HAL Id: tel-01937224
https://tel.archives-ouvertes.fr/tel-01937224
Submitted on 28 Nov 2018
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of
sci-entific research documents, whether they are
pub-lished or not. The documents may come from
teaching and research institutions in France or
abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est
destinée au dépôt et à la diffusion de documents
scientifiques de niveau recherche, publiés ou non,
émanant des établissements d’enseignement et de
recherche français ou étrangers, des laboratoires
publics ou privés.
Régulations chromatiniennes et transcriptionnelles
impliquées dans le cycle de vie du puceron du pois
Gautier Richard
To cite this version:
Gautier Richard. Régulations chromatiniennes et transcriptionnelles impliquées dans le cycle de vie du
puceron du pois. Génétique des plantes. Agrocampus Ouest, 2017. Français. �NNT : 2017NSARB130�.
�tel-01937224�
Thèse AGROCAMPUS OUEST sous le label de l’Université Bretagne Loire pour obtenir le grade de DOCTEUR D’AGROCAMPUS OUEST Spécialité Génétique, génomique et bio-informatique
Gautier RICHARD
•
20 octobre 2017
•
ÉCOLE DOCTORALE • Écologie, Géosciences, Agronomie et Alimentation (EGAAL)
LABORATOIRE D’ACCUEIL • INRA - Institut de génétique, environnement et protection des plantes (UMR IGEPP)
Régulations chromatiniennes et
transcriptionnelles impliquées
dans le cycle de vie
du puceron du pois
Maria MANZANARES-DAULEUX
Professeure, AGROCAMPUS OUEST, UMR INRA - AO - UR1 IGEPP / présidente
Sébastien BLOYER
Professeur, Université Paris Sud / rapporteur Nadia PONTS
Chargée de recherche, INRA Bordeaux Aquitaine/ rapporteur Emmanuelle JACQUIN-JOLY
Directrice de recherche, INRA Versailles Grignon / examinatrice Stéphanie JAUBERT-POSSAMAI
Chargée de recherche, INRA PACA / examinatrice Denis TAGU
Directeur de recherche, UMR INRA - AO - UR1 IGEPP / directeur de thèse
Gaël LE TRIONNAIRE
Chargée de recherche, UMR INRA - AO - UR1 IGEPP / co-encadrant
the males’ single X in somatic cells. On the other hand, specifi c seem to participate in the defi nition of their reproduction mode the fi xation of transcription factors. Their analysis shows the
informatiques de données d’expression des gènes (RNA-seq) et
d’acces-fi ques d’ouverture de la chromatine entre embryons sexués ou asexués semblent participer à la défi nition de leur mode de reproduction en modu-lant l’expression de certains gènes et en permettant la fi xation de
Thèse de Doctorat AGROCAMPUS OUEST pour obtenir le titre de DOCTEUR d’AGROCAMPUS OUEST Spécialité Biologie et Agronomie
École Doctorale : Vie-Agro-Santé
Laboratoire d’accueil : UMR Institut de Génétique, Environnement et Protection des Plantes (IGEPP, INRA, Agrocampus Ouest, Université Rennes 1), Ecologie et Génétique des Insectes (EGI)
Régulations chromatiniennes et transcriptionnelles
impliquées dans le cycle de vie du puceron du pois
Gautier RICHARD • 20 Octobre 2017
Présentée devant la commission d’examen composée de :
Présidente : Maria Manzanares Rapportrice : Nadia Ponts Rapporteur : Sébastien Bloyer
Examinatrice : Emmanuelle Jacquin-Joly Examinatrice : Stéphanie Jaubert-Possamai Directeur de thèse : Denis Tagu
Maître de thèse : Gaël Le Trionnaire
Professeure, Agrocampus Ouest, UMR IGEPP, Rennes Chargée de Recherche, INRA, UR MycSA, Bordeaux Professeur, Université Paris-Sud, UMR I2BC CNRS, Paris Directrice de recherche, INRA, UMR iEES, Versailles Chargée de recherche, INRA, UMR ISA, Sophia Antipolis Directeur de Recherche, INRA, UMR IGEPP, Rennes Chargé de Recherche, INRA, UMR IGEPP, Rennes
0. Table des matières 0.
Remerciements
Je tiens tout d’abord à remercier l’ensemble des membres du jury et mes deux rapporteurs, Nadia Ponts et Sébastien Bloyer, ainsi que l’ensemble des membres de mon comité de thèse pour le soutien qu’ils m’ont apporté durant mes 3 années de thèse.
Bien évidemment, j’adresse un immense merci à mes encadrants, Denis Tagu et surtout Gaël Le Trionnaire qui a su me supporter, me motiver dans les coups durs, et me transmettre son énergie, sa passion de la science et bien plus encore durant ces 3 ans et demi (il ne faut pas oublier le stage de M2 !).
Merci à mes trois techniciennes préférées, Sylvie T., Nathalie et Sylvie H., et merci à mes deux bioinformaticiens préférés, Fabrice et Anthony, sans qui cette thèse n’aurait pas pu être possible. Un immense merci à mes parents, Corinne et Olivier, et à mon frère Thibaud, sans eux et l’amour qu’ils m’ont apporté, je ne serais pas là où j’en suis actuellement. Merci à mes beaux-parents Daniel et Dominique et bien évidemment à ma chérie d’amour, Laure, pout tout le soutient et l’amour qu’ils ont su m’apporter durant ma thèse et l’ensemble de mes études.
Un immense merci à tous les collègues et amis de l’INRA : Benoît (tu me manqueras mon pote, mais on est jamais très loin t’inquiète ! Toujours chaud pour une partie de babyfoot !), Lydie, Yoann A., Yoann B., Julien, JB, Camille, David, Marion, Paola, Morgane H., Thibaut et Thibault (les 3 orthographes dans la même page !).
Un grand merci à tous mes potes Rennais pour tous ces moments de rigolade et d’amitié : David, Morgane R., Florent, Étienne, FX, Geoffroy, Thomas, Heathcliff, Axel et Ludo.
Un bisou sur la truffe et une caresse aux différents animaux de compagnie de la famille : Tessa, Moumousse (vous nous manquerez à tous), Lilou, et plus récemment Odin (pour les titounets), Choupi et Pompon (pour les chats !).
Merci à mes sponsors : Spotify, Discord, Steam, Bonzini et René Pierre. Merci à Risitas et son inspiration quotidienne, et la chancla.
