NEIL MARCHAND
CROISSANCE RADIALE DE LA PRUCHE (TSUGA
CANADENSIS) ET DU PIN BLANC (PINUS STROBUS)
DANS LA FORÊT TEMPÉRÉE DU QUÉBEC EN
RELATION AVEC LES PERTURBATIONS
FORESTIÈRES ET LE CLIMAT
Thèse présentée
à la Faculté des études supérieures et postdoctorales de l’Université Laval dans le cadre du programme de doctorat en Sciences géographiques
pour l’obtention du grade de Philosophiæ Doctor (Ph. D.)
DÉPARTEMENT DE GÉOGRAPHIE
FACULTÉ DE FORESTERIE, DE GÉOGRAPHIE ET DE GÉOMATIQUE UNIVERSITÉ LAVAL
QUÉBEC
2013
Résumé
La présente thèse porte sur la croissance radiale de la pruche (Tsuga canadensis L. Carr.) et du pin blanc (Pinus strobus L.), deux espèces arborescentes de la forêt tempérée du sud du Québec où de multiples perturbations interviennent dans la dynamique forestière. Fondée sur une analyse dendroécologique et dendroclimatique, cette recherche comprend trois volets hautement complémentaires qui documentent les relations entre la croissance radiale chez ces deux espèces et les perturbations forestières, à savoir le prélèvement sélectif d’individus, la défoliation lors des épidémies de la tordeuse des bourgeons de l’épinette (TBÉ) (Choristoneura fumiferana (Clemens), ainsi que le climat.
Le premier volet présente les longues séries dendrochronologiques de pruche et de pin blanc confectionnées en Mauricie à l’aide d’arbres vivants et de diverses catégories de bois morts, surtout des billes de bois de flottage extraites de quelques lacs du Parc national de la Mauricie. Les années d’abattage montrent que le pin blanc a été exploité à des fins domestiques au moins 30 ans avant l’émission des premiers droits de coupe, c’est-à-dire depuis au moins 1800, et 70 ans après la disparition présumée des pinèdes dans la région. Les séries montrent des reprises de croissance découlant d’un prélèvement sélectif au sein des forêts. La chronologie des épidémies de la TBÉ établie à l’aide de la pruche, qui est pourtant une espèce hôte secondaire de la TBÉ, est la plus longue produite à ce jour dans l’est du Canada. Les données obtenues de plusieurs régions du Québec méridional (Mauricie, Outaouais et Lotbinière) suggèrent que la pruche peut subir une défoliation sévère lors des épidémies, là où des peuplements de sapin baumier et d’épinette blanche abondent à proximité. À l’échelle régionale, la fréquence des épidémies de la TBÉ augmenterait, vraisemblablement en lien avec l’augmentation de l’abondance du sapin dans les peuplements en régénération.
Le second volet de la thèse a permis d’évaluer la réponse climatique de la pruche et du pin blanc dans les forêts tempérées du sud du Québec. Les analyses dendroclimatiques ont montré que la pruche peut être un bon indicateur du climat si elle a crû dans un milieu où les perturbations naturelles et anthropiques ont été légères. Dans les études dendroclimatiques, on ciblera de préférence des peuplements situés dans des régions où dominent les forêts de feuillus et dont l’historique des perturbations est connu. Le pin blanc
est moins sensible aux perturbations biotiques que la pruche, mais la qualité de la réponse climatique chez ce conifère dépend des contraintes hydriques (comme en Outaouais) ou thermiques (comme sur l’île d’Anticosti).
Le troisième volet porte sur la formation de faux cernes chez le pin blanc dans les collines de l’Outaouais, une réponse dendroanatomique à un stress hydrique. Des chronologies de faux cernes ont été élaborées pour deux groupes de sites (sur affleurements rocheux et sur dépôts minces). La fréquence la plus élevée a été obtenue en 1955, 1959 et 1988 (62 % à 90 % des tiges). Les facteurs prédisposant à la formation de faux cernes sont l’âge cambial des tiges et les conditions de drainage rapide sur les affleurements rocheux. Les facteurs déclencheurs sont des événements climatiques (faibles précipitations) survenant pendant la saison de croissance.
Abstract
Our dissertation documents the radial growth among eastern hemlock (Tsuga canadensis L. Carr.) and white pine (Pinus strobus L.), two tree species of the southern Québec temperate forests, where several disturbances get involved in stand dynamics. Based on a dendroecological and dendroclimatical approach, this study includes three highly complementary sections which address the relationship between radial growth among these two tree species and forest disturbances, i.e., selective tree harvest, defoliation during spruce budworm (SBW) (Choristoneura fumiferana (Clemens)) outbreaks, and climate.
The first section describes eastern hemlock and white pine long tree-ring series which were built from both living trees and dead wood, mainly submerged floated wood extracted from several lakes in the Mauricie National Park. One can deduce from white pine logging years that this tree had been harvested for domestic use 30 years before the first issue of logging rights, i.e., since the 1800s onwards, and about 70 years after presumed decimation of pine forests in the area. Tree ring series show major growth release which likely resulted from selective harvest in forest stands. Our chronology of SBW outbreaks established from eastern hemlock tree-ring series is the longest chronology produced so far in eastern Canada, although hemlock is a secondary host of SBW. Series from several regions (Mauricie, Outaouais and Lotbinière) suggest that eastern hemlock may experience severe defoliation and radial growth suppression in areas where primary hosts of SBW are abundant. At the regional scale, the frequency of SBW outbreaks increases likely in response to an increased abundance in balsam fir in second-growth stands.
In this section, we attempted to evaluate climate responsiveness among eastern hemlock and white pine growing in the southern Quebec Temperate Forest. Dendroclimatic analyses showed that hemlock radial growth may be a good climatic indicator in stands where natural and anthropogenic disturbances were light. Sampling strategies when conducting dendroclimatic studies should focus on trees growing in hardwood-dominated stands and areas where past disturbances have beendocumented. White pine is less sensitive to biotic disturbance than eastern hemlock but the climate responsiness of this conifer depends on constraints associated with water deficit or thermal stress.
The third section deals with false ring formation in the white pine of the Outaouais Hills, a dendroanatomic response to water stress. False ring chronologies were established for two groups of sites (rocky outcrops and shallow soils). The years 1955, 1959, and 1988 were those with the highest occurrence of false rings (62% to 90% of all stems). Factors predisposing false ring formation include the cambial age of stems bearing false rings and drainage conditions (dry rocky outcrop sites). Triggering factors were climatic events (low precipitation) occurring during the growing season.
Avant-propos
Cette thèse comprend cinq chapitres dont un, rédigé et publié en anglais, a fait l’objet d’un article scientifique (Chapitre 4). Les chapitres 2 et 3 feront aussi l’objet de publications distinctes, après quelques remaniements. Bien que complémentaires, les chapitres qui forment cette thèse peuvent être lus séparément. Par conséquent, le lecteur voudra bien faire preuve d’indulgence quant à certaines répétitions dans le texte. L’auteure est la principale rédactrice des chapitres et des illustrations, ainsi que la responsable des travaux de terrain et de la majorité des analyses en laboratoire. En sa qualité de directrice Louise Filion (biogéographe) a participé au cheminement de cette thèse et à l’amélioration de son contenu.
Chapitre 1 : Introduction générale
Chapitre 2 : Analyse dendroécologique de la pruche (Tsuga canadensis) et du pin blanc (Pinus strobus) en relation avec la coupe forestière et les épidémies d’insectes défoliateurs dans le Parc national de la Mauricie (Québec, Canada).
Chapitre 3 : Analyse dendroclimatique du pin blanc (Pinus strobus) et de la pruche (Tsuga canadensis) dans la forêt tempérée du Québec, dans un contexte de perturbations multiples. Chapitre 4 : Marchand, N. et Filion, L. 2012. False rings in the white pine (Pinus strobus) of the Outaouais Hills, Québec (Canada), as indicators of water stress. Publié dans Canadian Journal of Forest Research, 42 : 12-22
Remerciements
Je tiens d’abord à remercier ma directrice de recherche, Louise Filion, de m’avoir confié ce travail passionnant, mais surtout de m’avoir soutenue dans sa réalisation. Je voudrais souligner la rapidité avec laquelle elle a toujours répondu à mes questionnements. La franchise et la pertinence de ses commentaires ainsi que la qualité de ses corrections ont grandement contribué à ma formation et sa grande disponibilité m’a permis de rester motivée pendant toute la durée de cette recherche.
Je voudrais exprimer ma gratitude aux membres de mon comité, le professeur Marc Saint-Hilaire (département de Géographie, Université Laval), et la professeure Diane Saint-Laurent (département des Sciences environnementales, Université du Québec à Trois-Rivières) pour leur encadrement et leurs conseils judicieux. Merci également au professeur Hubert Morin (département des Sciences fondamentales, Université du Québec à Chicoutimi), membre externe du jury qui a aussi évalué cette thèse.
