T
T
H
H
È
È
S
S
E
E
En vue de l'obtention du
D
DO
OC
C
TO
T
OR
RA
AT
T
D
DE
E
L
L’
’U
UN
NI
IV
VE
E
RS
R
SI
IT
TÉ
É
D
DE
E
T
T
OU
O
UL
L
OU
O
US
SE
E
Délivré par l’Université Paul Sabatier – Toulouse III Discipline : Physiopathologie moléculaire, cellulaire et intégrée
JURY
Pr Bertrand PERRET Président Dr Julie DECHANET-MERVILLE Rapporteuse
Dr Francis HARAUX Rapporteur
Pr Monique CAPRON Examinatrice
Dr Eric CHAMPAGNE Examinateur
Dr Laurent MARTINEZ Invité
Dr Xavier COLLET Invité
Ecole doctorale : Biologie - Santé - Biotechnologie Unité de recherche : INSERM U563, Département LML, CPTP Directeurs de Thèse : Dr Eric CHAMPAGNE, Dr Laurent MARTINEZ
Présentée et soutenue par Pierre VANTOUROUT Le 30 Juin 2009
R
OLE DE L’E
CTO-F
1-ATP
ASE DANS LAR
ECONNAISSANCE DESC
ELLULES« As an adolescent I aspired to lasting fame, I craved factual certainty, and I thirsted for a meaningful vision of human life - so I became a scientist. This is like becoming an archbishop so you can meet girls. »
REMERCIEMENTS
Je tiens à remercier tout d’abord les membres de mon jury de thèse
Madame le Docteur Julie Déchanet-Merville pour m’avoir fait l’honneur d’être rapporteuse de ma thèse. Je vous remercie pour vos commentaires très gentils sur mon travail de thèse ainsi que pour la discussion lors de la soutenance qui était enrichissante. Je suis très content que vous ayiez pu venir malgré le contretemps et j’ai apprécié nos discussions.
Monsieur le Docteur Francis Haraux pour m’avoir fait l’honneur d’être également rapporteur de ce travail. Je vous suis très reconnaissant d’avoir accepté malgré le peu de points communs entre vos travaux et mon sujet de thèse. Je vous remercie également pour vos commentaires, pour la discussion très intéressante ainsi que pour votre aide sur les propriétés enzymatiques de l’ATP synthase, qui m’a été très précieuse lors de la rédaction de mon manuscrit.
Madame le Professeur Monique Capron pour m’avoir fait l’honneur d’assister à ma soutenance de thèse bien que l’université ne vous ait pas retenue comme rapporteuse. Je vous remercie d’avoir fait tout ce trajet avec un timing très serré, ainsi que pour votre gentillesse et la discussion intéressante que nous avons pu avoir au cours de et après la soutenance.
Monsieur le Professeur Bertrand Perret pour m’avoir fait l’honneur de présider mon jury de thèse et pour vos commentaires, questions et remarques extrêmement pertinents et très utiles. Je vous remercie également de m’avoir accueilli au sein du département LML ainsi que pour votre gentillesse, votre implication et l’intérêt que vous avez porté à mes travaux, et votre disponibilité.
Monsieur le Docteur Eric Champagne, mon directeur de thèse. Comme tu l’as dit nous avons passé de nombreuses années ensemble avec des hauts et des bas dans nos recherches mais je pense (et j’espère !) que dans l’ensemble nous avons fait du très bon travail. Je te remercie de m’avoir fait découvert les γδ et d’avoir entretenu mon intérêt pour l’immunologie bien que ce ne soit pas ma formation initiale. Je te suis également reconnaissant de m’avoir accordé ta confiance dans le développement de ces projets ainsi que de m’avoir laissé une grande autonomie qui me sera je pense très utile par la suite. Je te remercie également pour tes mots très gentils pendant la soutenance qui m’ont beaucoup touché. J’espère que tes nouveaux projets porteront tous leurs fruits et que les travaux que nous avons menés à bien ouvriront de nouvelles directions fructueuses.
Monsieur le Docteur Laurent Martinez, mon co-directeur de thèse. Merci d’avoir cru en moi, de m’avoir soutenu et de t’être grandement impliqué dans mes travaux, bien que très éloignés de tes perspectives de recherche. Je te remercie également pour ta disponibilité de tout instant et pour toutes les discussions que nous avons pu avoir, scientifiques ou non. Un grand merci également pour tes commentaires très gentils lors de la soutenance. Je te souhaite le meilleur pour ta nouvelle équipe et j’espère que la voie Ecto-F1-ATPase / P13Y2 (coquille volontaire !) continuera de fournir des résultats toujours aussi intéressants, tout comme tes nombreux nouveaux projets.
Monsieur le Docteur Xavier Collet pour avoir participé activement à mon jury. Je te remercie pour toutes tes questions (nombreuses comme toujours !) très intéressantes et pour ton implication dans ce sujet et le grand intérêt que tu lui as porté. Je te remercie également de m’avoir accueilli pendant toutes ces années au sein de ton équipe, pour ta disponibilité de tout
instant (malgré ta double casquette !) et pour ton soutien tout au long de ma thèse. Sans oublier ta bonne humeur et ton humour, bien sûr.
Je remercie tous les membres présents et passés du Thème A
Tous les chercheurs de l’équipe.
Je vous remercie tous pour votre gentillesse, votre disponibilité et pour votre soutien sur un projet bien éloigné de vos thématiques de recherche. Désolé de vous avoir assomé avec toute cette immunologie !
François pour ton humour, ta gentillesse, et désolé mais tu n’auras pas réussi à me convertir aux Macs ! Ha, et puis désolé pour la bio mol aussi, mais tu le sais, c’est ma grande passion ! Mêmes excuses pour Ronald, et je te souhaite le meilleur dans ta nouvelle direction et j’espère que l’aventure Cerenis sera pleine de succès.
Stéphane pour tout ton temps et ta patience pour tous ces concepts d’enzymologie qui me semblaient bien obscurs, ainsi que ta bonne humeur permanente.
Dernière (mais pas des moindres), merci Christine pour ta gentillesse, tous tes conseils (labo et hors labo) et pour ta disponibilité de tout instant. Je te suis infiniment reconnaissant de m’avoir énormément aidé pendant les périodes difficiles, je ne l’oublierai jamais. Je te souhaite de tout cœur le meilleur pour tes nouveaux projets de recherche et je suis sûr que tes petites goutelettes t’apporteront de nombreux résultats passionants avant que tu t’envoles vers de nouveaux horizons plus « verts » !
Tous les post-doctorants (et affiliés) du laboratoire.
Véro, un grand MERCI en Arial 56 gras souligné (oui je ne le fais pas pour de vrai sinon la mise en page va me faire hurler). On ne se sera pas cotoyés très longtemps mais ce fut un réel plaisir. Merci pour toutes ces soirées organisées avec brio et pour ton dévouement pour toujours être Sam ! Un grand merci également pour tes corrections très efficaces, et pour t’être intéressée à mon projet. Je te remercie pour ta gentillesse, ton soutien, et ton aide dans les coups durs. J’espère que tu auras très bientôt ton poste (satané gamma factor) car tu le mérites amplement. Je t’attends donc très bientôt à Londres, sans faute !
Gérald (saluuuuuuuut ! namasteeeee !) merci pour ton soutien et tous tes conseils. Je te souhaite le meilleur pour la suite et j’espère que tu finiras par obtenir un poste car tu le mérites vraiment et tu seras sans aucun doute un excellent chercheur « officiel » sur qui tout le monde peut compter. J’ai beaucoup apprécié toutes les discussions que nous avons pu avoir au cours de ces quelques mois.
Annelise (ma presque compatriote !) merci pour ton soutien, ton humour et ton extrême gentillesse. Encore toutes mes félicitations pour ton nouveau bonheur, et je te souhaite le meilleur dans ton futur parcours.
Safouane, un grand merci à toi (le seul hormis notre petite équipe à faire de l’immuno, je me sentais moins seul !) pour tous tes conseils très avisés et l’intérêt que tu as porté à mes travaux. Je te souhaite toute la réussite que tu mérites pour tes nouveaux projets (même s’il ne s’agit plus d’immuno, traître !).
Michela, notre italienne préférée ! Ton passage très rapide dans l’équipe restera je pense dans toutes les mémoires. Je n’oublierai certainement pas ton infinie gentillesse et ta bonne humeur permanente qui ont fait partie des petits rayons de soleil du labo. Il semble que tu t éclates dans ton nouveau poste qui est plus près de ce que tu attends et aimes. Je te souhaite le meilleur pour cette nouvelle direction.
Finally, I would like to thank Jayati who joined our team for 2 years. It was a real pleasure to meet you and work with you. Your everlasting good mood was very pleasant and helpful. Thank you for all your advices and your great help on this project. I wish you and Ananda the best for your new positions and hope you will be able to get back to India soon and find a position there. On second thought, maybe I should have written this in French since you are almost fluent now!
Je remercie également tous les techniciens de l’équipe.