0. T ab le d es m atières 0.
Tab le des m atières
I. INT RO D U CT IO N GEN ERAL E ... 4
II. SYNT HES ES BIBLIO GRAP H IQ UES ... 8
A . .Lh [ h DL9 5 9S t U / 9wh b S... 8
1. Les pucerons : des ravageurs m ajeurs des cultures agronom iques ... 8
2. Cycle de vie e t alternance du m ode de reproduction ...10
3. Traits d’histoire de vie et plasticité phénotypique ...13
4. Le chrom osom e X chez les p ucerons : cytologie et com pensation ...20
5. B ases m oléculaires du polyphénism e de reproduction ...21
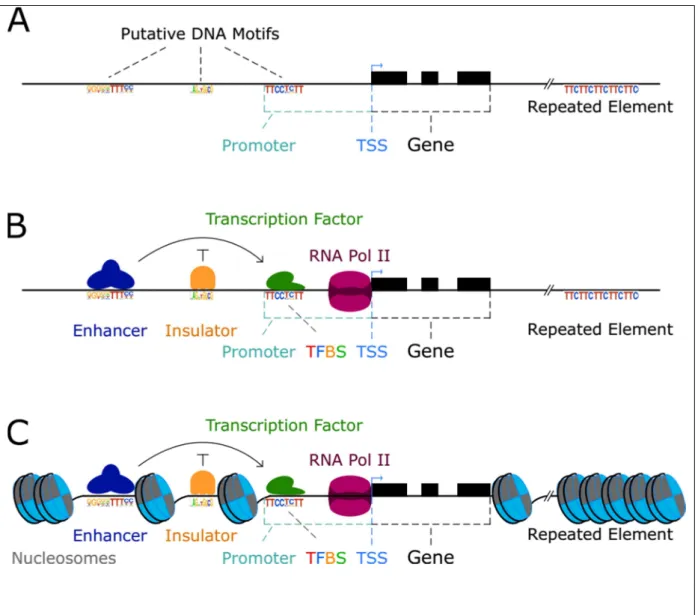
. . a9/ A b LS a 9S 9t LD9b 9TLv U 9S 9T S TwU / TU w9 5 9 [ A / H wh a A TLb 9... 25 1.Unités de transcriptions ...25
2.Chrom atine et régions régula trices longue distance ...27
3.Accessibilité de la chrom atine ...31
4.M odifications post-traductionnelles des histones ...35
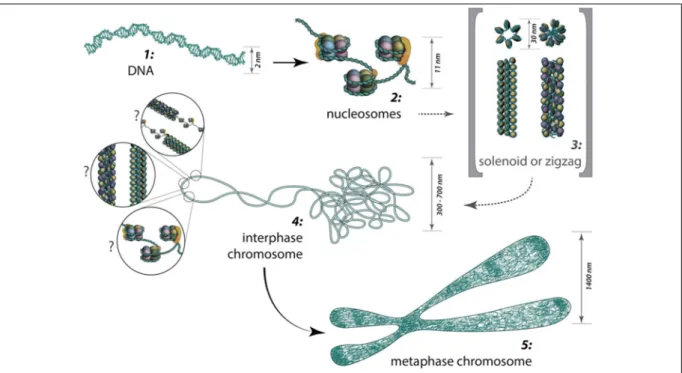
5.Structure tridim ensionnelle de la chrom atine : définition et m éthodes d’études ...40
6.M éthylation de l’AD N ...42
7.ARN non-codan ts ...45
8.Epigénétique chez les pucerons ...49
/ . /H wh a h S h a 9 X 9T / h a t 9b S A TLh b 5 9 5 h S 9... 50 1.D éfinition de la com pensation de dose ...50
2.Les m écanism es épigénétiques participant à la com pensation de dose chez les organis m es m odèles ...52
3.Étude de la com pensation de dose chez les insectes n on-d rosophilides ...56
5 . t[ A S TL/ LT9 t H 9b h TYt Lv U 9, t h [ Yt H 9b LS a 9S 9T a 9/ A b LS a 9S 9t LD9b 9TLv U 9S / H 9Z [ 9S Lb S 9/ T9S... 57 1.Le polyphénism e de ca stes chez l’abeille ...57
2.Le polyphénism e de ca stes chez les fourm is ...59
3.Le polyphénism e de phase chez le criquet ...60
0. T ab le d es m atières 0.
IV. MAT ERIEL ET M ET HO D ES ... 64
A . A/ v U LS LTLh b 5 U a A T9wL9[ . Lh [ h DLv U 9... 64 1.Com pensation de dose ... ErrorA Bookm ark not defined. 2.Polyphénism e de reproduction ...65
. . Ch wa A [ 5 9H Y5 9 AS S LS T95 LS h [ A TLh b h C w9DU [ A Th wY9[ 9a 9b TS S U L± L 5’U b S 9v U 9b 4A D9 H A U T-5 9. LT (CA Lw9-S 9v).. 68 1.Protocole u tilisé ...68 2.M éthodes d’analyse bioinform atique de données FAIRE-s eq ...70
V. CHAPIT RE I : CO MPENSAT IO N D E D O S E ET ACCESS IB ILIT E SEXE-SPE CIF IQ UE D U
CH RO M O S O M E X CH EZ A CYRTHO SIP HO N PISUM ... 74
A . Lb Twh 5 U / TLh b... 74 1.Contexte ...74 2.Rappel des objectifs e t stratégie ...75
. . tw9a L9w A wTL/ [ 9 ... 75
/ . 5LS / U S S Lh b ... 108 1.Absence de données de F AIRE -seq chez les fem elles sexuées ... 108 2.Challenge bioinform atique : des données de qualité m oyenne... 110 3.Identification de longs ARN n on-codants m âles-biaisés ... 112
VI. CHAPIT RE II : R EG ULAT IO NS CHRO M AT IENNES IM PLIQ UEES D ANS LE PO LYPHENISM E D E REP RO D UC T IO N D U PUCERO N D U PO IS ... 115
A .Lb Twh 5 U / TLh b... 115 1.Contexte ... 115 2.Rappel des objectifs e t stratégie ... 115
. . S9/ h b 5 A wTL/ [ 9 ... 116
/ . w9S U [ TA TS / h a t [ 9a 9b TA Lw9S ... 149 1.RNA-seq : un potentiel m anque de profondeur de séquenç ag e par rapport aux donnée s FAIRE-seq
149
0. T ab le d es m atières 0.
VII. D IS CUSS IO N GENE RALE ... 154
A . [A a 9TH h 5 9 CA Lw9-S 9v : A ± A b TA D9S, Lb / h b ± 9b L9b TS 9T [ La LT9S... 154 1.Choix de la m éth ode FAIRE-seq vis-à-vis du D NAse-seq et de l’AT AC-seq ... 154 2.L’analyse de données FAIRE-s eq ... 157
. . [9 D9b h a 9 5’A . t LS U a : U b 9 A b b h TA TLh b 9T U b A S S 9a . [ A D9 Lb / h a t [ 9TS... 162 1.Le génom e d’A. pisum : état de l’art ... 162 2.L’incidence des lim ites du génome d’A. pisum sur l’analyse de données de régulation d e la
transcription ... 163
/ . /Y/ [ 9 5 9 ± L9 5 9S t U / 9wh b S : t h [ Yt H 9b LS a 9 9T / h a t 9b S A TLh b 5 9 5 h S 9 ... 170 1.Un m écanism e de com pensation de dose perm ettrait d’éq uilibrer la transcription du chrom osom e X vis-à-vis des autosom es ch ez les pucerons m âles ... 170 2.Le polyphénism e de reproduction des pucerons es t poten tiellem ent associé à différentes voies de signalisation neuro-endocrine ... 172
VIII.PERS PECT IVES ET CO N CL US IO N ... 178
I. Introd uction générale 0.
I
.