Je souhaite remercier très sincèrement Michel Plante pour son accueil au Parc de la Mauricie et ses encouragements de tous les instants. M. Plante a utilisé tous les moyens dont il disposait pour faciliter le déroulement de mes travaux sur le terrain et il a volontiers partagé ses vastes connaissances du parc. Je remercie également François Auger, dont je n’oublierai ni les petites chansons et ni les itinéraires toutistiques. L’aide précieuse et indispensable de Guillaume Caron, Michel Mongrain et Gilles Bellemare, « les gars » de l’équipe Billot-canot, a été très appréciée, ainsi que celle d’Yves Marcouiller et de Simon Plante. En plus de partager votre expérience et votre amour des forêts et des lacs du parc, vous avez tous contribué à faire de mes travaux de terrain, non seulement une réussite, mais surtout une expérience inoubliable.
Un grand merci à Ann Delwaide et à Luc Cournoyer, deux professionnels de recherche au CEN qui ont été des personnes ressources avec qui j’ai eu grand plaisir à travailler. Merci donc Ann pour les discussions et réflexions devant des échantillons et Luc pour les innombrables versions du logiciel Dendro, les formules Excel et tout le reste !
Je tiens également à mentionner le travail de mes nombreux assistants, tant sur le terrain qu’en laboratoire : Emmanuel Tousignant, Audrée Mercier-Remillard, Bryan Synkunas, mais surtout David Carignan et Anne Beaudoin pour leur patience dans les analyses en laboratoire qui, ont toujours été de très grande qualité. Mes remerciements s’adressent également à Isabelle Beaudoin-Roy et aux « Amis du parc de la Gatineau » pour leur aide financière.
Une pensée toute particulière va au professeur Pierre Pech, mon directeur de maîtrise à Paris I, que je remercie de m’avoir accompagné dans mes premier pas sur le chemin de la recherche, de la dendrochronologie et du Québec.
Un doctorat est un travail qui vous poursuit jour et nuit. C’est pourquoi je souhaite exprimer ma gratitude à mes amis de Québec qui m’ont appuyée pendant toutes ces années : Marianne, Julie, Jonathan, Francis, Max, Geneviève, Marc et Linda, ma super belle-maman. Un merci particulier va à Corinne Vezeau pour son amitié inconditionnelle, son aide dans la rédaction, ses encouragements et les excellents moments que nous avons passés ensemble. Je tiens à remercier mes amis et ma famille en France, dont ma maman, ma mamie Poulet et Ariane, qui m’ont toujours soutenue dans mes choix même s’ils impliquaient l’éloignement. Je tiens à remercier chaleureusement tout ceux qui m’ont fait le plaisir de venir partager mon expérience en visitant mon Québec, en particulier mes frères Adam et Léo, mes neveux Mars et Lala et mes amis Angeline et Arnaud. Il est difficile de trouver des mots assez forts pour remercier Jean-François Verville, mon lecteur le plus fidèle, mon cartographe, mon amoureux et équipier de toutes les circonstances.
Enfin, j’ai une pensée toute spéciale pour mon père qui aurait aimé ce travail et la vie qui vient avec, dans une cabane au Canada.
Il faut prendre le taureau par les cornes, parce que par la queue, il n’aime pas ça. Charles Patenaude
Table des matières
Résumé ... ii
Abstract ... iv
Avant-propos... vi
Remerciements ... vii
Table des matières...x
Liste des figures...xiv
Liste des tableaux ... xviii
Chapitre 1. Introduction générale ...20
Chapitre 2 : Analyse dendroécologique de la pruche (Tsuga canadensis) et du pin blanc (Pinus strobus) en relation avec la coupe forestière et les épidémies d’insectes défoliateurs dans le Parc national de la Mauricie (Québec, Canada) ...36
Résumé ...36
Abstract ...38
2.1. Introduction...39
2.2. Méthodes ...43
2.2.1 Région et sites d’étude...43
2.2.2. Échantillonnage dendrochronologique ...45
2.2.2.1. Bois de flottage ...45
2.2.2.2. Arbres vivants et autres catégories d’arbres morts ...47
2.2.3. Préparation des échantillons, analyses en laboratoire et élaboration des séries dendrochronologiques ...48
2.2.4. Évaluation du degré d’altération des billes en fonction des conditions de conservation ...50
2.2.5. Détermination et correction de l’année d’abattage des arbres ...50
2.2.6. Recherche documentaire ...51
2.2.7. Identification dendroécologique des épidémies d’insectes défoliateurs ...51
2.3. Résultats ...54
2.3.1. Série dendrochronologique de la pruche (série Isaïe)...54
2.3.2. Série dendrochronologique du pin blanc (série Houle) ...57
2.3.3. Série dendrochronologique de l’épinette rouge (série Patrick) ...58
2.3.4. Conditions de conservation et degré d’altération des bois lacustres ...59
2.3.5. Années d’abattage du pin blanc et de la pruche ...60
2.3.7. Épidémies d’insectes défoliateurs ...64
2.3.7.1. Chez la pruche ...64
2.3.7.2. Chez l’épinette rouge ...66
2.3.7.3. Périodes de réduction de croissance associées à l’activité des insectes défoliateurs dans Lotbinière et en Outaouais ...66
2.4. Discussion ...71
2.4.1. Particularités du matériel dendrochronologique lacustre ...71
2.4.1.1. Qualité du bois ...71
2.4.1.2 Durée des séries dendrochronologiques ...71
2.4.2 Activités de coupe forestière et données dendroécologiques ...72
2.4.2.1 Le pin blanc ...72
2.4.2.2. La pruche ...75
2.4.3. Liens entre la coupe forestière sélective et la croissance radiale des conifères ...77
2.4.4. Activité des insectes défoliateurs ...79
2.4.4.1. Particularités méthodologiques de l’analyse de l’activité des insectes défoliateurs dans un contexte de perturbations anthropiques ...79
2.4.4.2. Insectes responsables de la défoliation ...79
2.4.4.3. Potentiel d’analyse d’une espèce hôte secondaire de la TBÉ ...81
2.4.4.4 Historique des épidémies dans les régions avoisinant la Mauricie tel qu’établi à l’aide de la pruche...84
2.4.4.5 Historique des épidémies en Mauricie tel qu’établi à l’aide d’une espèce hôte secondaire et dans les régions avoisinantes tel qu’établi à l’aide d’espèces hôtes principales ...85
2.4.4.6. Influence des activités anthropiques sur l’activité de la TBÉ ...87
2.5 Conclusion ...89
2.6. Références...90
Chapitre 3 : Analyse dendroclimatique de la pruche (Tsuga canadensis) et du pin blanc (Pinus strobus) dans la forêt tempérée du Québec, dans un contexte de perturbations multiples ... 102
Résumé ... 102
Abstract ... 103
3.1. Introduction... 104
3.2. Méthodes ... 106
3.2.1. Régions et sites d’étude ... 106
3.2.1.1. Parc de la Gatineau ... 107
3.2.1.3. Réserve écologique de la Rivière-du-Moulin... 109
3.2.1.4. Île d’Anticosti... 110
3.2.2. Échantillonnage dendrochronologique ... 111
3.2.3. Analyses dendroclimatiques ... 113
3.2.3.1. Données météorologiques... 113
3.2.3.2. Fonctions de réponse et fonctions de corrélation ... 114
3.3. Résultats ... 116
3.3.1. Caractéristiques des séries dendrochronologiques ... 116
3.3.1.1. Séries de pruche ... 116
3.3.1.2. Séries de pin blanc ... 119
3.3.2. Comparaison des séries dendrochronologiques des deux espèces entre les régions ... 122
3.3.2.1. Séries de pruche ... 122
3.3.2.2. Séries de pin blanc ... 123
3.3.2.3. Comparaison des séries dendrochronologiques des deux espèces au sein d’une même région ... 123
3.3.3. Relation cernes annuels/climat ... 124
3.3.3.1. La pruche ... 124
3.3.3.2. Le pin blanc ... 126
3.3.3.3 Stabilité des relations cernes annuels/climat... 128
3.4. Discussion ... 130
3.4.1. Relations cernes annuels/climat au sein des régions ... 130
3.4.1.1. Outaouais ... 130
3.4.1.2. Mauricie et Lotbinière ... 132
3.4.1.3. Île d’Anticosti... 135
3.5. Conclusion ... 137
3.6. Références... 138
Chapitre 4 : False rings in the white pine (Pinus strobus) of the Outaouais Hills, Québec (Canada), as indicators of water stress ... 146
Résumé ... 146
Abstract ... 147
4.1 Introduction... 148
4.2. Methods ... 151
4.2.1. Study area and sites ... 151
4.2.3. Laboratory... 152
4.2.4. False ring database ... 152
4.2.5. False ring typology and dendrometric parameters related to their formation ... 153
4.2.6. Meteorological data ... 154
4.2.7. Correlation functions ... 154
4.3. Results ... 156
4.3.1. False ring chronology and frequency ... 156
4.3.2. False ring characteristics... 159
4.3.3. The cambial age of stems at the time of false ring formation ... 160
4.3.4. Soil conditions ... 161
4.3.5. Distribution of precipitation throughout the growing season... 163
4.3.5.1 The driest month of the growing season ... 163
4.4. Discussion ... 165
4.4.1 Predisposing factors ... 165
4.4.2 Triggering factors ... 165
4.4.3. Radial growth and false ring formation ... 167
4.4.4. Geographical distribution of false rings ... 168
4.5. References... 170
Chapitre 5. Conclusion générale ... 174
5.1 Contributions de la thèse ... 174
5.2 Portée de la thèse et perspectives de recherche ... 176
Liste des figures
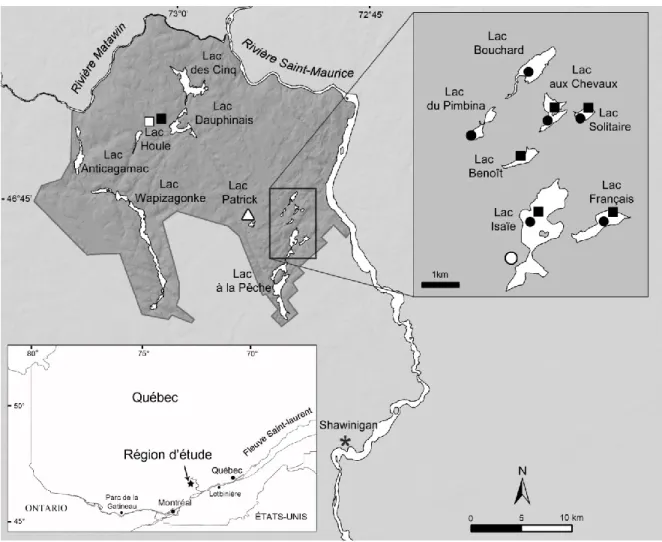
Figure 2.1. Localisation de la région d’étude et des sites d’échantillonnage des billes de bois de flottage et autres catégories d’arbres morts dans le Parc national de la Mauricie : pruche (●), pin blanc (■) et épinette rouge (▲). Les mêmes symboles en blanc représentent les sites d’échantillonnage d’arbres vivants. La carte montre le réseau hydrographique principal et localise la station météorologique de Shawinigan (*). Sur le carton, le site de Lotbinière étudié par Delwaide et Filion (1999) et le parc de la Gatineau sont aussi représentés………45 Figure 2.2. A) Empilement de billes de bois de flottage extraites du lac Isaïe (août 2007).