Corinne pour ta gentillesse, pour ces bons moments de rigolade au cours des soirées (un peu de verveine peut-être ?). Je te remercie pour ton aide inestimable (notamment ton expertise de l’HPLC !) et ta disponibilité permanente. Et bien sûr pour avoir veillé sur nous en tant que (ex) pro de l’hygiène et sécurité ! (je suis sûr que cette douche nécessite un contrôle supplémentaire, il faudrait qu’on s’en occupe avant que je parte !). Je te souhaite le meilleur pour tes nouveaux projets.
Chris, pour ta gentillesse, tes conseils, et ton moral d’acier ! Merci d’avoir été présente et également d’avoir assuré avec perfection le bon fonctionnement de nos réserves ! A toi aussi je souhaite le meilleur pour tes nouveaux projets et je suis sûr que tu exporteras avec brio la hyène attitude !
Super Michel pour ta bonne humeur constante et toute ton aide, notamment à mon arrivée dans le labo. Merci aussi pour toutes ces discussions très agréables que nous avons pu avoir sur tous les sujets possibles et imaginables. Je te souhaite une très bonne fin de carrière et une belle et heureuse retraite amplement méritée !
Et enfin Guillaume, merci pour ton amitié et ton soutien, ainsi que pour cette mémorable finale de l’euro (dois-je rappeler que tu as fait une lamentable performance à l’euro-foot ? Très décevant de la part d’un tel expert en sports !). Je te félicite pour ta persévérance malgré la difficulté à trouver un poste, et j’espère que tu finiras par en obtenir un « pour de vrai » car tu le mérites très largements. Je te souhaite bonne route ainsi qu’à Emilie, et si tu comptes faire un saut à Londres à l’occasion c’est sans problème !
Je n’oublie pas les petites nouvelles du laboratoire.
Céline félicitations pour ton DEA et mes meilleurs vœux de réussite pour la poursuite de tes études (avec l’activité clinique en plus !). Je suis sûr que tout se passera bien et j’espère que tu pourras poursuivre la carrière de ton choix. Et que tu resteras dans la recherche, bien sûr ! Manuela (bouh !), un grand merci pour ta gentillesse, ton soutien, et ton aide à beaucoup de niveaux. Je te félicite également pour ton DEA. A l’heure où j’écris ces lignes tu ne sais pas encore si tu auras une bourse ministérielle mais je crois en toi et je suis sûr que tu donneras le meilleur comme tu l’as fait pendant ton DEA (oui, je sais, on ne dit plus DEA et ça fait quatre fois déjà que je l’écris, mais ceci est dû à mon grand âge bien sûr). Je suis sûr que tu feras une très bonne doctorante et je te souhaite le meilleur. Tu peux bien sûr compter sur moi si tu as besoin de quoi que ce soit, et puis je ne serai pas bien loin, tu viens quand tu veux !
Un petit chapitre spécial pour mes deux ex-thésardes préférées !
Bin oui, je n’allais pas vous placer ni dans les thésards, chères Docteurs, ni dans les post-docs du labo, et puis vous méritez largement votre section rien qu’à vous !
Flo, merci pour tous tes conseils et ton soutien surtout ces derniers mois, pendant la douloureuse phase d’accouchement du présent bébé. Je n’oublierai pas ton humour, ta bonne humeur et ton côté déjanté qui te va si bien. Je suis content que tu aies pu faire ce qui te plaisait à l’issue de la thèse et que tu aies trouvé un bon post-doc où tu t’éclates visiblement. Tout le meilleur pour la suite, et on se voit de toute façon très bientôt à Amsterdam !
Ma petite Camcam (ou devrais-je dire Mère Noël ?), un très grand merci pour toutes ces années passées ensemble au labo. On a suivi le même parcours (complètement atypique on pourrait dire) et ça nous a pas trop mal réussi ! Je n’oublierai jamais ton humour, ta gentillesse, ta bonne humeur permanente et tous tes conseils. Et tes fringues, bien évidemment ! Il y a eu du relâchement sur la fin d’ailleurs, probablement la faute à Lilian ! Tous mes vœux de réussite pour la poursuite de ta carrière, je suis content que tu aies pu trouver toi aussi un très bon post-doc. J’ai beaucoup admiré ta persevérance et tes facultés d’adaptation malgré un sujet pas évident et une situation parfois très délicate. Ton petit lutin te souhaite le meilleur, accompagnés de vœux de bonheur pour toi et le grand chef Lilian (rien à voir avec les indiens). On se revoit très bientôt (plusieurs fois !) et vous êtes tous les deux les bienvenus à Londres quand vous voulez, évidemment. TADAAAAAA !
Et pour finir, mes très chers compagnons, les inestimables, inimitables et irremplaçables thésards du thème A !
Aurélie, halala le dossier… la spécialiste ès-diplomatie ! Bon bon, je plaisante… un grand merci à toi pour ton soutien à beaucoup de niveaux, pour tous ces bons moments en soirée et pour nos nombreuses discussions. Je suis vraiment content de t’avoir rencontrée et d’avoir travaillé avec toi (très partiellement, mais nous avons deux articles en commun quand même, la classe !). Hormis ta diplomatie (oui oui, j’insiste) je retiendrai de toi ta gentillesse et ta timidité qui a très vite disparu (la faute à Véro ? Je suis sûr que les dates coïncident) pour laisser place à « la petite » que nous aimons tous. Je te souhaite une très bonne fin de thèse ete si tu as besoin de quoique ce soit tu n’hésites pas, c’est la tradition (j’ai suffisamment embêté Camille pour le savoir !). Pas de faux espoirs, je participerai activement à ton anti-thèse, et promis je serai là pour la soutenance ! Je suis sûr que tout se passera très bien pour toi et j’espère que tu trouveras très vite un post-doc qui permettra à Fred de t’accompagner. Tous mes vœux de bonheur à tous les deux (c’est pour quand ? OK OK j’ai rien dit !). Et quand vous voulez vous passez me voir à Londres bien sûr !
Pequeñita Claudia ! Oui hum, bon, je vais arrêter là l’espagnol, j’ai vraiment besoin de m’y remettre. Un grand merci pour ta gentillesse, ton humour (et tes magnifiques chansons !) et pour ton soutien et ton aide sur tous les plans. Je te souhaite à toi aussi une très bonne fin de thèse qui approche (t’en es oùùùùùù ? T’as fini le chapitre endothélium ???? Oui moi aussi je fais ma Véro, y’a pas de raisons !) et bien sûr je ferai le maximum pour assister à ton triomphe ! Et puis un très bon retour au pays et je vous souhaite à toi et Pablo le meilleur pour la suite. Vous êtes tous les deux formidables et je suis très heureux d’avoir pu vous cotoyer ! Et si jamais vous revenez en cette bonne vieille Europe j’espère vous voir à Londres tous les deux !
Emmanuel (cher Docteur !) merci pour ta gentillesse, ton humour et ton soutien. Je te souhaite également une bonne fin de thèse et j’espère que tes travaux se concrètiseront bientôt. Bonne chance pour la suite de ta carrière et j’espère que tu resteras dans la recherche car tu sembles vraiment t’y épanouir et y être brillant.
Zeina, tu démarres tout juste ta thèse donc bon courage et accroche toi. Tout le meilleur pour la suite.
Je profite enfin de cette partie dédiée au thème A pour remercier nos « membres invités » de l’ICR/Chimie/Fac/etc (bon désolé mais c’est compliqué votre histoire !).
Denis (ouuuuuyaaaaah. Oui bon, je ne sais pas comment retranscrire notre salut officiel par écrit). J’ai vraiment beaucoup apprécié de te cotoyer (à mi-temps ? enfin partiellement, enfin bref) pendant ces quelques mois. Merci pour ta sympathie et ton soutien. Bon courage pour la rédaction de ta thèse et je te souhaite le meilleur pour la suite.
Greg, très heureux également de t’avoir connu. On a bien rigolé et passé de bonnes soirées ensemble. J’espère vraiment que les galères s’arrêteront vite et que tu trouveras un poste le plus rapidement possible. Et n’oublie pas le logo de l’association internationale des toilettes, ça peut toujours servir (ainsi que l’albatros. Ou la mouette, je ne sais plus).
Edern, rencontre très brève et sporadique mais j’ai apprécié de discuter et rigoler avec toi. Bon courage pour la suite et j’espère que tu pourras trouver rapidement un job après ton M2.
Je remercie également tous les membres du thème B
Et plus particulièrement :
Fabienne (même si tu n’es pas venue à ma soirée de thèse !). Un immense merci pour ton soutien, ta disponibilité et ton aide inestimable dans les périodes difficiles. Je te remercie également pour ta bonne humeur, ton humour, et toutes nos discussions autour d’un café ou d’une cigarette. J’ai décidé comme tu le sais de laisser tomber les macaques pour mon post-doc (oui, tu n’allais pas y couper désolé !), la souris c’est bien plus porteur… Je te souhaite du fond du cœur le meilleur pour la suite et j’espère vraiment que nous resterons en contact. Muriel, merci pour ta bonne humeur, ton dynamisme et tous ces bons moments à la table café. Je te remercie également de t’être intéressée à mes travaux, et pour ton aide irremplaçable pour les dossiers de soutenance (c’est combien de rangs A extérieurs déjà ? Et puis c’est quoi cette histoire de prix de thèse ?!). Tout le meilleur pour la suite, pour cette nouvelle équipe avec Laurent !