I
ntroduction générale
Les organismes vivants doivent constam me nt et rapideme nt faire face au x fluctuations constantes d e leur enviro nne me nt biotique et abiotique pour as surer leur survie. Si le patrim o ine génétiq ue dont ils héritent est fixe, il leur est néa nmoins possible de moduler leur phénotype pour m ie ux s’ada pter aux contraintes e nvironnem e ntales. Cette mod ulation du p hénotyp e pe ut nota mm ent se faire par l’express ion différe ntielle rapid e et coordonnée d e différe nts sets de gènes, à sa voir des pro gra mm es génétiq ues alternatifs. Il leur est ainsi possible de produire à partir d’un g énotyp e unique une continuité de p hénotypes spécifiqu em ent a daptés à un e nse mble de conditio ns environnem entales différe ntes et fluctuantes . U n tel m écanism e ad aptatif es t com m uném ent appelé plasticité phénotypique et génère d es normes de réaction, définies com me la prod uction par un génotype donné d ’une gra dation continue de p hénotyp es en répo nse à la mod ulation d ’un facteur environnem ental [G riffiths et al., 2000].
Chez l’Hom m e la plas ticité phénotypique p eut par e xem ple p erm ettre d ’ada pter la m usculature d’un individ u selon le régime d’exercices auq uel il est confronté en générant une continuité de phénotyp es musculaires adaptés à ces différe nts efforts, et ce à partir d’un génotyp e fixe [ H op pele r et Flü ck, 2002]. Ainsi, en réponse à des exercices d’e ndurance et d e faible intensité, un individu prés ente ra des changem ents q ualitatifs d e fibre m usculaire caractérisés par une augm entation du volume des structures transporta nt et consomm ant l’oxygène, tandis qu’un individ u réalisant des exe rcices inte nses de force brute comportera une m usculature caractérisée par une q uantité plus importa nte de protéines co ntractiles [Hoppeler et al., 2011] comm e l’actine et la m yosine [Barde n et al., 1988]. Ces phénotyp es musculaires différents et adaptés au type d’effort s ont ainsi perm is par la m ise en place de pro gra mm es génétiq ues .
La plasticité phénotypiq ue est égale me nt im portante chez les vég étaux q ui sont des orga nism es imm o biles pour qui une adaptation a ux conditions environne me ntales est donc particulière me nt cruciale [G ratani, 2014 ]. Chez la plante mod èle Ara bidopsis thaliana, e n cas de dévelop pe me nt en milie u s alin, un génotyp e donné prod uira une co ntinuité de p hénotypes suivant la concentration en sel du m ilie u. C es phénotypes corres pond e nt nota m m ent à une réduction globale de la taille de la plante, une floraison tardive, une diminutio n du nomb re de ram ifications , ainsi que des sections entre nœ uds et des inflores ce nces plus co urtes [K ooke et al., 2015], l’e nse mble de ces phénotypes adaptatifs perm ettant ainsi à la plante de résister à des concentrations salines élevées.
Les organis m es ectothe rm es, comm e par exe mple le s insectes, adaptent la taille de le ur corps à la tem pérature de le ur environne m e nt [ Steve nson, 1985]. C ette plasticité p hénotypique se trad uit par une augm e ntation de la taille du corps d’un individu adulte lorsque celui-ci est exposé à de bass es tem pératures au co urs d e son dévelop pem e nt, et rés ulte d ’une a ugme ntation d e la taille des cellules plus que d’une au gm entation de leur nom bre. Ce mécanisme perm et de limiter les pertes de chale ur dues à de faibles te mp ératures en réd uis ant propo rtionnelle m e nt la surface de contact du corps d es
I. Introd uction générale 0.
ectothermes avec l’air froid environna nt [French et al., 1998]. Chez la D rosophile, la taille du corps et la taille des ailes doive nt res ter pro portionnelles afin d e maintenir une stabilité lors du vol. Ainsi, les D rosophiles élevées à de faibles tem p ératures présente nt des ailes plus grand es que chez d es D rosophiles élevées da ns des tem pératures médianes [Stanley, 1935], et cette plasticité phénotypiq ue est as sociée à une augm e ntation de la taille des cellule s constituant les ailes [D e Moed et al., 1997].
Concerna nt ce s exem ples , et dans la majorité des cas observés dans la nature, la plasticité phénotypique est continue. D ans d’autres cas plus rares et spectaculaires ap pelés polyp hénis mes [Mayr, 1963], la plasticité phénotypique es t discrète, avec la prod uction d ’un nom bre défini d e phénotyp es par un s e ul e t m êm e génotyp e en répons e à u n facteur e nvironnem e ntal do nné. Les exe m p les les m ieux caractérisés chez les insectes correspondent au polyp hénism e de cas te chez les abeilles (Figure 1), comm e par ex em ple A pis m elifera [Evans et W heele r, 1999], ou chez les fourmis telles que Cam po notus floridanus [Bonasio, 2014 ]. Ainsi, les colonies s ont composées d’individ us ayant différe nts phénotyp e s discrets associés à des fonctions spécifiques au sein de la ruche ou de la fourmilière . Ces castes peuve nt par ex em ple correspondre à des ouvrière s, des soldats, des explo ratrices ainsi qu’à des reines, lesq uelles sont les seules cap ables de se reproduire au sein de la ruche ou de la fourmilière , et donc d’as surer la survie de la colonie. Chez ces deux organis m es, le régime nutritif auquel sont soumises les larves est le facteur enviro nnem ental maje ur permetta nt d’orienter le ur d eve nir et faç onner leur p hénotype à l’âge a dulte. Ainsi chez les a beilles , selon que les larves soient nourries avec de la gelée de type royale ou com mune, q ui diffère nt notamm e nt par leur com position en Acid e (E )-10 -hydro xy-2 -d écénoïque (10HD A) [Viuda -Martos et al., 2008], des progra m m es génétiques alternatifs vont être m is en place et aboutir à l’express ion des phénotypes reine ou ouvrière , respec tiveme nt. Selon deux co nditions nutritives distincte s, deux p hénotypes prése ntant des caractéristiques morp hologiq ues et fonctionnelles très différente s s ont donc produits à partir d’un m ême g énotype (Fig ure 1).