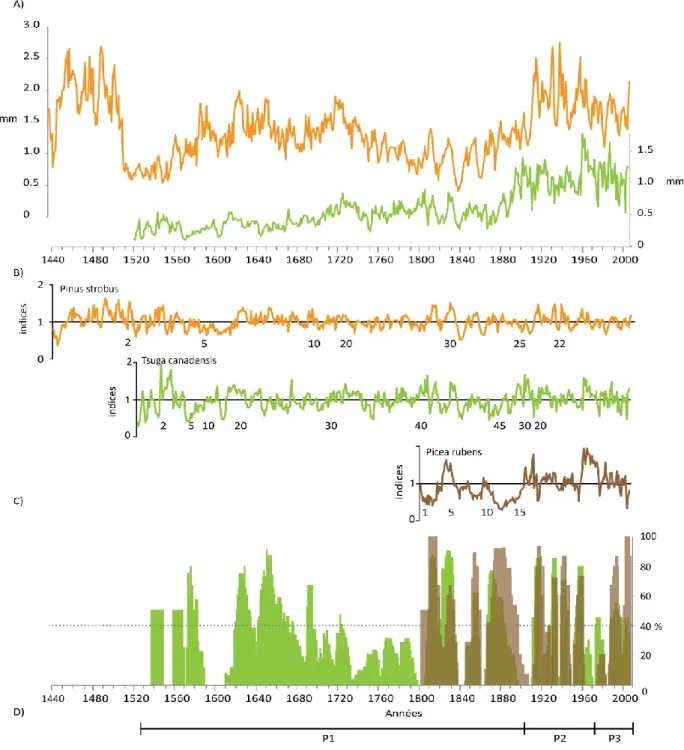
B) Bille de pruche extraite du lac Isaïe dont la partie lisse qui porte encore un fragment d’écorce (1) a été conservé dans les sédiments lacustres. La partie rugueuse (à gauche) a été altérée au contact de l’eau (2). C) Extrémité d’une bille de pruche extraite du lac Isaïe et portant des marques de poinçon LP identifiant la compagnie exploitante, dans ce cas la Laurentide Paper. L’échantillon couvre la période 1717-1906. D) Manœuvres d’extraction d’une bille de pin blanc à l’aide d’un pic, lac Français. L’échantillon couvre la période 1513-1832………..48 Figure 2.3. A) Courbes de la croissance radiale moyenne (mm) du pin blanc (Pinus strobus, en orange) et de la pruche (Tsuga canadensis, en vert) dans le Parc national de la Mauricie. B) Séries dendrochronologiques indicées des trois espèces de conifères. Sous les courbes, le nombre d’échantillons utilisés dans la confection des séries dendrochronologiques est indiqué. C) Histogrammes de fréquence (%) de tiges montrant une réduction de croissance telle qu’identifiée dans le logiciel OUTBREAK (Holmes et Swetnam, 1996) : pruche en vert et épinette rouge en brun clair. Après 1800, lorsque les deux espèces sont touchées, les histogrammes prennent une teinte foncée. D) Périodes (P) historiques retenues pour effectuer l’analyse des données dendroécologiques………56 Figure 2.4. A) Histogramme montrant les années de formation des derniers cernes annuels
présents sur les billes de bois de flottage de pruche (vert et bleu) et de pin blanc (teintes de brun et orangé) dans l’ensemble des sites d’échantillonnage. En vert foncé, les années exactes d’abattage des tiges de pruche avec écorce. Les autres couleurs montrent les années maximales d’abattage déterminées sur des billes dont la partie externe est altérée (sans écorce), par secteur. B) Période d’exploitation par secteur (BEN : lac Benoît; ISA : lac Isaïe; SOL : lac Solitaire; HOU : secteur du lac Houle; BOU : lac Bouchard; PIM : lac du Pimbina; FR : lac Français) et par espèce (pin blanc en jaune, pruche en vert). La zone hachurée correspond à la correction qui a été appliquée afin de tenir compte de l’altération des tiges et des cernes externes manquants. C) Grandes phases d’exploitation de la forêt et principales compagnies exploitantes sur le territoire correspondant au Parc national de la Mauricie………63
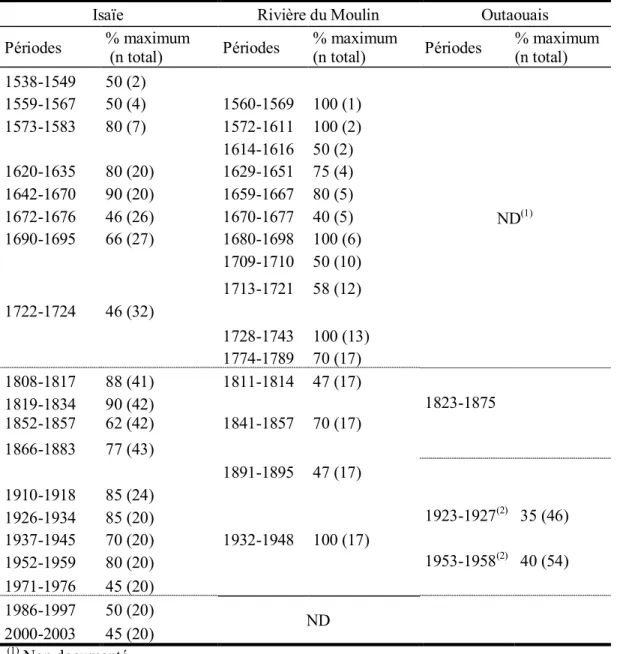
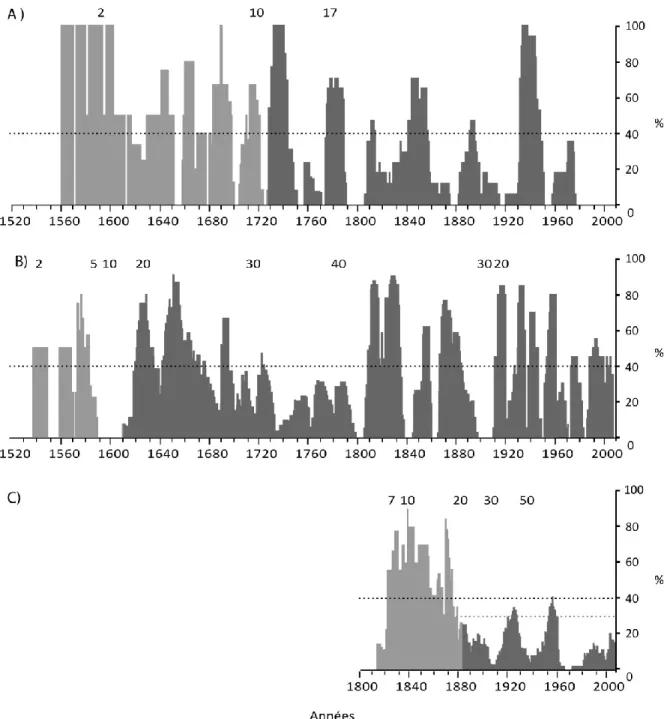
Figure 2.5. Histogrammes de fréquence (%) de tiges montrant une réduction de croissance telle qu’identifiée dans le logiciel OUTBREAK (Holmes et Swetnam, 1996) chez la pruche A) dans Lotbinière (série Rivière-du-Moulin), B) en Mauricie (série Isaïe) et C) dans l’Outaouais (série Belisle). Les chiffres au dessus des histogrammes indiquent le nombre d’échantillons. Le seuil de 40 % retenu afin d’identifier les épidémies est indiqué par un trait en pointillés. En C, le seuil de 30 % a aussi été utilisé après 1880. La partie claire des histogrammes montre la période couverte par un nombre restreint d’échantillons ne permettant pas une bonne interprétation des résultats………..71 Figure 2.6. A) Équarrissage à la hache d’un gros tronc de pin blanc (CIEQ http://mauricie.cieq.ca DIAPO_GRM_165). B) À l’avant-plan, glissoire en travers de la pente, en contrebas d’un site industriel à Shawinigan (CIEQ http://mauricie.cieq.ca N60-38). C) Scène hivernale prise à l’occasion de la construction de la voie ferrée entre Shawinigan et Trois-Rivières, 1906 (CIEQ http://mauricie.cieq.ca DIAPO_GRM_58). D) Scierie à l’usine de la Laurentide Paper à Grand-Mère, vers 1900 (Musée McCord http://www.musee-mccord.qc.ca VIEW-3503. E) Usine de papier de la Laurentide Paper à Grand-Mère et centrale hydroélectrique, 1900 (DIAPO_GRM_007). F) Draveurs piquant les billes de bois dans un rapide en Mauricie, vers 1940 (CIEQ http://mauricie.cieq.ca N 60-69)…..75 Figure 2.7. Périodes d’épidémie de la tordeuse des bourgeons de l’épinette identifiées dans les études dendrochronologiques antérieures à l’aide d’espèces hôtes principales (sapin baumier, épinette blanche) (trame grise) et dans cette étude à l’aide d’une espèce hôte secondaire (pruche) (trame noire). La durée des séries est donnée par les deux traits verticaux externes. Adaptée de Boulanger et Arsenault, 2004…………82 Figure 2.8. Courbes de croissance radiale (mm) A) d’une épinette rouge vivante, B) d’une pruche vivante, C) d’une pruche morte et D) série de référence (non indicée) du pin blanc. Les bandes grises représentent les périodes d’épidémie telles que décelées chez la pruche par l’entremise du logiciel Outbreak (Holmes, 1983)………...85 Figure 3.2. Localisation des régions d’échantillonnage dendrochronologique : pruche ( ● ) et pin blanc ( ■ ). En Outaouais : parc de la Gatineau (2 espèces); en Mauricie : Parc national de la Mauricie (2 espèces); dans Lotbinière : réserve écologique de la Rivière-du-Moulin (pruche) et sur l’île d’Anticosti: bassin de la rivière Prinsta (pin blanc). La carte montre aussi la localisation des stations météorologiques de Chelsea, Shawinigan, Donnacona et Natashquan ( * )……….107 Figure 3.2. Courbes de croissance radiale (mm) (A) et séries indicées de la pruche B)
obtenues des différentes régions d’échantillonnage dendrochronologique. Les chiffres sous les chronologies indicées indiquent le nombre d’échantillons qui forment chacune des séries………..119
Figure 3.3. Courbes de croissance radiale (A) (mm) et séries indicées du pin blanc (B) obtenues des différentes régions d’échantillonnage dendrochronologique. Les chiffres sous les chronologies indicées indiquent le nombre d’échantillons qui forment chacune des séries………..121 Figure 3.4. Coefficients de corrélation (r de Pearson) et coefficients de réponse (valeurs de P) entre les paramètres climatiques (Températures maximums : bâtonnets oranges; précipitations totales : bâtonnets verts) des mois d’avril à août et la largeur des cernes annuels chez la pruche dans les différentes régions d’échantillonnage dendrochronologique ( p < 0,05 ; p < 0,05 récurrent dans les deux types d’analyse)………126 Figure 3.5. Coefficients de corrélation (r de Pearson) et coefficients de réponse (valeurs de