Armelle, un grand merci à toi pour tous tes conseils et ton soutien. J’espère que tes projets se développeront et que tu seras bientôt à la tête d’une armée de thésards qui sauteront partout en salle de culture lorsqu’ils écriront leur thèse !
Enfin je ne pense pas que cela arrivera avec Audrey… si ? Merci à toi Dreydrey (un p’tit cookiiiiiiiie ?) pour ta gentillesse, ton écoute et ton soutien. Je te souhaite bon courage pour la poursuite de ta thèse qui sera brillante je suis sûr. Tous mes vœux à toi et Thibault qui est également quelqu’un de bien que j’ai eu plaisir à rencontrer bien que brièvement.
Anne, merci à toi également pour ta gentillesse, ton humour, et les nombreuses pauses clope passées à discuter. Et un grand merci d’avoir récupéré mon PV tu as assuré ! Bon courage pour la fin de ta thèse à toi aussi.
Clo, merci pour ta gentillesse, ta bonne humeur et ces concours endiablés de mots croisés. Merci également pour m’avoir dépanné si souvent pour de nombreux réactifs, qu’aurais-je fait sans toi ? Tu viens de quitter le département à l’heure où j’écris ces lignes donc je te souhaite le meilleur pour ta nouvelle direction.
Marianne, un grand merci pour ton humour et ta bonne humeur permanente bénéfique et très communicative. Bon courage pour la suite et j’espère que tu trouveras un vrai poste très prochainement.
Hélène, tu es la prochaine sur la liste ! Malheureusement je ne pourrai pas assister à ta thèse, mais je suis content que tu te sois finalement sortie des péripéties de date et autres formalités ! Je suis sûr que tout se passera bien et je te souhaite le meilleur pour ton post-doc. Peut-être en Angleterre, qui sait ?
Jean-Phiphi, merci à toi pour tous tes conseils et nos discussions. Et toutes mes félicitations pour ton poste, ct toutes mes félicitations pour ton poste, c’est génial !.
Pour finir tous mes remerciements également à Patrick, Nicole, Monique, Elvire, Emmanuelle et Françoise que j’ai beaucoup moins cotoyés mais que je n’oublie pas !
Pour finir un grand merci aux autres habitants du bâtiment C (les dom-tom)
Justine et son équipe de choc du plateau de lipidomique, Véronique et Séverine. Je n’ai pas eu à utiliser vos talents mais j’ai apprécié de vous cotoyer au cours de ces années. J’espère que le plateau se développera et je vous souhaite le meilleur pour la suite.
Tous les membres de l’équipe de Jean-Luc Davignon, Michel, Myriam et Jean-Frédéric. Ce fut un plaisir de vous rencontrer et de partager les locaux avec vous.
Et pour finir notre très chère Yvette qui prend tant soin de nous et sans qui nous serions perdus ! Je te remercie pour ta gentillesse, ta disponibilité et toute ton aide dans les démarches administratives. A bientôt à Londres alors ?
Je tiens à remercier tous les membres de l’IFR et du CPTP que j’ai rencontrés au cours de ces 6 années
Merci à tous pour votre accueil et votre aide.
Je pense notamment aux meilleures responsables de plateau de cytométrie (ma deuxième maison) et de d’imagerie de France (voire du monde !) Fatima (ainsi que Valérie) et Sophie. Merci pour votre aide inestimable dans la réalisation de ces travaux, pour votre gentillesse et pour toutes les discussions que nous avons pu avoir. Un grand merci également d’avoir participé à ma soirée de thèse, j’ai été très touché par votre présence.
Je tiens également à remercier les autres responsables des plateaux qui ont participé directement et indirectement à mes travaux. Hélène et Claudie, Jacques et Nordine (les McGuyver de l’U563 !) ainsi que Joël Teyssier, notre invité spécial de la pause déjeuner ! J’ai également une pensée pour tous les gens que j’ai cotoyés depuis la fac ou mon arrivée à l’U563. Damien, Cécile, Marinela, Hicham, Nico, Gaëtan, Frédérique Gaits, Hélène
Tronchère, Loïc Dupré et son équipe (notamment Ronan et Fanny), Bernard Payrastre et bien d’autres personnes que je n’oublierai pas.
Un grand merci spécial à Anne Huchenc-Champagne ! Merci infiniment d’avoir été aussi disponible le jour de ma thèse tu mérites vraiment une médaille (ou une casquette, c’est vrai). Merci pour ta gentillesse et ton soutien.
Je remercie finalement les enseignants de l’UAG et de l’UPS qui ont grandement contribué à mon désir de m’orienter vers la recherche.
Voilà donc pour la partie travail ! Encore merci à tous et tous mes vœux pour vos futurs travaux. Et de tout cœur, bon courage aux futurs impétrants. Si j’ai réussi à écrire ma thèse (moi et mon horreur de l’écriture…), vous pouvez tous le faire !
Je remercie également tous mes amis.
Djébraïl, trop longtemps perdu de vue mais récemment retrouvé, merci pour ton soutien et ton amitié indéfectible. Tous mes autres amis de collège et lycée, Florent, Julien, Frédo, Nicolas et tous les autres que je n’oublie pas.
Mes amis de l’université (UAG et UPS) et notamment Loïc, Charles, Arthur, Aline, Hayat, Anne-Claire, Greg et tous les autres. Nous avons tous pris des chemins différents mais je suis heureux d’avoir croisé votre route et je vous souhaite tout le bonheur possible.
Une mention spéciale à Donia pour être venue assister à ma thèse, ainsi qu’à toi et Magda pour être venues à ma soirée. Merci pour votre amitié, votre gentillesse et votre soutien. Vous venez quand vous voulez à Londres !
Mes amis de l’auberge, Jean-Pierre, Bachir (mon bro !), François, Matthieu et Mathieu (Dupontt et Dupont), Dom le tenancier, Pascal, Reda, Arnaud et tous les autres !
Egalement mes nouveaux amis de phdcomics (dont je conseille la lecture à tout le monde !), Catherine, Emily, Caoimhe, Fiona, Erwin, Michael, Joel, John et les autres. You all have my deepest thanks for all your advices, help and support. I wish you the best for your PhD, post-docs or jobs. I really look forward to meeting some of you soon and I hope I will be able to meet all of you guys one day.
Je terminerai ces remerciements par ma famille.
Un grand merci à mon papa de m’avoir aidé et accompagné pour mes études. Le résultat final n’est pas celui originellement prévu, mais le titre est le même après tout. Merci à toi pour ton aide et ton amour et je sais que je n’en serais pas là aujourd’hui sans toi.
A mes deux sœurs adorées, merci pour votre soutien et votre amour. Vous me manquez beaucoup et j’espère que vous me rendrez visite à Londres.