D e faç on plus générale , les polyphénis mes chez ce s insectes font d’abord intervenir la perception d’un signal environneme ntal p ar l’organis me suivie de sa transduction jus qu’à des tissus cibles , et aboutissant à une rep rogram matio n de l’e xpression des gènes au niveau de ces tissus. Chez les fourmis et les abeilles, il a été montré q ue la mise en place de ces reprogra m mations transcriptio nnelles rapides impliq uait des régulatio ns épigénétiq ues. Ces dernières sont définies com me une modification de l’expression héréditaire des gènes sans modification de la séquence d’AD N, générale me nt e n répons e à l’e nvironne m e nt [W addingto n, 194 2, 2012]. D ’après l’exem ple précédem me nt décrit, les m éca nis mes épigénétiq ues peuvent par ex em ple participer au changem ent du p hénotype d ’un individ u, e n répo nse à des stim uli environnem e ntaux et sans m odification de so n génotype. Elle im pliq ue différe nts m écanism es te ls que la m éthylation d e l’AD N (cytosines notam me nt), les modifications post-traductio nnelles des his tones, ou e ncore l’e xpression de certains ARN non-coda nts. Chez A. m elifera, un nive au d e m éthylation différe ntiel a par exe mple été id entifié entre des larves nourries avec de la gelée royale et d’autres nourries avec de la gelée com m une [Foret
et al., 2012]. Chez C. floridanus, un nivea u différentiel global d’acétylatio n de la lysine 27 des histones H3 (H 3K2 7ac) a par exe m ple été id entifié entre les cas tes ouvrières maje ure et m ineure [ Simola et al., 2013b], qui se différencient par le ur co m portem ent et les tâches q u’elles accom p lissent au sein de la
I. Introd uction générale 0.
fourmilière . En revanche, la contribution de m écanis mes épig énétiq ues dans la mise en place du polyphénis me de re pro duction chez les pucerons n’a jus qu’alors pas été étudiée. O n pe ut ainsi émettre l’hypothès e q ue la signalisation neuroe ndocrine d’une diminution de la photop ériod e au s ein d’individ us asexués pe ut être à l’origine d ’une modification rapid e de l’orga nisation de la chromatine des tissus cibles – ici, les lignées germinales e mbryonnaires – à l’aide d e régulations épig énétiq ues permettant ainsi la mise en place des phénotyp es s exués.
Fig ure 1 | Le polyp hénisme d e caste chez l’ab eille. La biologie des abeilles com p rend des inte ractions sociales entre individ us d e différentes castes, prod uites par la mise en place d’un polyp hénism e. Pendant le d éveloppe m ent larvaire, les larves fem elles alime ntées par de la gelée royale (haut) se dévelop pe nt plus vite e n prod uis ant des ab eilles re ines. Celles-ci ont une d urée de vie allo ngée, exp rim ent d es phérom ones particulières, et sont capables de se reprod uire. Les larves femelles nourries d e gelée ouvrière ont qua nt à elle un développe me nt plus le nt et pro duise nt d es abeilles ouvrières, notam me nt caractérisées par la présenc e de paniers à pollen (corbicula). D ’après [Cridge et al., 2015].
D e faç on concomitante, le polyphénis me de re pro duction des p ucero ns a pour conséq uence la prod uction de trois morp hes sexuels au cours du cy cle de vie annuel des p uce rons : des fem elles asexuées vivipares, d es femelles sexuées ovipare s et des m âles. Ces trois m orphes sont génétiq ueme nt ide ntiques à l’exce ption d e leurs chro m osomes sex uels, p uisq ue les deux typ es de femelles comportent d eux chromosom es X, tandis q ue les mâles n’e n compo rte nt qu’un [Blac kma n, 1987]. Les p ucero ns corres ponde nt ainsi à une e spèce hétéroga m étiq ue mâle , co mporta nt un système chromosomique du type XX / X 0. Un sys tèm e déséquilib ré comm e celui-ci pose la question
I. Introd uction générale 0.
inhére nte à toute espèce hétéroga m étique : qu’e n est-il de la régulation d u niveau global de l’express ion des gènes portés par le(s) chro mosome (s ) X vis-à-vis des gènes portés par les autosomes chez les mâles et les femelles, puisq ue ces derniers n’ont pas le m êm e no mbre d’allèles pour chaq ue gène sur le chro m osome X ? Chez plus ieurs organis m es hétéroga métiques, d es m écanis mes dits de com pe nsation de dose ont été identifiés, im pliquant généralem ent des mo difications épigénétiques permettant d ’équilibre r la transcription des gènes portés par le chro mosom e X entre m âles et femelles. O n sait que chez le puceron du pois, les gènes du seul chromosom e X des m âles sont en moyen ne plus exprimés q ue ce ux portés par les deux X d es fe melles pour de s gènes autosoma ux globale me nt exprimés de manière éq uivale nte [Jaq uiéry et al., 2013]. O n peut donc ici émettre l’hypothèse qu’il existe rait un m écanis me global de régulation de l’e xp ress ion d e s gènes portés par le chrom osom e X, p utativem ent d e com pensation d e d ose, im pliq uant des rég ulations épig énétiq ues chez les puce rons.
Les travaux réalisés au cours de ma thèse visent donc à étudie r la résulta nte d es méca nism es épigénétiq ues sur l’accessibilité d e la chromatine et la transcription d es gènes a u nivea u du g éno m e d’Acyrthosipho n pisum (p uceron du pois) - es pèce ap hidienne m o dèle utilisée p our cette étude - da ns
le cadre : i) de la pote ntielle compe nsation de dose p ouvant avoir lieu au niveau des gènes portés par le chromosom e X des m âles et des fem elles , et ii) de la m odification d es prog ram m es génétiq ues à l’origine d e la trajectoire de d évelop pem e nt s exué ou asexué des e m b ryons dans le cadre du polyphénis me d e re prod uction.
Nous avons dans un p re mier te mps étud ié les profils d’e xp ression du ch romoso me X e ntre m âles et femelles (par RNA-se q) p uis le niveau global d ’accessibilité de la chro m atine de ce chromosom e entre mâles et fem elles, via notam m ent l’utilisatio n de la techniq ue d u FAIRE-seq (Formald ehyde Assisted Isolation of Regulatory Eleme nts suivi d ’un séq uenç age ha ut-débit) afin de tester l’hypothèse de l’existence d ’un m écanis m e global d e com pensation de dose. D ans un s econ d tem ps, nous avons com paré par RNA-se q les transcriptom es d’em bryo ns à devenir sexué ou as e xu é puis étudié le nivea u global d’ouv erture de le ur chrom atine par FAIRE-se q afin d’id entifier et caractériser des régions d u génom e différe ntielle m ent acces sibles et potentielle me nt impliquées dans la mise en place des phénotyp es alternatifs.
Afin de répondre à ces q uestions nous introd uirons tout d’a bord les éléme nts de bibliogra p hie e n relation avec ces différe ntes questions , p uis nous prés ente rons une vision synthétique des techniq ues moléculaires et bioinform atiques com munes à l’e nsemble de ces q uestions. Par la suite, nous prése nterons et discutero ns les principa ux m éthod es et résultats d évelop pés d urant cette thèse pour conclure s ur leurs implications dans le domaine de l’étude des méca nismes épigénétiq ues chez les puce rons dans le cadre de son cycle de vie et de son mode rep roductio n.
II. Synthèses b ibliographiq ues A. Biologie de s puce rons
I
I
.