P) entre les paramètres climatiques (Températures maximums : bâtonnets oranges; précipitations totales : bâtonnets verts) des mois d’avril à août et la largeur des cernes annuels chez le pin blanc dans les différentes régions d’échantillonnage dendrochronologique ( p < 0,05 ; p < 0,05 récurrent dans les deux types d’analyse)………128 Figure 3.6: Coefficients de corrélation (r de Pearson) calculés pour des périodes de 20 ans
décalées de 5 ans, entre les paramètres climatiques (Températures maximums et précipitations mensuelles) des mois d’avril à août et la largeur des cernes annuels chez A) la pruche et B) le pin blanc dans les différentes régions d’échantillonnage dendrochronologique. Seuls les coefficients significatifs sont indiqués………….130 Figure 4.1. Location of the study area (inset) and sites within Gatineau Park in the Outaouais Hills, southwestern Québec (Mmodified from Saint-Cyr 1992). The location of Environment Canada’s meteorological station at Chelsea is also shown………...150 Figure 4.2. (A) Histogram showing the number (N) of stems bearing false rings between
1801 and 2006. The three years during which a higher frequency of false rings was recorded are 1955, 1959, and 1988. The number of individuals/ per year used in the chronology is also indicated. The period 1928–2006 (between arrows) was used in this study for a number of statistical analyses. (B–C) Histogram showing the percentage of stems bearing false rings (B) on the rocky outcrop sites (n = 43 to 54) and (C) on shallow soil sites (n = 36 to 40). In B and C, the dotted line corresponds to a 70% frequency of stems bearing false rings……….158 Figure 4.3. Macroscopic view of false rings formed in (A) 1955, (B) 1959, and (C) 1988 (60–80× magnification). The intra-annual bands (shown by the arrows) are darker and denser than the adjacent wood. The symbols ┤ and ├ define the outer limits of the annual ring containing one (A and B) or two (C) false rings………....159 Figure 4.4. Linear relationship between the cambial age of stems at the time of false ring formation and percentage of false rings. The symbols represent ages for which the number of recording stems was ≤65. The number of individuals (N) is also indicated (grey line)………...161
Figure 4.5. Correlation coefficients (Pearson’s r) between monthly maximum temperature (solid bars) and total precipitation (haded bars) and (A) false ring occurrence and (B) mean standardized ring widths (*,p < 0.05)………...163 Figure 4.6. Partial (May to August) ombrothermic diagrams for the three years (A, 1955; B, 1959; C, and 1998) for which a higher frequency of false rings was recorded and the three years containing no or few false rings (D, 1952; E, 1994; F, 2000). The solid and broken lines represent the maximum monthly temperature (T, °C) and the total monthly precipitation (P, mm), respectively………...165
Liste des tableaux
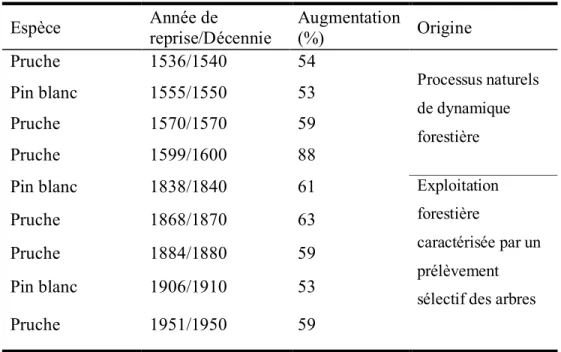
Tableau 2.1. Caractéristiques des séries dendrochronologiques Isaïe (pruche), Houle (pin blanc) et Patrick (épinette rouge)………..55 Tableau 2.2. Reprises importantes de la croissance radiale chez la pruche et le pin blanc,
identifiées à l’aide des méthodes de Lorimer et Frelich (1989) et de Cook (2000)……… ………..58 Tableau 2.3. Nombre de cernes manquants chez la pruche et le pin blanc dans chaque
secteur et selon les conditions de conservation……….61 Tableau 2.4. Périodes de réduction de croissance chez la pruche et l’épinette rouge identifiées dans le logiciel Outbreak et pourcentage maximum et moyen d'individus affectés par la défoliation en période d’épidémies de la TBÉ………...66 Tableau 2.5. Périodes de réduction de croissance chez la pruche en Mauricie (série Isaïe), Lotbinière (série Rivière-du-Moulin) et Outaouais (série Lauriault) identifiées dans le logiciel Outbreak et pourcentage maximum d'individus affectés par la défoliation en période d’épidémies de la TBÉ. Les traits pointillés représentent les limites des périodes d’analyse communes………...69 Tableau 3.1. Données des quatre stations météorologiques d’Environnement Canada utilisées dans l’analyse dendroclimatique………...109 Tableau 3.2. Caractéristiques des séries dendrochronologiques de la pruche dans chacune des régions d’échantillonnage dendrochronologique………..117 Tableau 3.3. Cernes diagnostiques chez la pruche et le pin blanc dans les régions d’échantillonnage dendrochronologique……….118 Tableau 3.4. Caractéristiques des séries dendrochronologiques du pin blanc dans chacune des régions d’échantillonnage dendrochronologique………..120 Tableau 3.5. Statistiques comparatives des séries dendrochronologiques de pruche (A) et de pin blanc (B) entre les sites d’échantillonnage et pour la période commune à deux séries. Les résultats significatifs (p ≤ 0,001) sont en caractères gras………123 Tableau 3.6. Statistiques comparatives des séries dendrochronologiques de pruche et de pin
blanc au sein d’une même région d’échantillonnage et pour la période commune aux deux séries. Les résultats sont tous significatifs (p ≤ 0,001)………...124 Table 4.2. Number of false rings relative to their position within a growth ring and to the number of tracheids forming the intraannual band……….160 Table 4.3. Characteristics of the false ring chronologies derived from the rocky outcrop and
Table 4.3. Extreme radial growth data (widths of the narrowest and largest growth rings) for select populations of white pine (Pinus strobus), along with the mean yearly precipitation and temperature from Massachusetts (US), the Mauricie region, and the Outaouais Hills (Québec)………..169
Chapitre 1. Introduction générale
Les milieux forestiers, tels que nous les connaissons actuellement au Québec et, de manière générale, dans l’est de l’Amérique du Nord, sont le résultat de l’action combinée des perturbations naturelles et des activités humaines d’exploitation (Turner, 1989). Bien que la forêt y ait été très fréquentée et localement exploitée par les populations autochtones depuis longtemps (Litalien, 1993), les activités humaines qui ont eu un réel impact sur le couvert végétal ont débuté avec l’établissement permanent des Européens en Nouvelle-France, en particulier en bordure du fleuve Saint-Laurent et de ses affluents. Ces activités initiales ont mené à une déforestation qui a permis l’occupation des terres à des fins agricoles ou d’urbanisation (Marie-Victorin, 1935). C’est au début du 19e siècle, avec l’avènement de l’exploitation industrielle de la forêt, que l’impact de cette activité sur la dynamique de la végétation elle-même s’est fait sentir (Hardy et Séguin, 2004).