S
Sommaire
ABREVIATIONS ET ACRONYMES p.5
RESUME p.10
AVANT-PROPOS p.11
INTRODUCTION BIBLIOGRAPHIQUE p.17
Chapitre I : Le système immunitaire p.17
1) Généralités p.18
1.1) Organes et cellules p.18
1.2) Molécules fondamentales du système immunitaire : quelques exemples p.21
1.3) Déclenchement de la réponse immunitaire p.23
2) Les lymphocytes T p.25
2.1) Développement p.27
2.2) Les sous-populations T conventionnelles p.30
3) Les lymphocytes T non-conventionnels p.33
3.1) Les lymphocytes NKT p.34
3.2) Les lymphocytes MAIT p.37
Chapitre II : Les lymphocytes T γδ p.41
1) Généralités p.42
2) Développement : la priorité aux lymphocytes T γδ p.44
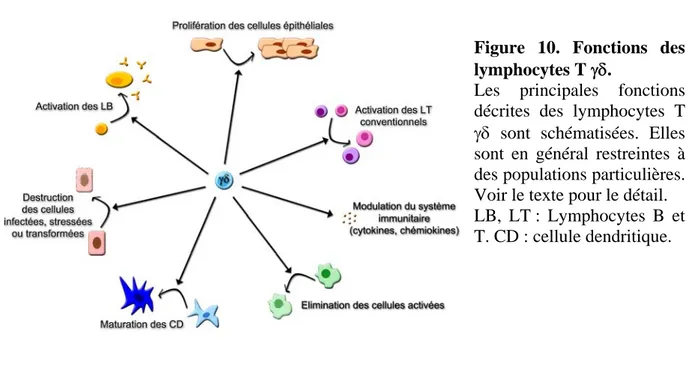
3) Fonctions des lymphocytes T γδ : multiplicité et diversité p.48
3.1) Fonctions effectrices p.49
3.2) Fonctions régulatrices p.53
4) Lymphocytes T γδ et pathologies : dualité de leurs rôles p.55
4.1) Pathogènes intracellulaires p.55
4.1.1) Bactéries et parasites p.55
4.1.2) Virus p.56
4.2) Pathologies inflammatoires et auto-immunes p.58
4.3) Cancer p.60
5) Antigènes reconnus par les TCR γδ : une diversité déroutante p.62
5.1) Molécules apparentées au CMH p.63
Sommaire
5.2.1) Phosphoantigènes p.67
5.2.2) Aminobisphosphonates et alkylamines p.70
5.3) L’Ecto-F1-ATPase p.72
Chapitre III : L’ATP synthase p.75
1) La F1FoATP synthase mitochondriale p.76
1.1) Généralités p.76
1.2) Structure p.77
1.3) Mécanisme enzymatique p.78
1.4) Pathologies liées à l’ATP synthase p.80
2) L’Ecto-F1-ATPase p.80
2.1) Ecto-F1-ATPase, la même enzyme que l’ATP synthase mitochondriale ? p.81
2.2) Une localisation inattendue et toujours inexpliquée p.84
2.3) Nouvelle localisation, nouveaux rôles p.87
2.3.1) Métabolisme du HDL-cholestérol p.88 2.3.2) Survie et prolifération des cellules endothéliales p.90 2.4) Synthétiser ou ne pas synthétiser, telle est la question p.93
3) Une fascinante enzyme qui n’a pas encore révélé tous ses secrets p.96
RESULTATS EXPERIMENTAUX p.99
Partie I : Interaction entre l’Ecto-F1-ATPase et le CMH-I p.99
1) Introduction p.100
2) Article 1 p.101
3) L’interaction est-elle spécifique d’un ou plusieurs isotypes de CMH-I ? p.109
3.1) Introduction p.109
3.2) Matériel et méthodes p.110
3.3) Résultats p.114
3.4) Discussion p.117
Partie II : L’ApppI, un prototype de phosphoantigène nucléotidique p.121
1) Introduction p.122
2) Article 2 (Soumis) p.124
Partie III : Présentation des phosphoantigènes par l’Ecto-F1-ATPase p.141
1) Introduction p.142
Sommaire
3) Modulation de l’activité enzymatique de l’ATP synthase par l’ApppI p.150
3.1) Introduction p.150
3.2) Matériel et méthodes p.151
3.3) Résultats et discussion p.152
Partie IV : Etude du métabolisme nucléotidique à la surface des cellules p.157
1) Introduction p.158
2) Article 4 p.159
3) Rôle de l’ANT dans la conversion ADP Æ ATP à la surface des cellules p.168
3.1) Introduction p.168
3.2) Matériel et méthodes p.169
3.3) Résultats et discussion p.170
DISCUSSION GENERALE ET PERSPECTIVES p.173
1) Interaction entre CMH-I et Ecto-F1-ATPase p.174
1.1) HLA-B et autres partenaires potentiels p.174
1.2) Implications fonctionnelles et physiopathologiques p.175
1.3) Rôle du CMH-I dans l’expression de l’Ecto-F1-ATPase ? p.177
2) Mécanisme de présentation des phosphoantigènes p.178
2.1) Voie(s) de transit de l’Ecto-F1-ATPase de la mitochondrie à la surface p.178
2.2) Protéines impliquées dans le routage/chargement de l’Ecto-F1-ATPase ? p.179
3) Métabolisme des phosphoantigènes p.180
3.1) Mécanisme d’internalisation de l’ApppI p.180
3.2) Nucleotide pyrophosphatase p.183
3.3) Autres phosphoantigènes et dérivés nucléotidiques p.184
4) Interactions entre phosphoantigènes, Ecto-F1-ATPase et TCR Vγ9/Vδ2 p.186
4.1) Liaison de l’ApppI sur l’Ecto-F1-ATPase p.186
4.2) Reconnaissance de l’Ecto-F1-ATPase par le TCR Vγ9/Vδ2 p.187
5) Spécificité d’espèce de la présentation des phosphoantigènes p.189
BIBLIOGRAPHIE p.191
ANNEXES (Article 5) p.207
Sommaire
A
Abréviations et acronymes
AA : AlkylAmines AAC : ADP, ATP Carrier AB : Acide Bongkrékique
ABCC5 : ATP-Binding Cassette sub-family C member 5 ADCC : Antibody-Dependent Cell Cytotoxicity
ADN : Acide DésoxyriboNucléique ADNc : ADN complémentaire AK : Adenylate Kinase
AMP, ADP, ATP : Adénosine mono-, di- ou triphosphate ANT : Adenosine Nucleotide Translocase
apo : apolipoprotéine
ApppI : gamma-P-(3-méthyl-but-3-en-1-yl)-5'-adénosine disodium triphosphate ARN : Acide RiboNucléique
ARNi: ARN interférent ARNm : ARN messager ARNt : ARN de transfert β2m : beta-2-microglobuline
BCR : B Cell Receptor (récepteur B à l’antigène) CCR : CC chemokine Receptor
CD : Cellule Dendritique (ne pas confondre avec CD suivi d’un numéro qui correspond à la nomenclature Cluster de Différenciation utilisée pour les marqueurs de surface)
CDR : Complementarity Determining Region
CMH : Complexe Majeur d’Histocompatibilité (de classe I ou II) CMV : Cytomégalovirus
CPA : Cellule Présentatrice d’Antigènes CSF : Colony Stimulating Factor
CXCR : CXC chemokine Receptor DETC : Dendritic Epidermal T Cell DMAPP : DiMéthyl-Allyl PyroPhosphate DMEM : Dulbecco’s Modified Eagle’s Medium DO : Densité Optique
DOXP : DeOxy-Xylulose-Phosphate DN : Double Négatif
Abréviations et acronymes
EAE : Experimental Autoimmune Encephalomyelitis EBV : Epstein Barr Virus
EEA1 : Early Endosomal Antigen 1
ELISA : Enzyme Linked ImmunoSorbent Assay
EMAP II : Endothelial Monocyte-Activating Polypeptide II ETP : Early T cell Progenitor
EtPP : Ethyl PyroPhosphate FPP : Farnesyl PyroPhosphate GFP : Green Fluorescent Protein HDL : High Density Lipoprotein
HDMAPP : Hydroxy- DiMéthyl-Allyl PyroPhosphate (aussi appelé HMBPP) HLA : Human Leukocyte Antigen
HMBPP : Hydroxy-Méthyl-Butényl PyroPhosphate HPLC : High Pressure Liquid Chromatography Hsp : Heat shock protein
HSV : Herpes Simplex Virus
HUVEC : Human Umbilical Vein Endothelial Cells ICAM-1 : Inter-Cellular Adhesion Molecule-1 ICOS : Inducible CO-Stimulator
IFN : Interféron Ig : Immunoglobuline
IHH : Immortalized Human Hepatocytes IL : Interleukine
IMGT : Système d’information international en ImMunoGénéTique IPP : Isopentenyl PyroPhosphate
ITAM/ITIM : Immunoreceptor Tyrosine-based Activation/Inhibition Motif KGF : Keratinocyte Growth Factor
KIR : Killer Inhibitory Receptor LAK : Lymphokine Activated Killer
LAMP-1 : Lysosomal Associated Membrane Protein-1 LB : Lymphocyte B
LDH : Lactate DesHydrogenase LDL : Low Density Lipoprotein LIR : Leucocyte Ig-like Receptor
Abréviations et acronymes
LT : Lymphocyte T
MAIT : Mucosal Associated Invariant T
MAPL : Mitochondria Anchored Protein Ligase
MICA/B : MHC class I polypeptide-related sequence A/B MttP: Microsomal triglyceride transfer Protein
MR1 : MHC-I-Related protein 1
MRP5 : Multidrug Resistance Protein 5 MSC : Mesenchymal Stem Cell
NADH : NicotinAmide Dinucléotide (forme réduite)
NARP : Neurogenic muscle weakness, Ataxia, Retinitis Pigmentosa N-BP : Aminobisphosphonates
NCL : Neuronal Ceroid Lipofuscinosis NDPK : Nucleoside DiPhosphoKinase NK : Natural Killer
NKT : Natural Killer T
NKR : Natural Killer Receptor (aNKR : activateur, iNKR : inhibiteur) NPP : Nucleotide PyroPhosphatase
OSCP : Olygomycin Sensitivity-Confering Protein PBMC : Peripheral Blood Mononuclear Cells PCR : Polymerase Chain Reaction
PEP : PhosphoEnol Pyruvate PHA : PhytoHémAgglutinine Pi : Phosphate inorganique PK : Pyruvate Kinase
PPADS : Pyridoxal-Phosphate-6-Azophenyl-2',4'-DiSulfonate PPD : Purified Protein Derivative
Ppt1 : Palmitoyl protein transferase 1
RAET : Retinoic Acid Early inducible Transcript RAG1/2 : Recombination Activating Gene 1/2 RE : Réticulum Endoplasmique
RFP : Red Fluorescent Protein
ROCK-I : Rho-associated Coiled-coiled Kinase I ROS : Reactive Oxygen Species
Abréviations et acronymes
RPS : Résonance Plasmonique de Surface
RT-PCR : Reverse Transcription-Polymerase Chain Reaction SEA : Staphylococcal Enterotoxin A
SH : Sérum humain
SIDA : Syndrôme de l’ImmunoDéficience Acquise SVF : Sérum de Veau Fœtal
SP : Simple Positif
TCR : T Cell Receptor (récepteur T à l’antigène) TdT : Terminal deoxyribonucleotidyl Transferase TGF : Transforming Growth Factor
TIL : Tumor Infiltrating Lymphocytes TLR : Toll-Like Receptor
TNF : Tumor Necrosis Factor
Tom : Translocase of outer membrane TT : Toxine Tétanique
TUBAg : TUBerculosis Antigen ULBP : UL16-Binding Protein
RESUME
La biologie des lymphocytes T (LT) Vγ9/Vδ2 est encore très peu comprise. Cette sous-population non-conventionnelle, spécifique à l’homme et certains primates supérieurs, joue un rôle important dans la défense de l’organisme contre des pathogènes intracellulaires (tels que mycobactéries et plasmodium). La reconnaissance des cellules infectées implique de petites molécules, appelées phosphoantigènes, formées d’un radical alkyl et un pyrophosphate. Les plus communs sont l’isopentenyl pyrophosphate (IPP, intermédiaire de la voie du mévalonate) et l’hydroxy-methyl-butenyl pyrophosphate (HMBPP, intermédiaire de la voie DOXP, voie alternative de synthèse de l’IPP chez certains organismes). Leur reconnaissance est mal caractérisée mais requiert un contact cellulaire, suggérant qu’une structure de présentation est nécessaire. Les LT Vγ9/Vδ2 sont également capables de lyser des lignées tumorales in vitro. L’implication de l’IPP a été établie et l’équipe a récemment identifié l’Ecto-F1-ATPase, une forme de l’ATP synthase mitochondriale exprimée à la surface, comme antigène participant à la reconnaissance des cellules tumorales. Cette thèse a eu pour but de mieux caractériser le rôle de l’Ecto-F1-ATPase dans cette reconnaissance.