Synthèses bibliographiques
A. Biologie d es p uce rons
1. Les puce rons : d es ravage urs m ajeurs d e s culture s ag ronom iques
Les pucerons sont des insectes hém iptères de la fam ille des A phididae com ptant plus de 4700 espèces identifiées à ce jour. Ce sont des insecte s pique urs -suce urs de plante se nourriss ant exclusive me nt de la s ève élaborée issue du phloème. Ces ins ectes sont à l’origine de dégâts importa nts sur des plantes de g randes cultures (céréa les, fabacées…), s ur ce rtaines plantes horticoles et sur d es arb res forestiers ou fruitie rs. Ces insectes p rovoque nt d eux gra nds typ es de d égâts sur les plantes au dépe nd d esquelles ils se nourrissent (Fig ure 2).
a. D égâts directs
Le stylet des pucero ns est l’organe qui p ermet le ur nutritio n : cette structure pénètre les tiss us végétaux jusq u’a u phloèm e et perm et de prélever la sève élaborée, provoq uant ainsi des domm ag es directs sur la pla nte (bless ure , d étourne me nt d e ressources nutritives ). Ava nt d e pas ser en phas e de nutritio n à pro pre m e nt p arler po uvant d urer d e q uelques he ures à plusie urs jours, le p ucero n effectue de nom bre uses piqû res d’essai afin d ’évaluer l’acceptabilité de l’hôte végétal. Si l’hôte convient, le stylet d u p uceron va atteindre le p hlo èm e e n s’infiltrant entre la paroi d es cellules végétales d e l’épide rm e, afin d’initie r la phase de nutrition pe rm ettant une abso rption d e nom breux photosynthétats. Ces d erniers étant essentiels au développ em ent de la p lante , les puce rons p euve nt ainsi entrainer une croissance ralentie, une mauvaise fructification, ou la production de grains ou de fruits de taille plus faible et aux vale urs nutritio nnelles et co mme rciales am oind rie s. En outre , au cours de la piqû re, les pucerons s écrètent une salive liquide et une salive visqueuse, accompag nées de com posés chimiq ues et d e peptid es agiss ant sur la plante e n perturbant ses réactions de défe nse, em pêcha nt nota mm ent la réaction norm ale d es tubes criblés d u phloèm e en cas de blessure , à savoir une occlusion dép endante du calcium [T jallingii et Es ch, 1993 ; Miles, 1999 ; W ill et al., 2007].
Enfin, le m od e de nutrition des puce ro ns entraîne l’e xcrétion d’un miellat composé principalem ent d e fructose et glucose extraits du p hloèm e et no n assimilés par le p uce ron [La mb , 1959]. C e miellat pe ut d’une part e ntrainer le d esséche me nt d’organes q ui en sont recouverts, et il peut d’a utre part promo uvoir le dévelop pem e nt d e cha mpignons ca usant d es fum agines, réd uisant ains i l’activité photosynthétiq ue des feuilles atteintes [D edryve r, 20 07].
II. Synthèses b ibliographiq ues A. Biologie de s puce rons
Fig ure 2 | Exem ple d e dégâts causés p ar les p uce rons sur les cultures . A) Une femelle as exuée ad ulte de l’es pèce Acyrthosip hon pisum, le puceron d u pois. B ) Colonie de pucero ns du pois sur un rosier. C) Sym ptômes d u virus de la mozaïq ue d u pois (PEMV) transmis par le p ucero n du p ois.
b. D égâts indirects
Les pucero ns provoquent égalem e nt des dégâts indire cts à leurs plantes hôtes pa r la transm ission de phytovirus lors de leur p ha se de nutritio n, en s e déplaç ant et se nourrissant d’une plante à l’autre. Les symptôm es causés par ces infections sont dépendants du virus transmis et sont très divers, pouvant aller d’une simple décolora tion des feuilles (chlorose ) à la mort de la plante. Par exem ple, le virus de la jaunisse nanisante de l’orge (BYD V) est transm is par plus de 25 espèces de pucerons [D u et al., 2007] et est responsable de pertes de récoltes allant d e 10 à 85% par parcelle [B ra ult et al., 2007]. L es phytovirus circulant au sein du phloèm e de la plante peuve nt être trans mis par les pucerons de manière circula nte ou non-circulante.
Les virus non-circula nt s ’a ttachent au nivea u d ’une s tructure protéiq ue particulière d u stylet du puce ron : l’acrostyle [Uzest et al., 2010]. Au cours des piqû res d’essai ou de nutrition, les virus non-circulant s’y attachent dès la pénétratio n du stylet dans les tissus de la plante hôte, pour être ens uite relâchés lors des piqû res d’épreuve s uivantes, et po tentiellem ent s ur une no uvelle pla nte. Il e xiste ainsi de ux stratégies d’accrochage d es virus su r le stylet des p uce rons [Bra ult et al., 2010]. D ans le
II. Synthèses b ibliographiq ues A. Biologie de s puce rons
cadre d e la stratégie dite « cap side », nota mm ent utilis ée par les Cucum ovirus, l’accrochage d u virus au stylet s’effectue par l’interaction directe e ntre les p rotéines constituant la cap side d u virus et des réce pteurs aphidiens situés sur la cuticule du stylet. D ans le cadre de la s tratégie dite « helper », l’accrochage du virus s’effectue par l’interm édiaire d ’un facteur vira l assistant l’inte raction e ntre la capside et un réce pte ur situé sur le stylet du pucero n. C ette seco nde stratégie est par exem ple utilis ée par les Caulimovirus et les Potyvirus [Ng et Falk, 2006].
Certains virus sont transmis selon un mode circulant [Ng et Perry, 2004], auquel cas ils transitent à l’intérie ur d u puce ron (intestin, hém olym p he) avant d’atteindre les glandes salivaires où les virions s’accum ule nt au niveau d ’e spaces intercellulaires connectés à la lum ière des cond uits s alivaires afin d’infecter une nouvelle plante via la s alive émise par le puce ron lors d e piq û res de nutritio n ou d’essai [Amm a r et Na ult, 1985 ; Brault et al., 2010]. Les virus transmis s elo n ce mode circulant p euv ent être propa gatifs ou non-propag atifs selon qu’ils se m ultiplient ou non à l’intérie ur du puce ron [Brault et al., 2010]. Les Ra hb dovirus sont des virus circulant propag atifs qui libère nt leur géno me viral au niveau du cytoplasm e d es cellules de l’intestin po ur s’y répliquer de m anière lente, sans cau ser de dom m ag es au puce ron. Après leur m ultiplication au niveau des cellules de l’intestin, les particule s virales s’échappent en traversant le plasm ale mm e bas al. Une fois libérés dans l’hém olym p he, les virions ont accès à différe nts orga nes, dont no tamm ent les organes re pro ducte urs q ui le ur p ermette nt une tra nsmission verticale de puis la fem elle infectée vers ses desce nda nts [Sylves ter, 1969 ; Nault, 1997]. Ce processus se déroule lente me nt com pte tenu d u nom bre d e barrières physiologique s à traverser (gaine ovarienne et ép ithélium folliculaire). Parmi les autres organes accessibles aux virions d ep uis l’hémolymp he, o n com pte le cervea u, les ganglions ne rveux, l’épide rm e, les tiss us conducte urs et de réserve , la rétine, les musc les ainsi que la trachée [Amm a r et Nault, 1985]. Les virus non-pro pagatifs adopte nt le mêm e trajet de circulation au sein des pucerons, sans toutefois s’y multiplier [Brault et al., 2010].