Jusqu'à son exploitation industrielle, la forêt mixte des régions tempérées du Québec a été essentiellement régie par des perturbations naturelles, d’étendue et d’intensité variables. Les épidémies d’insectes, les chablis et les feux sont des événements naturels récurrents qui peuvent constituer un facteur de mortalité des arbres au sein des peuplements affectés, dont l’ampleur varie beaucoup en fonction de l’intensité de la perturbation (Payette, 1992 ; Dansereau et Bergeron, 1993 ; Bergeron et Charron, 2010). Ces perturbations forestières et la succession qu’elles enclenchent ensuite contribuent au renouvellement des peuplements et entraînent un changement de la composition floristique des communautés végétales et de la répartition des individus ligneux dans les différentes classes d’âge (Lorimer, 1977 ; Pickett et White, 1985 ; Frelich et Lorimer, 1991 ; Payette, 2010 ; Frelich, 2002 ; Johnson et Miyanishi, 2007). Elles contribuent aussi au maintien de la diversité écologique à l’échelle du paysage (Mooney et Godron, 1983; Pickett et White, 1985 ; Talon et al., 2005).
Si la fréquence des feux en forêt boréale est assez bien connue, les intervalles variant de quelques décennies à plusieurs siècles (Bergeron et al., 2001 ; 2004 ; Talon et al., 2005 ; Bergeron et Charron, 2010), elle est cependant moins bien documentée en forêt tempérée.
Dans les régions où les forêts caducifoliées dominent, et malgré la présence de conifères comme le pin blanc (Pinus strobus L.) et la pruche (Tsuga canadensis (L.) Carr.), en peuplements purs ou mixtes (feuillus et conifères), le passage du feu est moins fréquent (Talon et al., 2005). Plus complexe à déterminer qu’en forêt boréale, la fréquence des feux de cime, entraînant plutôt un mode de renouvellement arbre par arbre, a toutefois été évaluée à 2700 ans dans les peuplements de pin gris (Pinus banksiana Lamb.) du Bas-Saint-Laurent (Parisien et al., 2004). En forêt tempérée, les feux peuvent certes constituer une perturbation sévère, mais leur fréquence reste faible.
Les perturbations dites secondaires, c'est-à-dire les épidémies d’insectes défoliateurs et les chablis, se produisent selon un intervalle de retour plus court que celui des feux (Lorimer, 1977 ; Boucher, 2008), mais elles sont généralement moins sévères (Pickett et White, 1985). En forêt tempérée, les épidémies d’insectes défoliateurs sont fréquentes, d’autant que les peuplements mixtes d’arbres feuillus et de conifères peuvent accueillir des insectes se rattachant tantôt à la forêt caducifoliée, tantôt à la forêt boréale. La livrée des forêts (Malacosoma disstria Hübner) est l’insecte phytophage ravageur des populations d’arbres feuillus de l’est du Canada le plus largement répandu en Amérique du Nord (Stehr and Cook 1968 ; Martineau, 1985 ; Frey et al., 2004 ; Cooke et al., 2011). Cet insecte est à l’origine du déclin des populations de peuplier faux-tremble (Populus tremuloides Michx.) et il constitue également le principal ravageur des populations de peuplier baumier (Populus balsamifera L.) en Alberta, en Ontario et au Québec (Hogg et al., 2002 ; Man and Rice, 2010). En période de pullulation, la livrée des forêts s’attaque à toutes les espèces d’arbres feuillus, sauf l’érable rouge (Acer rubrum L.) (Martineau, 1985). Les épidémies de cet insecte ont lieu environ tous les 10 ans (Frey et al., 2004 ; Moulinier et al., 2011). Elles sont d’une durée de 3 ans (à l’échelle du site) à 10 ans (à l’échelle du paysage) (Sippell, 1962 ; Witter, 1979 ; Cooke et al., 2009 ; Moulinier et al., 2011). Une défoliation de courte durée entraîne une diminution de la croissance radiale et la formation d’un cerne pâle chez les arbres défoliés (Frey et al., 2004), mais une défoliation soutenue, pendant au moins trois années consécutives, entraîne un accroissement du taux de mortalité dans les peuplements affectés (Churchill et al., 1964 ; Frey et al., 2004 ; Man and Rice, 2010 ; Moulinier et al. 2011). Cette mortalité crée des trouées dans le couvert forestier, qui sont à l’origine de la structure inéquienne des peuplements affectés par la livrée des forêts (Moulinier et al.,
2011). En forêt boréale, la tordeuse des bourgeons de l’épinette (TBÉ) (Choristoneura fumiferana (Clem.)) est le principal insecte ravageur des forêts de conifères (Martineau, 1985). Cet insecte défoliateur s’attaque principalement au sapin baumier (Abies balsamea (L.) Mill.) et à l’épinette blanche (Picea glauca (Moench) Voss). La défoliation peut causer une réduction importante de la croissance des arbres en période épidémique et entraîner une mortalité élevée au sein des peuplements affectés. Plus au sud, en forêt mixte, la TBÉ peut s’attaquer à environ 25 espèces de conifères, dont la pruche et toutes les espèces d’épinettes (Martineau, 1985). Au Québec, les épidémies de la TBÉ se produisent selon un intervalle de retour de 30 à 40 ans (Régnière et Lysyk, 1995 ; Jardon et al., 2003 ; Boulanger et Arsenault, 2004 ; Morin et al., 2010 ; Payette, 2010).
Les chablis résultent de la chute ou du déracinement d’un groupe d’arbres lors de vents violents ou d’ouragans (Peterson et Picket, 1995). Ce processus favorise un remaniement des sols, la disponibilité en nutriments et la régénération forestière, mais il augmente en revanche les conditions d’exposition des arbres qui restent sur pied ou se retrouvent en bordure des trouées ou des secteurs chablisés (Canham et Loucks, 1984; Payette et al., 1990). En forêt tempérée, la fréquence des chablis varie de quelques décennies (45 ans dans le cas de la réserve écologique de Tantaré) (Payette et al., 1990) à plus de 1000 ans (Canham et Loucks, 1984).
À ces perturbations naturelles, s’est ajoutée, au cours du 19e siècle, l’exploitation industrielle des forêts, d’abord dans la vallée du Saint-Laurent, puis au sein des bassins versants de ses principaux tributaires, en particulier ceux de la rivière des Outaouais et du Saint-Maurice. C’est en effet au début du 19e siècle que l’Angleterre, contrainte par le conflit qui l’opposait à la France, alors sous l’autorité de Napoléon, entreprit l’exploitation de ses forêts coloniales au Québec (Lower, 1938 ; Gaudreau, 1988 ; 1999). Dès lors, l’exploitation forestière va entraîner de profonds changements écologiques, tant dans les milieux forestiers que dans les cours d’eau et les bassins hydrographiques. On note, par exemple, un certain contrôle des feux dans les aires exploitées (Friedman et Reich, 2005 ; Bergeron et al., 2006), le prélèvement sélectif d’une grande quantité d’individus matures au sein des peuplements forestiers ou encore l’aménagement des cours d’eau pour le flottage du bois.