Nous avons montré que l’Ecto-F1-ATPase interagit en surface des cellules avec les molécules du Complexe Majeur d’Histocompatibilité de classe I. HLA-B est préférentiellement impliqué dans cette interaction et diminue fortement l’activation les LT Vγ9/Vδ2. Cette interaction joue donc un rôle important dans le contrôle de la réactivité de ces lymphocytes et renforce le lien entre Ecto-F1-ATPase et immunité.
Dans une deuxième partie, nous avons étudié les mécanismes de reconnaissance de l’ApppI, un dérivé nucléotidique de l’IPP. Contrairement aux phosphoantigènes classiques, cette molécule est résistante aux phosphatases terminales, son mécanisme de reconnaissance est indirect et implique une activité de type Nucleotide pyrophosphatase, et elle peut être capturée par des cellules et les rendre sensibles aux LT Vγ9/Vδ2. Pour finir, l’ApppI nous a permis de démontrer que l’Ecto-F1-ATPase est impliquée dans la reconnaissance des phosphoantigènes.
Nos travaux représentent une avancée importante dans la compréhension des mécanismes de reconnaissance des cellules tumorales par les LT Vγ9/Vδ2. L’Ecto-F1-ATPase représente la première molécule directement impliquée dans la présentation des phosphoantigènes. Ces résultats pourraient à terme permettre le développement de stratégies d’immunothérapie anticancéreuse.
A
Avant-propos
Le système immunitaire est composé d’un très grand nombre de populations cellulaires différentes coopérant pour assurer la protection de l’organisme contre les pathogènes. Les lymphocytes T jouent un rôle central dans cette protection, en participant directement à l’élimination des pathogènes et des cellules infectées ainsi qu’à la mise en place de la réponse immunitaire. Ils sont en effet capables de contrôler la différenciation et l’activation des autres populations formant ce système extrêmement complexe. Le compartiment T se compose lui-même d’une grande variété de cellules ayant des phénotypes et des fonctions bien distincts. Parmi celles-ci, on distingue les lymphocytes T αβ, dits conventionnels, et les γδ, qui font partie des lymphocytes T non-conventionnels avec des sous-populations αβ particulières, les cellules NKT et MAIT. Découverts au milieu des années 80, ces lymphocytes restent extrêmement mal connus à l’heure actuelle. Ceci est notamment dû aux nombreuses différences entre les sous-populations γδ humaines et murines, ce qui rend leur étude délicate. Ils font partie de l’immunité dite innée, et à ce titre font partie des premières lignes de défense contre les pathogènes. Ils participent également à l’homéostasie tissulaire et jouent probablement un rôle important dans l’immuno-surveillance de la transformation tumorale.
Chez l’homme et certains primates « supérieurs », la grande majorité des lymphocytes T γδ circulants expriment un TCR composé d’une chaîne Vγ9 appariée à une chaîne Vδ2. Ils ont initialement été décrits pour participer à la réponse dirigée contre Mycobacterium
tuberculosis, l’agent responsable de la tuberculose. Ils sont activés par des antigènes
particuliers appelés phosphoantigènes, qui sont des molécules de très faible poids moléculaire (<1kDa) et dont la structure générale comprend une courte chaîne alkyl et un pyrophosphate. Ces molécules ont été identifiées il y a 15 ans, mais le mécanisme exact permettant leur reconnaissance par les lymphocytes T Vγ9/Vδ2 n’a jamais été identifié. Cette reconnaissance fait probablement intervenir une structure de présentation antigénique. Dans le cas de toutes
Avant-propos
les autres sous-populations T, ce rôle est rempli par les molécules du Complexe Majeur d’Histocompatibilité (CMH), classiques ou apparentées. Néanmoins, en ce qui concerne les lymphocytes T γδ, il est communément admis qu’elles ne sont pas impliquées.
Ces lymphocytes sont également capables de lyser de nombreuses lignées tumorales in vitro. De nombreuses études ont cherché à identifier l’antigène reconnu et plusieurs candidats ont été proposés, bien qu’aucun travail n’ait pu mettre en évidence de reconnaissance directe de ces antigènes. Récemment, les phosphoantigènes ont également été impliqués dans cette reconnaissance.
Mon équipe d’accueil est multidisciplinaire, avec trois thématiques principales. Deux d’entres elles sont centrées sur le métabolisme du cholestérol : les mécanismes et récepteurs impliqués dans son absorption intestinale (thématique dirigée par Xavier Collet), et la captation hépatique des lipoprotéines de haute densité (HDL). Récemment, les travaux de Ronald Barbaras et Laurent Martinez ont permis l’identification de l’Ecto-F1-ATPase, une forme de l’ATP synthase mitochondriale exprimée à la surface des cellules, comme récepteur majeur le l’apolipoprotéine A-I, protéine majoritaire des HDL, à la surface des hépatocytes. En collaboration avec ce groupe, la dernière thématique de l’équipe, menée par Eric Champagne, mon directeur de thèse, a permis d’identifier l’Ecto-F1-ATPase comme antigène participant à l’activation des lymphocytes T Vγ9/Vδ2 par les cellules tumorales. Les données obtenues montrent que ce complexe enzymatique est capable d’activer spécifiquement ces cellules et qu’il lie directement leur récepteur antigénique. Cette thèse s’inscrit dans la poursuite de ces travaux.
L’Introduction Bibliographique sera composée de trois grands chapitres. La première partie sera dédiée à une vue globale du système immunitaire et particulièrement centrée sur
Avant-propos
les lymphocytes T, afin de fournir des éléments utiles pour la compréhension de la biologie de la population Vγ9/Vδ2. Elle s’achèvera par une description plus détaillée des sous-populations αβ non-conventionnelles qui partagent quelques caractéristiques communes avec les lymphocytes T γδ.
Le deuxième chapitre proposera une revue des connaissances actuelles sur les lymphocytes T γδ. Tous les aspects de leur physiologie, de leur développement aux antigènes qu’ils reconnaissent, seront abordés.
Enfin, l’objet de cette thèse ayant été de mieux comprendre le rôle de l’Ecto-F1-ATPase dans la reconnaissance des cellules tumorales par les lymphocytes T Vγ9/Vδ2, la dernière partie de l’introduction sera dédiée à une brève description de l’ATP synthase mitochondriale. Elle sera également destinée à présenter un résumé des données récentes sur l’Ecto-F1-ATPase, qui participe à de nombreuses fonctions cellulaires en plus de son rôle dans l’immunité anti-tumorale.
La partie Résultats Expérimentaux présentera les données obtenues au cours de cette thèse. Nous avons tout d’abord cherché à mieux caractériser l’expression de l’Ecto-F1-ATPase à la surface d’un nombre important de lignées cellulaires et nous avons montré qu’elle peut interagir avec les molécules de la famille du CMH-I. Cette interaction entraîne un masquage d’épitopes de l’Ecto-F1-ATPase, ce qui indique que l’expression de ce complexe à la surface des cellules peut ne pas être détectée par des méthodes directes. HLA-B, un des isotypes classiques de CMH de classe I, semble préférentiellement impliqué et responsable d’une forte inhibition de la stimulation des lymphocytes T Vγ9/Vδ2.