2. Cycle de vie e t a lternance d u m ode de re prod uction
Il existe principale me nt d eux types d e cycles de vie chez les puce rons : certains co mportent une parthéno génèse cycliq ue (CP pour « Cyclic Partheno genesis) tandis q ue d ’autres comporte nt une parthéno génèse obligatoire (O P pour « O bligatory Parthenoge nes is).
La plupart des espèce s et populations de pucerons sont CP, et alternent au cours de l’année un mod e de repro duction parthénogénétiq ue clonal avec une repro ductio n sexuée, et sont ainsi dites holocycliques (Fig ure 3). Au printe mps et en été, le s populations sont constituées d e femelles à rep rod uction parthénogénétique apomictique thélytoque vivipare. Ce m ode d e re pro ductio n correspond ains i à une repro duction ase xuée , sans fécondation et sans m éiose sensu stricto (apomixie ), ne pro duisa nt q ue d es femelles (thélytoquie) se dévelo ppa nt da ns l’ab dome n d e leur mère sous forme d’em bryo ns (viviparité). La ra pidité de pro duction d es desce ndants (jusqu’à 100 individus prod uits par adulte) confère aux pucerons une forte capacité de co lonisation de leur e nvironne m e nt. A la fin de l’été ces femelle s vont percevoir la diminution de la photop ériod e - ainsi qu’une diminutio n
II. Synthèses b ibliographiq ues A. Biologie de s puce rons
de la température - en me s urant la durée de la nuit (scotophas e) et en perceva nt une accum ulation d’un nom bre minim um de scotophases longues consécutives. En répo nse à ces stim uli, les fem elles parthéno génétiques vont prod uire dans leur descendance des femelles s exuées ovipares et des mâles . Après accouplem e nt, les fe m elles sexuées vont pond re des œ ufs fertilis és corresp o nda nt à une forme d ’ada ptation a ux hivers rigoureux, puis que ce s œ ufs sont capables de résister à de faibles tem pératures, no rmalem e nt létales pour les ad ultes et les larves. U n œ uf ains i pondu va rester e n diapa use p enda nt e nviron 90 jours durant lesquels l’e m bryo n conte nu dans cet œ uf se dévelo pp e de manière rale ntie [S hingleto n et al., 2003]. A la fin de cette période de dia paus e, c’est-à-dire à l’arrivée du printe mps , les œ ufs vont éclore en donna nt naiss ance à des femelles dites « fondatrices » q ui sont des fe melles parthénogénétiques vivipares p rod uisant uniq ue m e nt d es fe melles as ex uées. E n conditions contrôlées , ces « fondatrices » s ont incapa bles de répo ndre à une photopériod e courte et ainsi de prod uire des des cendants sexués pe nda nt plus ieurs générations. C’est « l’effet fondatrice » [Lees, 1960], q ui limite probable me nt la prod uction de morp hes sexuées au déb ut d u printe mps lorsque la p hotopériode est encore co urte et pote ntiellem e nt inductrice.
Fig ure 3 | Cycle de vie a nnuel et polyp hénism e d e re production d ’Acyrthos iphon pisum. Le cycle de vie des pucerons est basé s ur un polyphénism e de reprod uction caractérisé par une re prod uction asexuée clonale vivipare thélythoq ue au printem ps et en été, et par une rep rod uction sexuée ovipare en hiver. L ’alternance d u m ode de rep rod uction est fonction d’une répons e à la diminutio n annuelle de la photop ériod e. Au cours de la phas e de re pro d uction asexuée , des m orphes ailés peuve nt être prod uits en réponse à une surpopulation de la co lonie ou en réponse à une dim inution d e la qualité nutritive de la plante. Adap té d’a près [Jaq uiéry et al., 2013].
II. Synthèses b ibliographiq ues A. Biologie de s puce rons
D e nombreuses espèce s de pucerons comporte nt des populations (ou génotyp es) présentant un cycle parthéno génétique obligatoire (O P, Figure 4). Ces clones s ont caractérisés par une réponse altérée à la diminution de la p hotopériode, car ne prod uisant da ns leur desce nda nce q ue d e s femelles asexuées vivipares, voire quelq ues m âles [Simon et al., 2002, 2010]. D e par l’abse nce d e production d’œufs, ces lignées O P sont sensibles au froid : elles sont do nc contre -sélectionnées dans les climats prés e ntant des hive rs rigoure ux. C ’est pourquoi l’o n retro uve principale me nt ce s lignée s dans des régions géogra phiq ues présenta nt des hivers do ux ou une ab sence d ’hive r, où les pop ulations O P présentent un avanta ge d émog rap hiq ue certain vis-à-vis des lignées CP [Rispe et Pierre, 1 998 ; D edryver et al., 2001]. Récipro quem ent, les lignées CP sont m ajoritaire s au niveau des régions géograp hiq ues froides, tandis q ue les de ux typ es d e lignées coexistent da ns d es régions te mp érées prés enta nt des hivers aux tem pératures fluctua ntes d ’une a nnée s ur l’a utre [D ed ryver et al., 2001 ; Simo n et al., 2002]. Alors q ue le passage de la repro duction clonale à la reproduction sexuée chez les lignées CP est déclenché par des change m e nts de la p hotopériod e, la pe rte d e re production se xuée est supposée génétiq ue chez les lignées O P, p uisq ue les changem ents de conditions environne me ntales n’ont aucun effet sur leur phénotyp e re prod ucteu r [Simon et al., 201 0 ; D edryver et al., 2013 ; Jaq uiéry et al., 2014].
Fig ure 4 | Cycle de vie sim p lifié d u puce ron du pois. G auche : populatio ns C P alternant le ur mo de de rep rod uction entre les saisons. D roite : populatio ns O P n’alternant pas le ur m ode de rep rod uction, restant ainsi asexuée toute l'an née. D es m âles sont cependant p roduits e n autom ne.
II. Synthèses b ibliographiq ues A. Biologie de s puce rons
3.