Le transport par flottage des billes de bois, opération connue sous l’appellation de drave, est un symbole de l’exploitation forestière au Québec (Hardy et Seguin, 2004). Il s’agit d’une activité qui a été très importante pendant plus de 150 ans. Dans les régions exploitées, une grande partie du réseau hydrographique a été aménagé dans le but de faciliter le transport du bois (Lafleur, 1970 ; Pelletier-Bellefleur, 1974 ; Hardy et Séguin, 2004). Ce mode de transport a laissé des traces, surtout le long des cours d’eau qui ont servi à cette fin. En plus des infrastructures de bois érigées sur les cours d’eau (barrages de retenue des eaux et du bois), une grande quantité de billes de bois de drave a été abandonnée sur place et a coulé par la suite, tapissant la zone infralittorale de débris ligneux. La présence de ces billes a entraîné des changements importants au sein de ces écosystèmes aquatiques, comme la modification du bilan sédimentaire et des propriétés physico-chimiques de l’eau, ainsi que la dégradation des frayères (Plante, 2006).
À ces modifications du réseau hydrographique, viennent s’ajouter celles qui sont en lien direct avec l’exploitation du couvert forestier. D’abord locale et de type artisanal, l’exploitation forestière a connu de grandes phases au cours desquelles l’intensité des pratiques de coupe s’est accrue au fil du temps. Au début du 20e siècle, la récolte va prendre une ampleur telle que son influence sur le milieu forestier deviendra comparable, voire supérieure, à celle d’une perturbation naturelle sévère et de grande étendue (Carleton et MacLellan, 1994 ; Dussart et Payette, 2002). Les prélèvements massifs de bois ont entraîné une modification de la composition en espèces des peuplements et leur rajeunissement (Whitney, 1994 ; Bouchard et Domon, 1997 ; Friedman et Reich, 2005).
Ces changements brusques au sein des peuplements de la période préindustrielle se sont traduits, entre autres, par une quasi disparition des peuplements mono- ou bispécifiques de pruche et de pin blanc (Hardy et Séguin, 1984 ; 2004 ; Mladenoff et al., 1993 ; Foster et al., 1998). Ces deux espèces, parmi les plus longévives de ces régions avec le thuya occidental (Thuya occidentalis L.) (Arseneault, 2010), étaient bien représentées dans les forêts de la période préindustrielle dans le sud du Québec. Les billes de bois submergées constituent donc une source de données paléoenvironnementales de première importance, qui peuvent être mises en valeur grâce à une analyse dendroécologique.
Associée à l’écologie végétale, la dendroécologie aborde l’étude des relations spatiotemporelles entre les végétaux ligneux et leur habitat (Payette et Filion, 2010). Axée sur la notion de temps, elle met en valeur la capacité des plantes ligneuses à enregistrer des données qui permettent de mieux comprendre leur environnement (Payette, 2010). En ce sens, elle se rattache au domaine de l’écologie historique (McIntosh, 1985). Les méthodes dendroécologiques sont de plus en plus utilisées dans l’interprétation de la structure et du fonctionnement des peuplements forestiers. Elles permettent de reconstituer les régimes de perturbations à l’aide d’arbres vivants et d’arbres morts, et de statuer sur la dynamique passée, actuelle et à venir des forêts. L’utilisation d’arbres vivants permet de couvrir des périodes qui vont de quelques décennies à plusieurs centaines d’années, selon la longévité des arbres en présence. Toutefois, les vieux arbres vivants sont relativement rares et leur analyse ne permet pas de couvrir des périodes très longues. L’utilisation concomitante d’arbres morts, en revanche, permet des reconstitutions écologiques pour des périodes plus anciennes, qui ne sont habituellement que peu ou pas documentées dans les écrits.
De nombreux documents historiques font état des activités de coupe en forêt et du mode de transport du bois par la drave. On trouve des sources primaires comme les carnets d’arpentage qui décrivent l’état des peuplements avant la coupe (Mauri Ortuno, 2010), les actes notariés relatifs aux ventes de bois (Simard et Bouchard, 1996), les archives de certaines compagnies forestières qui font état de leurs activités commerciales (CIEQ), ou encore des photographies historiques (CIEQ, 2012 ; Musée McCord, 2012). Il existe également de nombreuses sources secondaires qui sont souvent le fruit de travaux de recherche et de synthèse d’envergure et qui prennent la forme de dossiers statistiques ou de monographies régionales (Hardy et Séguin 1980 ; 2004).
Le Parc national de la Mauricie (PNM) est une aire protégée qui vise la sauvegarde de territoires qui ont été le théâtre de ces activités passées d’exploitation forestière. Créé en 1971, le PNM occupe une superficie de 536,7 km2. Il est situé à la limite méridionale du
Bouclier canadien, dans une zone de transition où plusieurs espèces atteignent leur limite septentrionale ou méridionale de répartition (Latourelle et Blouin, 2010). Dans le cadre d’un projet de restauration de l’intégrité écologique des lacs, les autorités du parc ont entrepris l’extraction de la majeure partie des billes de bois de flottage qui ont coulé dans la
zone infralittorale de certains lacs (Plante, 2006). Si la première conséquence de ce programme fut la restauration des habitats aquatiques, la seconde fut le prélèvement de plus de 17000 billes de bois de 14 pieds (4,20 m) dans cinq lacs. Les autorités du parc souhaitaient la mise en valeur de cette importante source de données environnementales et historiques. L’analyse dendrochronologique préliminaire de quelques dizaines de billes, la grande majorité étant de la pruche, laissait entrevoir un grand potentiel de reconstitution dendroécologique et historique en raison de leur grand âge.
Ce type de matériel s’apparente aux mégafossiles ligneux décrits par Arseneault et Filion (2001). Les mégafossiles comprennent des troncs d’arbres, mais aussi certaines parties comme les souches ou les branches de grande dimension. Le bois peut se conserver pendant de longues périodes dans divers types de milieux terrestres, semi-aquatiques ou aquatiques, là où les conditions demeurent anoxiques. En milieu terrestre, le matériel minéral détritique, par exemple alluvionnaire, éboulisé ou déposé à la suite de mouvements de masse (ex. glissements de terrain) constitue un milieu privilégié de conservation des bois fossiles (Becker, 1993 ; Freidrich, 2004). Les tourbières et les lacs sont aussi des milieux où les mégafossiles ligneux peuvent être très abondants (Arseneault et Payette, 1998). Des troncs d’arbres enfouis et très bien conservés ont permis une reconstitution des conditions paléoenvironnementales pour des périodes souvent très anciennes. Par exemple, les troncs extraits de la tourbe dans les dunes paludifiées du complexe éolien de Saint-Flavien dans la région de Lotbinière, ainsi que les macrorestes de plantes et d’insectes, ont permis d’identifier les causes du déclin de la pruche à l’Holocène moyen (Bhiry et Filion, 1996). L’abondance de troncs de pruche, de pin blanc et de mélèze laricin (Larix laricina (Du Roi) K.Koch) y a aussi permis une reconstitution paléoenvironnementale de plusieurs millénaires en lien avec la paludification de ces vastes surfaces sableuses, en réponse aux changements hydroclimatiques survenus au cours de l’Holocène moyen et supérieur. De façon analogue, mais dans le Nord du Québec, Arseneault et Payette (1998) ont prélevé plus de 1300 échantillons d’épinette noire (Picea mariana [Mill.] BSP.), dans les tourbières à pergélisol situées le long du cours moyen de la rivière Boniface. Cette étude a permis de prolonger de plus de 600 ans la chronologie des cernes pâles chez l’épinette noire et de documenter le rôle respectif des conditions écologiques locales et du climat sur la formation de ce type particulier de cernes diagnostiques.
De nombreuses études ont permis l’analyse de troncs d’arbres qui ont été transportés à la surface de l’eau comme le bois de dérive (driftwood) qui peut se conserver très longtemps sur les berges, en particulier dans les conditions froides de l’Arctique (Eggertsson, 1994), ou encore le bois de flottage qui peut aussi persister longtemps au fond des lacs et des rivières (Eggertsson, 1994 ; Douglas et al., 2006 ; Boucher et al., 2009). Grabner et al. (2004) ont ainsi étudié les infrastructures qui ont servi à la drave des conifères pendant la période industrielle, depuis les hautes montagnes alpines sur les rivières Enns et Steyer en Autriche. Dans le sud-ouest de l’Allemagne, l’étude des bois archéologiques de milieu humide (bord de lacs et de tourbières) a permis, en plus de la datation des habitations palafittiques, une reconstitution paléoécologique des milieux d’origine de ces bois (Billlamboz, 2010). En retraçant les phases de l’occupation du sol entre -4000 et -2400 ans avant aujourd'hui dans les environs du lac de Constance, Billlamboz (2010) a effectué une reconstitution du cadre de vie des sociétés néolithiques, tout en faisant ressortir le rôle de l’Homme dans l’évolution de son environnement.
