des modifications de l’efficacité de la transmission synaptique (ou force synaptique) permettant la stabilisation de la trace mnésique à long terme. Le modèle cellulaire et moléculaire sur lequel reposeraient ces modifications synaptiques fonctionnelles, est la potentialisation à long terme (PLT) (pour
revue, Lynch 2004). C’est en 1973 que Bliss et Lømo ont démontré pour la première fois que la répétition de stimulations à haute fréquence sur des tranches d’hippocampe de lapin, entraînait une PLT dans les cellules granulaires du gyrus denté de l’hippocampe (Bliss et Lømo, 1973). Les caractéristiques principales de la PLT majoritairement étudiées in vitro (Bliss and Collingridge, 1993 ; Abraham et al., 1995), suggèrent que la PLT serait impliquée dans la formation de certaines formes de mémoire (pour revues, Martin et al., 2000 ; Kandel, 2001 ; Lynch, 2004 ; Hawkins et al., 2006 ; Reymann et Frey, 2007 ; Neves et al., 2008).
La PLT et la formation de la mémoire présentent les trois caractéristiques communes suivantes :
● La PLT a lieu dans certaines structures nécessaires à la formation de la mémoire comme l’hippocampe chez le mammifère (pour revue, Lynch, 2004).
● La PLT et la mémoire présentent des décours temporels similaires. De la même façon qu’il existe une mémoire à court terme, il existe une PLT précoce qui dure 2 à 3 heures. Ces deux processus sont indépendants de la synthèse protéique. Il existe également une PLT tardive qui perdure des heures in vitro et des semaines in vivo (Lynch, 2004). Cette PLT tardive, comme la mémoire à long terme, requièrent la transcription de nouveaux gènes et la synthèse de nouvelles protéines pour être maintenues (Krug et al., 1984 ; Nguyen et al., 1994 ; Mochida et al., 2001 ; Kandel, 2001 ; Lynch, 2004 ; Costa-Mattioli et al., 2009)
● La formation de la PLT tardive induite par l’activation des récepteurs NMDA, comme la formation de la mémoire à long terme, nécessite l’activation de voies de signalisation intracellulaire. Parmi ces voies de signalisation, on retrouve les voies impliquant la PKA, la CaMKII, les MAPKs et le facteur de transcription CREB (Morris, 1989 ; Lynch, 2004 ; Reymann et Frey, 2007).
Aujourd’hui, le lien entre la consolidation mnésique et l’induction de la PLT n’a pas encore été clairement démontré. Cependant, Whitlock et ses collaborateurs (2006) ont été les premiers à montrer très récemment que l’apprentissage induit une PLT in vivo dans l’hippocampe de rat.
Par ailleurs, il est important de noter que la PLT n’est pas le seul phénomène de plasticité synaptique probablement lié à la formation de la mémoire. En effet, la dépression à long terme (DLT), qui correspond à une diminution de l’efficacité de la transmission synaptique, peut avoir un rôle important dans la formation de la mémoire à long terme (Massey et Bashir, 2007). Cependant, peu de données sont disponibles sur ce phénomène.
Des modifications de l’efficacité synaptique qui se produisent très rapidement après l’apprentissage (comme le phénomène de PLT) ne suffiraient pas à maintenir et à stabiliser le réseau de neurones impliqués dans le stockage de l’information. Ainsi, des modifications au niveau de la structure même des synapses vont aussi avoir lieu afin de permettre, conjointement aux modifications synaptiques fonctionnelles, le stockage à long terme de l’information.
d. Mémoire à long terme et modifications de l’architecture synaptique
Le stockage à long terme des souvenirs s’accompagne également de modifications de l’architecture synaptique du réseau de neurones impliqués. Ces changements synaptiques structuraux peuvent être regroupés en deux principales catégories : des modifications de la structure de synapses pré-existantes et des variations du nombre de synapses. Ces modifications de l’architecture des synapses des réseaux neuronaux, support du stockage de l’information, sont dépendantes de la synthèse de nouvelles protéines (pour revues, Bailey et Kandel, 1993 ; Bailey, 1999).
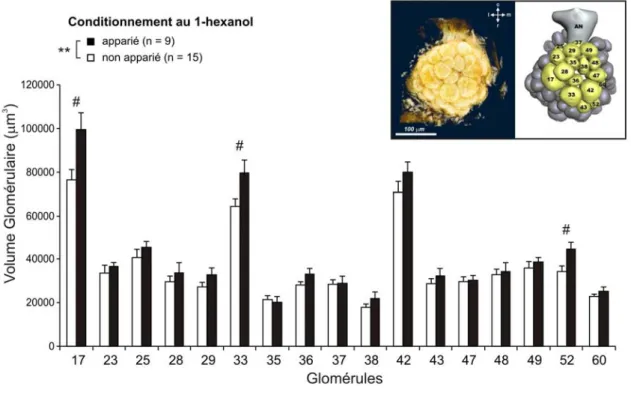
Des modifications synaptiques au niveau structural ont été observées suite à différents types d’apprentissage, pour différents modèles vertébrés ou invertébrés. En effet, suite à un apprentissage chez l’aplysie mais aussi chez le rongeur, des variations du nombre, de la taille et/ou du type de synapse ont été observées (Black et al., 1990 ; Bailey et Chen, 1991 ; Geinisman et al., 2000 a, b ; Kleim et al., 2002 ; Bailey et al., 2004) ainsi que des variations de la densité d’épines dendritiques, supports anatomiques des connexions synaptiques (Moser et al., 1994 ; Knafo et al., 2001). De la même façon, il a été relevé des variations de volume des structures nerveuses corrélées avec l’expérience, telles que l’hippocampe des conducteurs de taxis londoniens (Maguire et al., 2000), ou encore le lobe antennaire de l’abeille, premier centre de traitement de l’information olfactive, après la mémorisation spécifique à long terme d’une odeur (Hourcade et al., 2009). Les modifications structurales du réseau de neurones impliqués dans le stockage à long terme de l’information, peuvent être parfois dues à l’intégration de nouveaux neurones, comme c’est le cas dans l’hippocampe de souris adultes (Trouche et al., 2009).