T raits d ’
histoire de vie et p lasticité p hénotypiq ue
a. Bactéries e ndosym biotiq ues
Les puc erons com porte nt de nom bre ux traits d’histoire d e vie q ui e xpliquent leurs re marq uables capacités d’a daptatio n aux contraintes d e le ur e nv ironnem ent proche. Ils comportent ainsi d es bactéries endosym biotiq ues de différe ntes espèces et genres, le ur perm etta nt par exe m ple une meilleure adaptatio n aux p lantes hôtes au dépe nd d esquelles ils se nourrissent. Buchnera aphidicola est la s eule bactérie endosymbiotiq ue o bligatoire che z de no m b re uses esp èces de p ucero ns. Cette symbiose obligatoire est caractérisée p ar la localisation intrace llulaire de B uchnera aphidicola au niveau du cytoplas me des bactériocytes (adipo cytes sp écialisés co ntenant d es bactéries endosym biotiques) et par la transmission verticale de cette bactérie de la m ère à s es descendants [D ouglas, 1998]. Cette symbiose perm et à la fois aux puce rons et a ux bactéries d e se nourrir efficaceme nt. La sève élaborée dont les pucero ns se nourrissent est dépo urvue de ce rtains acides aminés ess entiels, com me le tryptopha ne ; ces acides am inés sont synthétisés pa r Buchnera [Lai et al., 1994] q ui ass ure égale me nt le recyclage de l’azote, et qui en écha ng e reç oit des composés carbonés et azotés de la part du méta bolisme du p ucero n hôte. Les pucerons comporte nt égale me nt un certain nom bre de bactéries end osymbiotiq ues secondaire s , leur conférant u ne gra nd e variété de traits phénotypiques. Actuellem e nt, 8 es p èces de bactéries seco ndaires ont été ide ntifiées chez le p uce ro n du pois Acyrthosiphon pis um : cinq G am map roteoba ctéries qui sont Ham iltonella defensa, Regiella insecticola, Serratia sym bio tica [Moran et al., 200 5], Rickettsiella viridis [T suchida et al., 2010, 2014] et PAXS (Pea Ap hid X-type Symbio nt) [G uay et al., 2009] ; de ux Alphaproteo bactérie s du g enre Rickettsia [Che n et al., 1996] et W olbachia [Russell et al., 2013] ; et Spiroplas m a [F ukatsu et al., 2001] appartena nt a ux Mollicutes . Ces bactéries so nt localisées dans différe ntes parties du puce ron du pois telles que l’hém olym phe ou les bactériocytes [Moran et al., 2005]. C es bactéries s ymbiotiq ues facultatives sont impliquée s dans divers processus biologiques tels qu’une valeur adaptative (fitness) accrue sur des plantes hôtes spécifiques [T suchida et al., 2004], des modifications de la couleur du corps [T suchida et al., 2010], une toléra nce accrue à la chaleur [Montllor et al., 2002], ou encore une protection contre des ennemis naturels [O liver et al., 2003 ; Scarborough et al., 2005]. Bien q ue transm is es de manière verticale comm e Buchnera aphidicola, ces bactéries sym biotiques facultatives peuvent aussi être trans m ises horizontale me nt entre pucerons de la m êm e espèce ou d ’es pèces différe ntes [He nry et al., 2013 ; G authier et al., 2015]. Notons que seul un no mbre réd uit d’es pèces de bactéries e ndosymbiotiq ues cohabitent a u sein d’un m ême p ucero n, et que différe ntes ass ociation de ces bactéries avec Buchnera aphidicola pe uve nt se trouver au sein d’une mêm e colonie.
II. Synthèses b ibliographiq ues A. Biologie de s puce rons
b. Plas ticité phénotypiq ue et polyphénis me chez les puc erons
D ’une ma nière générale , un organis me ayant un génotype do nné est capa ble d e pro duire un continuum de phénotypes en réponse à une condition e nvironne m e ntale particulière (plasticité phénotypique continue). M ais chez certains orga nismes, un mêm e génotype est capable d e pro duire des phénotypes discrets extrême m e nt différents en répo nse aux fluctuations de facteurs environnem enta ux. Ce phénom ène corres pond a u polyphénis me (d étaillé e n E. de cette synthèse bibliog rap hiq ue ) prése nt chez de nom breux insectes dont les pucerons. Chez ces derniers , il existe trois types de polyphénis m e : le polyphénis m e de dispersion, de ca ste et de reprod uction. Cette plasticité phénotyp iq ue dis crète explique en grande p artie la rémanence de ces ravageurs a u sein d es agroécosystèm es ainsi q ue l’im portance des dégâts causés sur les cultures.
Polyp hénism e de disp ersion
Les puce rons sont capa bles de répondre à une diminution d e la q ualité nutritive de la pla nte et/ou à une surpopu lation causée par le urs congénères sur une m ême plante. Les fe melles perç oive nt ces signaux et y réponde nt e n pro duisa nt da ns leur d es cenda nce d es fem elles ailées, capa bles d e coloniser de nouvelles pla ntes en as sura nt ainsi le m aintien d e la population sur une autre pla nte prése ntant une m eille ure q ualité nutritive [D ixon et Howard , 1986 ; Zera et D enno, 1997]. Ce type de polyphénis me a lieu au cours de la phase de repro duction asexuée des pucerons et leur confère une importa nte cap acité de colonisation. Une réce nte étude a mis en évidence q ue des gènes impliqués dans la voie hormo nale de l’ecdysone sont sous-exprim és chez les fe m elles pro ductrices d’individ us ailés [Vellichiram mal et al., 2017]. Ces gènes prés entent d es ho m ologues chez la D rosophile do nt ultraspiracle et ecdysone receptor (EcR), deux gènes co dant pour d es protéines impliquées dans la
formation hétéro dim érique du récepteur à l’ecdysone, ains i que des gènes dont l’exp ress ion est induite par l’ecd ysone, dont broad et ftz-fl. D e plus , des individus ayant subi des injections d’ecdysone prod uis e nt m oins d’individ us ailés , tandis que les individus traités avec un antagoniste d’un récepteur à l’ecdysone – ou traités pa r RN Ai ciblant le gène coda nt pour ce récepteur à l’ecdysone – pro duise nt davantag e d’individ us ailés dans leur desce nda nce vis-à-vis des individ us contrôle. Ceci démontre un lien de causalité e ntre le polyphénis me de disp ers ion des puce rons et la voie horm o nale de l’ecdysone.
Polyp hénism e de caste
Chez les insectes sociaux, certains individ us sont spécialisés dans la re pro duction, tandis q ue d’a utres ne prod uise nt que peu de - voire aucun - des cendants. Ces m orphes peu fertiles , voire infertiles, sont caractérisés par un co m portem ent et d es traits physiologiques distincts d es individus re prod ucte urs, en éta nt généralem e nt sp écialisés dans d es fonctions altruistes. Ces phénotyp e s altruistes infertiles sont prod uits en répo nse à des facteurs e nviro nnem enta ux précis et correspond ent à des polyphénis mes de ca stes, notamm ent présents chez certains p ucerons [Aoki, 197 7]. Certaines esp èces de p ucero ns prod uise nt e n effet des individ us sacrifiant leur capacité de repro duction afin d’a m éliorer la survie d e le urs congénères. C es individus sont co ns idérés comm e des « s oldats » puisq ue le ur rôle social prim aire correspond à la défense de la co lonie (Figure 5), m ême si certains de ces individus peuvent en plus comporter des rôles dom estiques [Aoki, 1980 ; Be nton et Foster, 1992]. Actuellem ent,
II. Synthèses b ibliographiq ues A. Biologie de s puce rons
60 esp èces de puce rons prod uctrices de soldats ont été identifiées parm i d eux so us-fam illes de puce rons (Pem phigina e et Horm a phidinae) [Aoki, 1987 ; Itô, 1989 ; Stern et Foster, 1996 ; Fukats u et al., 2005 ; Pike et Foster, 2008]. Chez T ubera phis styraci, la production de soldats dans la descendance des femelles es t corrélée positivement avec la densité en individus et la taille de la colonie. D e plus , la perce ption de ces sig naux environne m entaux se mb le être à la fois pré et post-na tale, indiq ua nt do nc une forme d e trans m ission de ces signaux de la m ère à sa descendance [Shibao et al., 2010].