Les bois fossiles permettent de faire le lien entre les études dendroécologiques des bois modernes et certaines méthodes de la paléoécologie (palynologie, anthracologie, analyse macrofossile) utilisées pour documenter des périodes anciennes, mais qui sont peu utiles dans l’étude de phénomènes récents. À ce titre, les billes de bois de flottage extraites des lacs du PNM constituent une source inestimable de données primaires (sensu Courville, 1995) qui permet de compléter les connaissances que nous avons de l’histoire de la forêt et de son exploitation. Dans ce travail, la dendroécologie entend favoriser la convergence des méthodes historiques, géographiques et écologiques afin de produire des données nouvelles qui permettront de mieux comprendre les activités d’exploitation des forêts en Mauricie, ainsi que certaines perturbations écologiques comme les épidémies d’insectes défoliateurs.
Cette recherche a pour but de montrer quels ont été les impacts de ces diverses perturbations sur la croissance radiale des arbres en exploitant le matériel dendrochronologique mis à notre disposition : les nombreuses billes de bois de flottage extraites de quelques lacs du PNM et quelques chronologies de pruche et de pin blanc d’autres régions (parc de la Gatineau, réserve écologique de la Rivière-du-Moulin dans Lotbinière et île d’Anticosti) qui sont utilisées ici à des fins de comparaison interrégionale.
De l’ensemble de ce matériel, nous pouvions escompter des données environnementales de grande valeur et ce, pour plusieurs raisons. En Mauricie, le nombre élevé de billes de bois et leur âge avancé (tel qu’évalué par le dénombrement des cernes annuels) ont permis d’envisager la construction de longues séries de pruche et de pin blanc dans une région où il est difficile de trouver de vieux arbres vivants. De plus, les échantillons de bois montraient de nombreuses séquences de cernes étroits suivies d’une récupération plus ou moins rapide. Il s’agit d’un patron de croissance analogue à celui que présentent les arbres touchés par une défoliation sporadique (Krause et Morin, 1999), ce qui laissait entrevoir un grand potentiel d’analyse dendroécologique. Enfin, la disponibilité de séries dendrochronologiques des mêmes espèces, mais provenant d’autres régions, a permis d’effectuer des comparaisons interrégionales en ce qui a trait à la croissance de ces conifères tempérés à la limite de leur aire de répartition.
D’abord axée sur l’historique de l’exploitation forestière en Mauricie, notre recherche s’est donc étendue à l’analyse du patron de croissance chez la pruche, un patron jugé a priori caractéristique de la défoliation par des insectes phytophages. L’épinette rouge (Picea rubens Sarg) a aussi été analysée car nous disposions d’une série dans le PNM. S’ouvrait ainsi à nous le vaste champ d’investigation de cette importante perturbation forestière.
Nous avons formulé l’hypothèse que le patron de croissance des pruches en Mauricie répondait à des perturbations multiples (exploitation forestière et épidémies d’insectes défoliateurs), avec en toile de fond des conditions climatiques qui ont pu changer au fil des siècles et qui diffèrent entre les régions d’échantillonnage dendrochronologique. Un des principaux défis à relever dans cette thèse résidait donc dans l’utilisation de méthodes d’analyse appropriées en vue d’une interprétation crédible du patron de croissance radiale des arbres dans un contexte de perturbations multiples.
L’objectif général de cette thèse est d’étudier la croissance radiale de la pruche et du pin blanc dans la forêt tempérée du Québec à l’aide des méthodes dendroécologiques, afin de comprendre les relations entre le patron de croissance et les perturbations forestières, d’une part, et le climat, d’autre part.
Les objectifs spécifiques sont les suivants :
1) Effectuer une analyse dendroécologique de la pruche et du pin blanc en Mauricie en lien avec les principales perturbations, à savoir l’exploitation historique des forêts et les épidémies d’insectes défoliateurs ;
2) Effectuer une analyse dendroclimatique des séries de la Mauricie et celles d’autres régions du sud (Outaouais, Lotbinière) et de l’est (Anticosti) du Québec, afin d’évaluer les possibilités d’extraire le signal climatique chez des arbres ayant subi ces perturbations ;
3) Analyser un cas particulier de cernes diagnostiques observé chez le pin blanc dans les collines de l’Outaouais, à savoir la présence généralisée de faux cernes qui sont des bandes intraannuelles de bois de densité plus élevée que le bois adjacent.
Après cette introduction générale (Chapitre 1), le chapitre 2, qui s’intitule « Analyse dendroécologique de la pruche (Tsuga canadensis) et du pin blanc (Pinus strobus) en relation avec la coupe forestière et les épidémies d’insectes défoliateurs dans le Parc national de la Mauricie (Québec, Canada) » présente, en plus des longues séries dendrochronologiques, les données concernant l’exploitation de la forêt et la reconstitution historique des épidémies d’insectes défoliateurs, nommément de la tordeuse des bourgeons de l’épinette. Ce chapitre met également en relation les perturbations naturelles et anthropiques qui se conjuguent dès le milieu du 19e siècle.
Le chapitre 3, intitulé « Analyse dendroclimatique chez le pin blanc et la pruche dans la forêt tempérée du Québec, dans un contexte de perturbations multiples » utilise les méthodes classiques de la dendroclimatologie que sont les fonctions de réponse et de corrélation. Les analyses portent sur les séries dendrochronologiques présentées dans le chapitre 2, mais uniquement sur la partie des séries élaborée à l’aide d’arbres vivants, ainsi que sur plusieurs autres séries des mêmes espèces. Il a permis d’évaluer la qualité de la relation cernes de croissance/climat dans un contexte de perturbations multiples, dans des régions climatiquement contrastées du sud-ouest (Outaouais), du centre-sud (Mauricie et Lotbinière) et de l’est du Québec (île d’Anticosti).
Le chapitre 4, intitulé « False rings in the white pine (Pinus strobus) of the Outaouais Hills, Québec (Canada), as indicators of water stress » porte sur une analyse des faux cernes chez le pin blanc dans une région du Québec où peuvent sévir des épisodes momentanés de sécheresse. Il a permis d’identifier les facteurs qui prédisposent et déclenchent la formation de faux cernes, ainsi que les seuils de précipitation qui entraînent la formation de ces bandes intraannuelles de bois plus dense. Ce chapitre est publié dans le numéro de janvier 2012 de la Revue Canadienne de Recherche Forestière.
Enfin, la conclusion générale (chapitre 5) résume les principales contributions de cette thèse et identifient certaines avenues de recherche qui pourraient être explorées.
Références
Arseneault, D. 2010. Dendrochronologie dans le nord-est de l'Amérique du nord. In Payette, S., et Filion, L. 2010. La Dendroécologie : Principes, méthodes et applications. Presses de l’Université Laval, Québec, Québec. pp. 731-747.
Arseneault, D. et Filion, L. 2001. Les mégafossiles ligneux. In Payette S., et Rochefort L. Écologie des tourbières du Québec-Labrador, Presses de l’Université Laval, Québec, Québec pp. 327-341.
Arseneault, D., et Payette, S. 1998. Chronologie des cernes pâles de l’épinette noire (Picea mariana [Mill.] BSP.) au Québec subarctique : de 706 à 1675 ap. J.-C. Géographie Physique et Quaternaire 52(2): 1-8.
Becker, B. 1993. An 11,000-year german oak and pine dendrochronology for radiocarbon calibration. Radiocarbon 35(1): 201-213.
Bergeron Y., et Charron, D., 2010. Dendrochronologie appliquée à l’aménagement forestier écosystémique : le cas de la forêt du Lac Duparquet, Québec. In Payette, S., et Filion, L. 2010. La Dendroécologie : Principes, méthodes et applications. Presses de l’Université Laval, Québec, Québec. pp. 415-436.
Bergeron, Y., Cyr, D., Drever, C.R., Flannigan, M., Gauthier, S., Kneeshaw, D., Lauzon, A, Leduc, A., Goff, H.L., Lesieur, D., et Logan, K. 2006. Past, current, and future fire frequencies in Quebec's commercial forests: implications for the cumulative effects of harvesting and fire on age-class structure and natural disturbance-based management. Canadian Journal of Forest Research 36(11): 2737-2744.
Bergeron, Y., Gauthier, S., Flannigan, M., et Kafka, V. 2004. Fire regimes at the transition between mixed wood and coniferous boreal forest in Northwestern Quebec. Ecology 85(7): 1916-1932.
Bergeron, Y., Gauthier, S., Kafka, V., Lefort, P., et Lesieur, D. 2001. Natural fire frequency for the eastern Canadian boreal forest: consequences for sustainable forestry. Canadian Journal of Forest Research 31(3): 384-391.
Bhiry, N., et Filion, L. 1996 Mid-Holocene hemlock decline in eastern North America linked with phytophagous insect activity Quaternary Research 45(3): 312-320. Billamboz, A.2010 Dendroarchéologie et écologie des palafittes In Payette, S., et Filion, L.
2010. La Dendroécologie : Principes, méthodes et applications. Presses de l’Université Laval, Québec, Québec. pp.509-536.