Nous avons ensuite étudié les propriétés antigéniques de l’ApppI, un analogue d’ATP nouvellement décrit et dont la structure est similaire à celle de dérivés nucléotidiques de phosphoantigènes identifiés dans des extraits mycobactériens. Nos résultats indiquent que
Avant-propos
l’ApppI n’est pas un antigène au sens propre du terme puisqu’il n’active pas directement les lymphocytes T Vγ9/Vδ2. En revanche, il possède des propriétés uniques qui le différencient des phosphoantigènes classiques et suggèrent que les dérivés nucléotidiques pourraient jouer un rôle important dans le mécanisme de présentation des phosphoantigènes.
L’ApppI nous a également permis de réconcilier les données impliquant les phosphoantigènes et l’Ecto-F1-ATPase dans la reconnaissance des cellules tumorales. En effet, nous avons pu montrer qu’elle est capable de présenter des phosphoantigènes aux lymphocytes T Vγ9/Vδ2. Ces données sont également en faveur d’une implication majeure des dérivés nucléotidiques dans les mécanismes de reconnaissance des phosphoantigènes, puisque l’ApppI est capable de lier l’ATP synthase de façon stable.
Pour finir, la dernière partie présentera les données obtenues en collaboration avec la thématique développée par Laurent Martinez sur le métabolisme nucléotidique à la surface des cellules. Cette collaboration m’a permis de développer des connaissances théoriques et pratiques sur l’activité enzymatique de l’ATP synthase et s’est avérée utile pour l’étude des propriétés de l’ApppI. Elle ouvre également des perspectives potentiellement intéressantes pour l’étude du métabolisme des dérivés nucléotidiques des phosphoantigènes.
Enfin, la partie Discussion et Perspectives sera consacrée à une conclusion générale de nos résultats, placés dans le contexte des données de la littérature. De futurs travaux, visant à caractériser de façon plus précise les mécanismes impliqués dans l’expression et la reconnaissance de l’Ecto-F1-ATPase ainsi que dans la présentation des phosphoantigènes par ce complexe, seront proposés.
I
I
N
N
T
T
R
R
O
O
D
D
U
U
C
C
T
T
I
I
O
O
N
N
B
B
I
I
B
B
L
L
I
I
O
O
G
G
R
R
A
A
P
P
H
H
I
I
Q
Q
U
U
E
E
C
C
H
H
A
A
P
P
I
I
T
T
R
R
E
E
I
I
-L
L
E
E
S
S
Y
Y
S
S
T
T
E
E
M
M
E
E
I
I
M
M
M
M
U
U
N
N
I
I
T
T
A
A
I
I
R
R
E
E
D
D
’
’
u
u
n
n
e
e
v
v
u
u
e
e
d
d
’
’
e
e
n
n
s
s
e
e
m
m
b
b
l
l
e
e
a
a
u
u
x
x
l
l
y
y
m
m
p
p
h
h
o
o
c
c
y
y
t
t
e
e
s
s
T
T
n
n
o
o
n
n
-
-
c
c
o
o
n
n
v
v
e
e
n
n
t
t
i
i
o
o
n
n
n
n
e
e
l
l
s
s
Introduction I : Le système immunitaire
Ce chapitre d’introduction est destiné à offrir une vue globale du système immunitaire. Il sera concentré sur la description succincte du système immunitaire des vertébrés supérieurs (les organismes les précédant dans l’évolution ne disposant généralement que de certains de ses éléments ou de systèmes « primitifs ») et des partenaires cellulaires et moléculaires participant à son organisation. Il fait intervenir de nombreux organes et une incroyable diversité de populations cellulaires, qui interagissent avec la totalité des cellules de l’organisme. Son rôle premier est fondamental : défendre l’organisme (le soi) contre les agents pathogènes (le non soi), ainsi que l’épurer des cellules stressées, endommagées ou cancéreuses (le soi modifié). Il participe également au maintien de l’intégrité des tissus en régulant leur homéostasie. Ses caractéristiques le rendent malheureusement responsable d’effets secondaires indésirables comme le développement de pathologies telles que les maladies auto-immunes ou inflammatoires, ainsi que le rejet de greffes. L’objet de cette thèse étant l’étude d’une sous-population lymphocytaire T non-conventionnelle particulière, les lymphocytes T Vγ9/Vδ2, nous partirons d’une vision globale et succincte des différentes composantes du système immunitaire pour finalement atteindre la description plus détaillée des lymphocytes T non-conventionnels, introduction au chapitre II qui portera sur les lymphocytes T de type γδ.
1) Généralités
1.1) Organes et cellules
Les organes impliqués dans la genèse et le fonctionnement du système immunitaire sont généralement divisés en deux catégories : les organes lymphoïdes primaires (ou centraux) et secondaires (ou périphériques). Les organes primaires (moelle osseuse et thymus) correspondent aux lieux de développement et de différenciation des cellules du système immunitaire alors que les organes secondaires (rate, ganglions lymphatiques et formations
Introduction I : Le système immunitaire
lymphoïdes associées aux tissus telles que les amygdales ou les plaques de Peyer) sont des centres fonctionnels, assurant la rencontre des lymphocytes avec les antigènes et le développement de la réponse immunitaire dite adaptative.
Les cellules du système immunitaire (Figure 1) comprennent deux lignées majeures (myéloïde et lymphoïde) et sont séparées en deux groupes fonctionnels, formant l’immunité innée et l’immunité adaptative. L’immunité innée est caractérisée par une réponse rapide et directe. C’est la première ligne de défense cellulaire de l’organisme contre les pathogènes et elle est de ce fait souvent associée aux tissus exposés au milieu extérieur. Elle regroupe les cellules phagocytaires (telles que les polynucléaires neutrophiles, les monocytes/macrophages et les cellules dendritiques), les cellules dites « autres » (polynucléaires éosinophiles et basophiles, mastocytes, cellules Natural Killer ou NK) et enfin les lymphocytes non-conventionnels (Natural Killer T cells ou NKT, lymphocytes T γδ, et les Mucosal Associated Invariant T cells ou MAIT), même si ces derniers peuvent être considérés comme étant à la frontière innée / adaptative.
L’immunité adaptative est caractérisée par une réponse plus lente, mais disposant d’une mémoire, c’est-à-dire que des cellules spécifiques d’un antigène préalablement activées vont persister, ce qui permettra une réponse plus rapide en cas de nouvelle rencontre avec l’antigène. La mémoire du système immunitaire est la base de la vaccination. Le système adaptatif comprend les lymphocytes T αβ (dits conventionnels) et les lymphocytes B.
Il existe de nombreuses interactions entre l’immunité innée et l’immunité adaptative, la principale étant la capacité de certaines cellules de la première à activer celles de la seconde
via le mécanisme de présentation d’antigènes. Ce mécanisme implique généralement les
phagocytes (macrophages et cellules dendritiques principalement) d’une part, et les lymphocytes T d’autre part.
Introduction I : Le système immunitaire
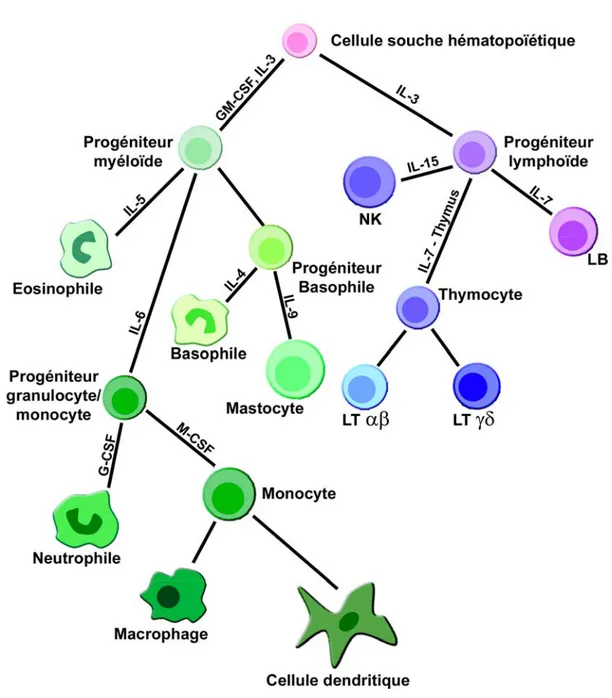
Figure 1. Les cellules du système immunitaire. Adapté de (1)
Le système immunitaire comporte de très nombreuses sous-populations cellulaires, dont les principales familles sont décrites ici. En teintes bleues, les membres de la lignée lymphoïde, en teinte verte, ceux de la lignée myéloïde. A noter que les populations les plus différenciées de cet arbre comportent également de nombreuses sous-populations différentes pouvant se distinguer par des phénotypes moléculaires très variés.
Quelques cytokines importantes pour leur différenciation sont mentionnées.
LB : Lymphocyte B, LT : Lymphocyte T, NK : Natural Killer, IL : Interleukine, CSF : Colony Stimulating Factor (G : Granulocyte, M : Monocyte).
Introduction I : Le système immunitaire
1.2) Molécules fondamentales du système immunitaire : quelques exemples
Le système immunitaire s’appuie sur une grande variété de molécules indispensables à son fonctionnement. Nous verrons ici quelques exemples principaux pertinents pour les sujets traités au cours des prochains chapitres, cette liste n’étant bien évidemment pas exhaustive mais visant à donner une base moléculaire pour la compréhension du fonctionnement général du système immunitaire.