En conclusion, ces travaux posent la question de l’existence d’un déclencheur de ces voies de signalisations, qui serait à l’origine de la formation de la mémoire à long terme. De nombreux efforts ont été investis pour identifier l’événement initial de la formation de la mémoire à long terme, mais aujourd’hui, ces recherches restent sans réponses claires. Ainsi, identifier ce déclencheur nous permettrait alors d’évaluer s’il joue un rôle important dans la régulation de l’expression de gènes requis pour de possibles
modifications structurales et fonctionnelles à long terme afin de stocker une information au sein d’un réseau de neurones.
Dans cette thèse, nous avons émis l’hypothèse que le calcium pourrait jouer un tel rôle. En effet, la majorité des « molécules de la mémoire » sont activées directement ou indirectement par le calcium. Ainsi, le chapitre qui suit fait état des travaux relatant de la nature de la régulation calcique intracellulaire ainsi que son rôle dans de nombreuses et diverses fonctions neuronales qui nous ont amenés à émettre notre hypothèse.
I.2.
Partie II. Le calcium, un acteur clé des fonctions
nerveuses
I.2.1.
Le calcium et les fonctions neuronales
Le calcium, qui peut sembler être un messager ubiquitaire, régule les processus vitaux de la cellule tout au long de la vie. La régulation des variations de la concentration intracellulaire de calcium ([Ca2+]i) est réalisée de façon spatio-temporelle. Ce signal calcique est médié par différents récepteurs impliquant diverses sources de calcium, extracellulaire mais également intracellulaire, au sein des organites contenant les stocks calciques. Pour assurer son homéostasie, le calcium interagit avec de nombreux éléments de la cellule (« calcium toolkit ») (Berridge, 1998, 2003).
Le signal calcique joue un rôle important dans la régulation de nombreux processus neuronaux. En effet, il est impliqué dans l’induction neurale (Moreau et Leclerc, 2004 ; Moreau et al. 2008) dans le développement neuronal , comme la migration cellulaire au niveau du système nerveux central (Komuro et Kumada, 2005), la croissance axonale (Gomez et Spitzer, 1999), le développement des dendrites (Zou et Cline, 1999 ; Redmond et Ghosh, 2005), la synaptogenèse (Lohmann et al., 2005), mais aussi l’apoptose (Szalai
et al., 1999) et la survie neuronale (Collins et al., 1991). Le calcium régule aussi la
libération de neurotransmetteurs et l’excitabilité de la membrane (Hille, 1978, 1992 ; Sudhof, 2004 ; Neher et Sakaba, 2008). Il est aussi fortement impliqué dans la régulation de l’expression de gènes (Greenberg et al., 1986 ; Alkon et al., 1998 ; Bito, 1998 ; Carrasco et al., 2004 ; Batut et al., 2005) et dans la plasticité synaptique telle que la PLT ou la DLT, modèles moléculaires sous-jacents à la formation de la mémoire (Bliss et Collingridge, 1993 ; O’Mara et al., 1995 ; Lisman, 2001 ; Lynch, 2004 ; Raymond et Redman, 2002, 2006). Enfin, un nombre important d’expériences comportementales a révélé, par des études indirectes, l’implication du calcium dans la formation de la mémoire (Ohnuki et Nomura, 1996 ; Quevedo et al., 1998 ; Balschun et al., 1999 ; Blackwell et Alkon, 1999 ; Futatsugi et al., 1999 ; Kouzu et al., 2000 ; Rodrigues et al., 2001 ; Salinska et al., 2001 ; Bauer et al., 2002 ; Woodside et al., 2004 ; Edwards et Rickard, 2006 ; Baker et al., 2008, 2009 ; Seoane et al., 2009).
Afin d’étudier et de mieux appréhender le rôle du calcium dans les fonctions nerveuses, et notamment dans la formation de la mémoire, il est indispensable de comprendre au niveau cellulaire, sa distribution spatiale, son homéostasie, et à un niveau plus intégré, son implication dans les processus de plasticité synaptique et dans les processus mnésiques. En effet, ces informations permettent de comprendre l’effet