Bouchard, A., et Domon, G. 1997. The transformations of the natural landscapes of the Haut-Saint-Laurent (Québec) and their implications on future resource management. Landscape and Urban Planning 37(1-2): 99-107.
Boucher, Y. 2008. Dynamique de la forêt du Bas-St-Laurent depuis le début de l’exploitation forestière (1820-2000). Thèse de doctorat, Département des sciences de l’environnement. Université du Québec à Rimouski, Rimouski.124p.
Boucher, Y., Arseneault, D., et Sirois, L. 2009. Logging history (1820-2000) of a heavily exploited southern boreal forest landscape: Insights from sunken logs and forestry maps. Forest Ecology and Management 258(7): 1359-1368.
Boulanger, Y., et Arseneault, D. 2004. Spruce budworm outbreaks in eastern Quebec over the last 450 years. Canadian Journal of Forest Research 34(5): 1035-1043.
Canham, C.D., et Loucks, O.L. 1984. Catastrophic windthrow in the presettlement forests of Wisconsin. Ecology 65(3): 803-809.
Carleton, T.J., et Maclellan, P. 1994. Woody vegetation responses to fire versus clear-cutting logging: A comparative survey in the central Canadian boreal forest. Ecoscience 1(2): 141-152.
Churchill, G.B., John, H.H., Duncan, D.P., et Hodson, A.C. 1964. Long-Term Effects of Defoliation of Aspen by the Forest Tent Caterpillar. Ecology 45(3): 630-636. CIEQ. 2012. Base de donnée en histoire régionale, Mauricie, http://mauricie.cieq.ca/
Cooke, B.J., Lorenzetti, F., et Roland, J. 2009. On the duration and distribution of forest tent caterpillar outbreaks in east-central Canada. Journal of the Entomological Society of Ontario 140: 3-18.
Cooke, B.J., MacQuarrie, C.J.K., et Lorenzetti, F. 2011. The dynamics of forest tent caterpillar outbreaks across east-central Canada. Ecography 35(5): 422-435.
Courville. 1995. Introduction à la géographie historique. Presses de l'Université Laval, Sainte-Foy, Québec.
Dansereau, P.R., et Bergeron, Y. 1993. Fire history in the southern boreal forest of northwestern Quebec. Canadian Journal of Forest Research 23(1): 25-32.
Douglas H., R., Panyuyshkina I. P., Leavitt, S. W. Wiedenhoeft, A. C, et Zawiskie, J. 2006. A multiproxy environmental investigation of Holocene wood from a submerged conifer forest in Lake Huron, USA. Quaternary Research. 66(1): 67-77.
Dussart, E., et Payette, S. 2002. Ecological impact of clear-cutting on black spruce-moss forests in southern Quebec. Ecoscience 9(4): 533-543.
Eggertsson, O. 1994. Mackenzie River driftwood - A dendrochronological study. Arctic 47 (2): 128-140.
Foster, D.R., Motzkin, G., et Slater, B. 1998. Land-use history as long-term broad-scale disturbance: Regional forest dynamics in central New England. Ecosystems 1(1): 96-119.
Freidrich, M., Remmelel, S., Kromer, B., Hofmann, J., Spurk, M., Kaiser, K.F., Orcel, C., et Kuppers, M. 2004. The 12,460-year Hohenheim oak and pine tree-ring
chronology from central Europe - A unique annual record for radiocarbon
calibration and paleoenvironment reconstructions. Radiocarbon 46(3): 1111-1122. Frelich, L.E. 2002. Forest dynamics and disturbance regimes: studies from temperate
evergreen-deciduous forests. Cambridge University Press, New York.
Frelich, L.E., et Lorimer, C.G. 1991. Natural Disturbance Regimes in Hemlock-Hardwood Forests of the Upper Great Lakes Region. Ecological Monographs 61(2): 145-164. Frey, B.R., Lieffers, V.J., Hogg, E.H., et Landhausser, S.M. 2004. Predicting landscape
patterns of aspen dieback: Mechanisms and knowledge gaps. Canadian Journal of Forest Research-Revue Canadienne De Recherche Forestiere 34(7): 1379-1390.
Friedman, S.K., et Reich, P.B. 2005. Regional legacies of logging: Departure from presettlement forest conditions in northern Minnesota. Ecological Applications 15(2): 726-744.
Gaudreau, G. 1988. L'exploitation des forêts publiques au Québec (1874-1905): transition et nouvel essor. Revue d'histoire de l'Amérique française 42(1): 3-27.
Gaudreau, G. 1999. Les récoltes des forêts publiques au Québec et en Ontario, 1840-1900, McGill-Queen’s University Press. Montréal & Kingston.
Grabner, M., Wimmer, R., et Weichenberger, J. 2004. Reconstructing the history of log-drifting in the Reichraminger Hintergebirge, Austria. Dendrochronologia 21(3): 131-137.
Hardy, R., et Seguin, N. 1984. Forêt et société en Mauricie, 1850-1930. Bulletin d'histoire de la culture matérielle 13: 51-57.
Hardy, R., et Seguin, N. 2004. Histoire de la Mauricie. Institut Québécois de recherche sur la culture, Sainte-Foy.
Hogg, E.H., Brandt, J.P., et Kochtubajda, B. 2002. Growth and dieback of Aspen forests in northwestern Alberta, Canada, in relation to climate and insects. Canadian Journal of Forest Research-Revue Canadienne De Recherche Forestiere 32(5): 823-832. Jardon, Y., Morin, H., et Dutilleul, P. 2003. Périodicité et synchronisme des épidémies de
la tordeuse des bourgeons de l’épinette au Québec. Revue canadienne de recherche forestière 33(10): 1947-1961.
Johnson E.A., et Miyanishi, K. 2007 Plant Disturbance Ecology: the Process and the Response. Elsevier, Academic Press, Amsterdam.
Krause, C., et Morin, H. 1999. Tree-ring patterns in stems and root systems of black spruce (Picea mariana) caused by spruce budworms. Canadian Journal of Forest Research 29(10): 1583-1591.
Lafleur, N. 1970. La Drave en Mauricie, des origines à nos jours : histoire et traditions. Éditions du Bien public, Trois-Rivières, Québec.
Latourelle, A., et Blouin, T. 2010. Plan directeur, Parc national du Canada de la Mauricie. Litalien, R. 1993. Les explorateurs de l’Amérique du Nord 1492-1795. Septentrion,
Québec.
Lorimer, C.G. 1977. The Presettlement Forest and Natural Disturbance Cycle of Northeastern Maine. Ecology 58(1): 139-148.
Lower, A.R.M. 1938. The North American assault on the Canadian forest: A history of the lumber trade between Canada and the United States. The Ryerson Press, Toronto. Man, R.Z., et Rice, J.A. 2010. Response of aspen stands to forest tent caterpillar defoliation
and subsequent overstory mortality in northeastern Ontario, Canada. Forest Ecology and Management 260(10): 1853-1860.
Marie-Victorin, Fr. 1935 Flore Laurentienne. Troisième édition (2002), Presses de l’Université de Montréal, Montréal, Québec.
Martineau, R. 1985. Insectes nuisibles des forêts de l'est du Canada, Service canadien des forêts : Centre d'édition du gouvernement du Canada Québec.
Mauri Ortuno, E. 2010. Modélisation de la distribution précoloniale du pin blanc en
Moyenne-Mauricie à partir des carnets d'arpentage. Mémoire de maîtrise, Faculté de Foresterie, deGéographie et de Géomatique, Département des sciences du bois et de la forêt, Université Laval, Québec.
McIntosh, P.R. 1985. The background of ecology, concept and theory. Cambridge University Press, Cambridge.
Mladenoff, D.J., White, M.A., Pastor, J., et Crow, T.R. 1993. Comparing spatial pattern in unaltered old-growth and disturbed forest landscapes. Ecological Applications 3(2): 294-306.
Mooney, H.A., et Godron, M. 1983. Disturbance and ecosystems: Components of response. Springer-Verlag.
Morin, H., Jardon, Y., et Simard, S. 2010. Détection et reconstitution des épidémies de la tordeuse des bourgeons de l’épinette (Christoneura fumiferana) à l’aide de la dendroécologie. In Payette, S., et Filion, L. 2010. La Dendroécologie : Principes, méthodes et applications. Presses de l’Université Laval, Québec, Québec. pp. 415-436.
Moulinier, J., Lorenzetti, F., et Bergeron, Y. 2011. Gap dynamics in aspen stands of the Clay Belt of northwestern Quebec following a forest tent caterpillar outbreak. Canadian Journal of Forest Research 41(8): 1606-1617.
Musée McCord 2012. Musée MacCord, notre monde, nos histoires. http://www.musee-mccord.qc.ca/fr/clefs/collections/.
Parisien, M.A., Sirois, L. et Babeau, M. 2004. Distribution and dynamics of jack pine at its longitudinal range limits in Québec. In Engstrom, R.T., Galley K.E.M., and De Groot W.J., (eds.). Proceedings of the 22nd, and Conference: T.T.F.E. 2001.