Les cellules de l’immunité adaptative, ainsi que certaines cellules de l’immunité innée, sont caractérisées par l’expression de récepteurs dédiés à la reconnaissance de molécules précises. C’est le cas des lymphocytes B qui expriment un récepteur B à l’antigène (B Cell Receptor ou BCR) ainsi que des anticorps, et des lymphocytes T exprimant un récepteur T (T Cell Receptor ou TCR). Ces récepteurs sont extrêmement spécifiques d’un antigène donné et font l’objet d’une incroyable variabilité. On trouve également des récepteurs moins spécifiques, appelés Toll-Like Receptors (TLR), qui reconnaissent des motifs moléculaires généraux. Par exemple, certains vont être spécialisés dans la reconnaissance d’ARN viraux, ou de polymères bactériens. Tous ces récepteurs jouent un rôle central dans l’activation des cellules du système immunitaire et sont complétés par de nombreux corécepteurs, molécules d’adhésion, de co-stimulation, etc.
Comme mentionné précédemment, il existe des cellules spécialisées dans la présentation des antigènes. Ces cellules vont donc capturer un pathogène, et apprêter les antigènes pour les présenter aux lymphocytes T. Ce mécanisme fait intervenir d’une manière générale des structures présentatrices, les plus connues et les mieux caractérisées étant les molécules du Complexe Majeur d’Histocompatibilité « classique ». Ces protéines, regroupées en deux familles (classe I et classe II) lient des antigènes peptidiques et sont reconnues par le TCR des
Introduction I : Le système immunitaire
lymphocytes T αβ conventionnels. Chez l’homme, le CMHI regroupe les isotypes HLAA, -B, et -C (dits classiques ou CMH-Ia) et -E, -F et -G (non-classiques ou CMH-Ib), généralement reconnus par le TCR des lymphocytes T cytotoxiques et/ou les NKR (Natural Killer Receptors) ; le CMH-II regroupe HLA-DP, DQ et DR et il est reconnu par le TCR des lymphocytes T auxiliaires. Le CMH est extrêmement polymorphe, permettant ainsi au système immunitaire de réagir contre une grande variété de pathogènes, et d’assurer la survie globale d’une espèce en permettant à une proportion non négligeable d’une population de se défendre contre un large spectre de pathogènes. Cette grande variabilité est par ailleurs la cause principale du rejet de greffe, comme le terme « Histocompatibilité » l’indique.
Les molécules du CMH-I, ainsi que d’autres protéines qui leur sont apparentées, ont également la propriété de moduler l’activation de la quasi totalité des lymphocytes T (conventionnels et non-conventionnels) ainsi que des cellules NK, via leur interaction avec des récepteurs appelés Natural Killer Receptor (ou NKR) qui peuvent transduire des signaux inhibiteurs ou activateurs selon le récepteur considéré. Par exemple, l’hétérodimère CD94/NKG2A reconnaît HLA-E et transmet des signaux inhibiteurs. L’absence d’expression de molécules du CMH-I à la surface d’une cellule étant généralement, hormis cas particuliers, le signe d’une infection ou d’un « soi modifié » (les virus par exemple ont tendance à inhiber l’expression de ces molécules pour échapper aux lymphocytes T cytotoxiques), il n’y aura plus de signaux inhibiteurs ce qui peut favoriser l’activation d’une cellule effectrice. Au contraire, CD94/NKG2D reconnaît des molécules apparentées au CMH-I et dites « de stress » telles que MICA/B, et transmet des signaux activateurs. L’activation d’une cellule effectrice est donc déterminée par le bilan de signaux transmis par une grande variété de récepteurs.
Les cytokines et chimiokines jouent également un rôle central dans l’organisation des défenses immunitaires. Elles sont regroupées en plusieurs superfamilles (telles que les
Introduction I : Le système immunitaire
interleukines, interférons, etc) pouvant compter des dizaines de membres. Elles assurent de nombreux rôles fondamentaux et indispensables au fonctionnement du système immunitaire : différenciation et activation cellulaires, migration, communication entre cellules, etc. Les cytokines sont classées en 2 groupes antagonistes, pro-inflammatoires (Th1) ou anti-inflammatoires (Th2), et la plupart des cellules sont spécialisées dans la production d’un seul de ces groupes. La grande majorité des cytokines a des effets pléiotropes, synergiques et redondants.
1.3) Déclenchement de la réponse immunitaire
D’une manière générale, les barrières naturelles de l’organisme protègent contre la très grande majorité des pathogènes, de manière physique (en s’opposant simplement à leur entrée), biochimique (pH, enzymes) ou biologique (par exemple, la présence d’une flore microbienne naturelle, non pathogène, limite le développement de populations pathogènes par simple compétition pour les ressources disponibles). Les réponses rapides et à large spectre, bien que peu spécifiques, des cellules de l’immunité innée contrôlent ensuite la majorité des pathogènes ayant pénétré les barrières mentionnées précédemment.
En cas d’échec de ces deux systèmes, la réponse immunitaire adaptative va se déclencher. Elle implique de nombreux acteurs et se déroule en plusieurs phases précises et ordonnées aboutissant à l’activation, la prolifération et la mise en route des fonctions effectrices de cellules spécifiques du pathogène ciblé, les lymphocytes T et B (Figure 2).
La peau et les muqueuses sont colonisées par de nombreuses cellules dendritiques résidant en permanence dans ces tissus, dans un état immature. L’entrée d’un pathogène déclenche rapidement une réponse inflammatoire locale se traduisant par l’expression de molécules d’adhésion par l’endothélium vasculaire, de chimiokines et de cytokines.
Introduction I : Le système immunitaire
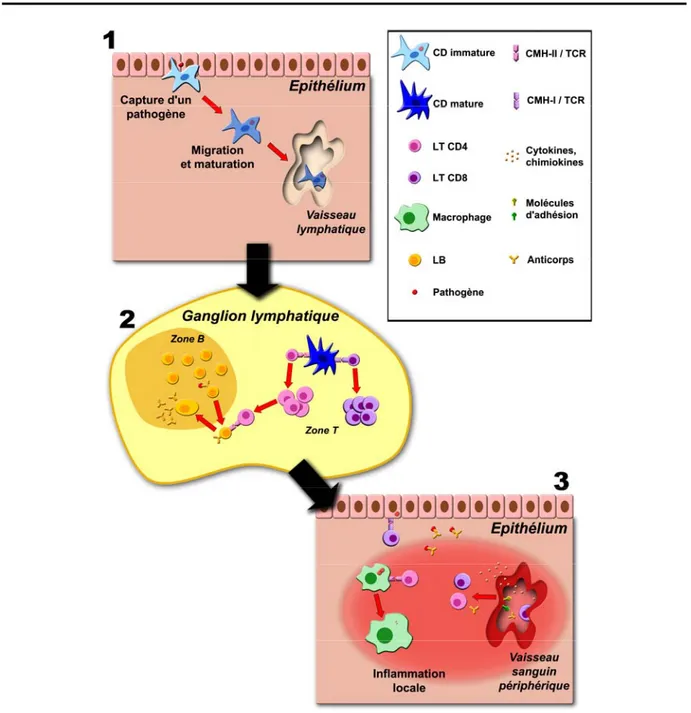
Figure 2. Vue générale de la réponse immunitaire. Adapté de (1)
(1) Une cellule dendritique (CD) capture un pathogène et migre vers un ganglion lymphatique
proche, en passant d’un phénotype phagocytaire à « présentateur ».
(2) La CD active les lymphocytes T (LT). Les CD8 migrent vers la zone infectée, comme les
CD4 qui participent également à l’activation des lymphocytes B (LB). Ces derniers sortent du follicule primaire (zone B) après reconnaissance et phagocytose du pathogène pour rencontrer les LT CD4 de même spécificité, puis y retournent pour finir leur différenciation et sécréter des anticorps.
(3) Les lymphocytes atteignent la zone infectée balisée par des molécules d’adhésion et un
gradient de cytokines et chimiokines. Les CD8 détruisent les cellules infectées, les CD4 participent à l’activation des macrophages. Les anticorps neutralisent les pathogènes, et facilitent leur élimination via la phagocytose ou le complément.
Introduction I : Le système immunitaire
Dès qu’une cellule dendritique capture un pathogène, elle migre vers le ganglion lymphatique le plus proche et effectue sa maturation au cours de son trajet. Les protéines du pathogène seront clivées en peptides courts chargés sur les molécules du CMH. Ainsi, la cellule dendritique activera les lymphocytes T naïfs qui vont proliférer et exprimer des récepteurs pour certaines chimiokines et molécules d’adhésion, leur permettant de migrer vers le lieu de l’infection et de traverser l’endothélium pour éliminer les pathogènes.
Les lymphocytes B quant à eux peuvent être activés soit par les lymphocytes auxiliaires exprimant le corécepteur CD4, soit par des antigènes solubles, polymériques, dits « thymo-indépendants » (car ne nécessitant pas l’intervention des lymphocytes T). Ils comprennent eux aussi diverses sous-populations qui ne seront pas détaillées ici. Ils sécrèteront les anticorps à distance du site d’infection, en restant dans les organes lymphoïdes secondaires.
Cette description succincte de la réponse immune est bien évidemment très simplifiée et restreinte aux phénomènes principaux. Elle fait intervenir de nombreux autres types cellulaires et des mécanismes bien plus variés et complexes.
2) Les lymphocytes T
Quel que soit leur sous-type, tous les lymphocytes T sont caractérisés par l’expression du TCR, qui ne se limite pas au récepteur antigénique seul, mais comprend également les molécules de signalisation γ, δ, ε et ζ formant le complexe CD3 (Figure 3). Le type de récepteur antigénique (αβ ou γδ), l’expression (ou non) des corécepteurs CD4 ou CD8, ainsi que d’autres récepteurs, détermine le sous-type général ainsi que les fonctions d’un lymphocyte T donné. On sépare cette population en deux groupes, les lymphocytes T conventionnels (αβ), et les lymphocytes T non-conventionnels qui comprennent les lymphocytes T γδ, ainsi que les lymphocytes αβ de type NKT et les MAIT.
Introduction I : Le système immunitaire
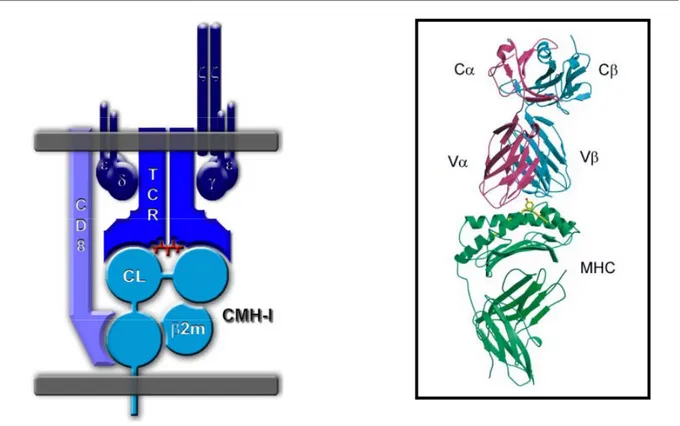
Figure 3. Structure générale du TCR : Exemple du TCR αβ d’un lymphocyte T CD8+
On parle souvent du TCR comme étant l’association de deux chaînes αβ ou γδ, mais le TCR dans son ensemble comprend également les sous-unités CD3 (γ, δ, ε et ζ) portant des motifs de type ITAM et participant à la transduction du signal, en association avec de nombreuses kinases. Le TCR peut être également couplé (en fonction du type de lymphocyte T considéré) à un corécepteur de type CD4 ou CD8 renforçant l’interaction avec le CMH (de classe II ou I, respectivement).
Ici, le TCR d’un lymphocyte T CD8 reconnaît un peptide antigénique (en rouge) présenté par le CMH-I composé d’une chaîne lourde (CL) formée de trois domaines immunoglobuliniques α1, α2 et α3, et de la beta-2-microglobuline (β2m).
Le corécepteur CD8 interagit avec le domaine α3 (très conservé) tandis que le TCR reconnaît à la fois le peptide antigénique et les domaines α1 et α2 (présentant une importante variabilité) du CMH-I via des domaines appelés boucles CDR pour Complementarity Determining Region (CDR3 pour le peptide et CDR1 et 2 pour le CMH-I). La chaîne α du TCR reconnaît le domaine α2 du CMH-I ; la chaîne β, le domaine α1.
Encadré : structure tridimensionnelle du complexe TCR / CMH-I. Vert : CMH-I, Jaune : Peptide, Bleu et Rose : Chaînes β et α du TCR (2).
Les populations non-conventionnelles faisant l’objet de la section suivante, nous nous concentrerons ici sur des généralités sur les lymphocytes T et sur les sous-populations de lymphocytes T conventionnelles αβ.
Introduction I : Le système immunitaire
2.1) Développement
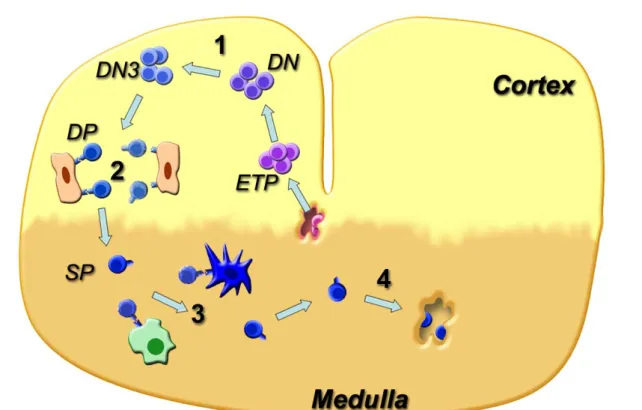
Les lymphocytes T se développent dans le thymus, à partir de progéniteurs lymphoïdes pluripotents (ETP, Early T cell Progenitors) provenant de la moelle épinière ou du foie. Ils sont appelés thymocytes une fois qu’ils sont entrés dans le cortex thymique, et qualifiés de Double Négatifs (DN) à ce stade car ils n’expriment pas les corécepteurs CD4 et CD8. Ce développement s’effectue par étapes clés, comprenant deux points de contrôle majeurs, les sélections positive et négative, pour former un répertoire T efficace et tolérant, c’est-à-dire qu’il ne doit pas reconnaître le « soi » comme étranger.
L’évènement initiant la différenciation des thymocytes est le réarrangement des gènes du TCR. Ces gènes sont morcelés et se composent de très nombreux segments géniques, appartenant à plusieurs groupes : V (Variable), D (Diversité), J (Jonctionnel) et C (Constant). Cette propriété est identique pour les TCR αβ ou γδ (les segments D n’existant néanmoins que pour les chaînes β et δ), et le BCR (et par extension les immunoglobulines). Les gènes du TCR étant morcelés, leur expression nécessite un mécanisme de remodelage génique irréversible particulier et unique aux gènes des récepteurs antigéniques, appelé recombinaison somatique (Figure 4). Dans tous les cas, elle fait intervenir les recombinases RAG-1 et -2 (Recombination Activating Gene) qui vont aléatoirement assembler un gène fonctionnel à partir d’un segment V, D, et J, dans l’ordre D+J puis V+DJ (ou V+J directement pour les chaînes α et γ). Le segment C ainsi que le peptide signal (L) seront quant à eux assemblés aux autres segments par épissage après la transcription (3). Ce processus étant aléatoire et basé sur une grande variété de segments géniques (Tableau 1), il permet d’obtenir une incroyable diversité de TCR différents. D’autres processus de la recombinaison introduisent une variabilité supplémentaire, en ajoutant ou en retirant des nucléotides au niveau des jonctions entre les segments V, D et J.
Introduction I : Le système immunitaire
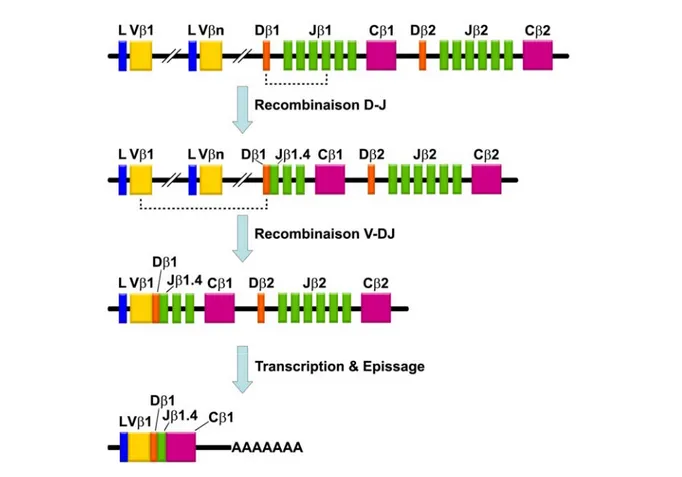
Figure 4. Structure et recombinaison d’un locus TCR : Exemple de la chaîne β.
Adapté de (1)
Les recombinases RAG 1 et 2 effectuent la recombinaison des gènes du TCR dans l’ordre indiqué. Les autres segments J, D et C ainsi que les introns sont éliminés par épissage. Au niveau des jonctions, des endonucléases ainsi que des ADN polymérases et la Terminal deoxyribonucleotidyl Transferase (TdT) retirent ou ajoutent respectivement des nucléotides, ce qui contribue à la variabilité du TCR. Les segments sont assemblés aléatoirement et toutes les combinaisons sont possibles, bien que ne codant pas forcément pour un TCR fonctionnel.
Locus Localisation chromosomique V D J C
α 14q11.2 * 54 ** 0 61 1
β 7q34 64-67 2 14 2
γ 7p14 12-15 0 5 2
δ 14q11.2 * 8 ** 3 4 1
Tableau 1. Organisation des loci TCR
Ce tableau récapitule le nombre de segments géniques pour chaque locus TCR.
* Le locus δ est situé dans le locus α ** 5 segments V sont communs aux loci α et δ