Fig ure 5 | Photograp hies de la caste rep rod uctrice (A) et soldat (B) d e l’esp èce Pseudoregma bambusicola, le puce ron du ba mbou de la fa mille des H orm a phidinae. Les soldats sont prése nts e n
grand nom b re au s ein d es colonies et assure nt la défe nse d e celles-ci grâc e à la longueur et la larg eur importa ntes de le ur prem ière paire de pattes (flèc he blanche), leur p erm etta nt de re pouss er certains pre date urs.
Polyp hénism e de rep rod uction
Comm e d écrit plus en am o nt, les pucero ns com porte nt un polyphénis me de re production au cours de leur cycle de vie ca ractérisé par un basc ulem e nt d’une repro duction as e xuée ve rs une re pro duction sexuée en répons e à la dim inution d e la photopério de à la fin de l’été. En répo nse à ce stimulus, les femelles asexuées parthénogénétiques vivipares perç oivent la diminution de la photopério de, intèg re nt ce signal, p uis le trans duise nt via le système ne uro endocrine aux organes cibles, les em bryons. Ces derniers y répo nd ent par une modification de leur trajectoire d évelopp em entale, et notam me nt un changem en t du deve nir de le ur lignée germinale, pass ant d ’ovocytes diploïd es q ui se dévelop pe nt en e mbryo ns dans le ca dre de la repro d uction asexuée à une lignée germ inale haploïd e (gam ètes) p rod uisant soit des ovocytes pour les futures fem elles ovipares, s oit des sperm atozoïdes pour les mâles dans le ca d re de la re prod uction sex uée. Le génom e des e mb ryons est donc plas tique , car capable d e mettre en place différe nts prog ram mes génétiques en réponse au sig nal neuroe ndocrine afin d e pro duire ces trois morphes discrets.
II. Synthèses b ibliographiq ues A. Biologie de s puce rons
Chacun d e ces morphes est adapté à l’environnem e nt au sein duq uel ils s ont produits, avec des femelles parthénogénétiq ues adaptées à une re prod uction et une colonisation rapide des agroécosystèm es dans le cadre de conditio ns favorables (printem ps et été), et des mâles et femelles sexuées adaptés à un mode de survie de la co lonie dans le cad re de conditions hivernales défavorables. U ne rés ultante d e ce polyphénis me de re pro duction est q ue ces trois morphes prése ntent des ap pareils re prod ucteurs distincts.
c. L’ap pareil rep rod ucteur des femelles as exuées et sexuées
Les pucero ns fem elles prés entent d eux ovaires constitués d e plusieurs ovarioles, chacun étant constitué d ’un ge rma rium et d’ovocytes haploïd es se différenciant en œufs non-fertilis és chez les femelles ovipares ou d’ovocytes diploïdes se différencia nt e n e m bryo ns chez les fe melles parthéno génétiques (Fig ure 6). L es ovarioles des puce rons s ont polytro phiques, a ins i les ovocytes sont accompagnés de cellules nourricières. Chez ces d eu x m orp hes, les cellules ge rminales à la base de chaque ovariole form e nt un kyste sp hérique, ap pelé germ arium , constitué d e 3 2 ce llules ovogoniales interconnectées. C es 32 cellules ovogoniales se d ifférencient soit e n ovocyte, soit en cellule nourricière, avec un ra pport de un pour un [Black man, 1978].
En plus du type d ’ovocytes prod uits par ces d eux mo rphes, une a utre différence se situe au niveau des cellules nourricières, p uis que chez les femelles sexuées , le noyau de ces cellules augm e nte considéra blem e nt de taille par endomitose pe rm ettant ens uite la prod uction d’œufs non fertilisés, ce qui n’est pas le cas chez les femelles ase xuées. E n plus de cette différe nce m aje ure, l’a ppareil rep rod ucteur d es fem elles sexuées com porte une spermathèque perm ettant d e stocker la sem ence des mâles pour la féco nda tion des ovocytes haploïd e s, et ains i d e form e r d es œufs fécondés p rêts à l’oviposition.
D ans le cas de l’ovogénèse des fem elles sexuées, les futurs ovocytes entrent e n méiose I a u sein d u germarium , et sont donc haploïdes en fin de m éiose. Les ovocytes ainsi formés restent bloqués en méta phas e I avant d ’e ntre r en phase d e croissance, perm etta nt l’accum ulation de vitellus da ns leu r cytoplasm e grâce aux cellules nourricières. Les chromosom es restent co nd ensés jus q u’à la fécondation d es œ ufs par les spermatozoïd es localisés dans la sperm athèq ue lors du pas sage de l’ovocyte haploïde dans l’oviducte. Ces œ ufs fécondés sont ens uite d éposés s ur la plante hôte (oviposition). D ans les jours suivant la ponte , l’oe uf se mélanis e par la form ation d’une cuticule sérosale som bre. La bande germinale d e l’em bryo n s’invagine dans le vitellus deux à trois jours après la ponte, p uis l’inversion d e s axes ainsi que la segmentation com plète de l’em bryon s ont term inés 15 jours après l’oviposition [Miura et al., 2003]. L’œ uf re ntre ens uite e n dia paus e pe ndant 90 jo urs durant lesquels l’e mbryo n se développe d e m a nière rale ntie [S hingleton et al., 2003].
II. Synthèses b ibliographiq ues A. Biologie de s puce rons
Fig ure 6 | Appa reil rep rod ucte ur d es puce rons fem elles . A) Appareil re pro ducte ur de fe melles asexuées. Après expulsion dep uis les germ aria, les ovocytes diploïdes s e différe ncient e n em bryons . B ) Appareil re pro ducte ur de fe melles sexuées . Après expulsion d ep uis les g e rmaria , les ovocytes haploïdes se différe ncie nt e n œ ufs no n fertilisés par accum ulation de vitellus.
II. Synthèses b ibliographiq ues A. Biologie de s puce rons
Concerna nt l’e m b ryogénès e chez les femelles asexuées, les futurs ovocytes situés dans le germ arium sont bloqués e n prop hase et so nt ensuite expuls és du germ arium da ns une chambre folliculaire. Ils y subissent une m éiose II m odifiée ne présenta nt p as de phas e de réd uction chromosomom iq ue caractéristique de la m éios e I, permetta nt la form ation d’ovocytes diploïdes. Les chromosom es de ces ovocytes sont ensuite ra pide me nt condens és. Un e seule division d e maturatio n a alors lieu, ayant pour effet la libération d’un glob ule polaire. L’ovocyte diploïde subit ens uite plusieurs divisions mitotiq ues initiant le d éve loppe m ent em bryo nnaire. Au sein de l’ovariole, d ès qu’un ovocyte est exp ulsé en de hors d u g e rmarium, un a utre ovocyte prend sa place , form a nt ainsi une chaine d’e m bryons en cours de développe me nt dont les em b ryons les plus développ és progressent au fur et à m esure vers l’orifice génital situé au nivea u de la partie postérie ure du p uceron. U ne fois un em bryon pa rvenu au d ernier stad e em bryonnaire, il est expuls é sur la plante hôte, et est alors considéré comm e une larv e de p re mier stad e (L1). Environ s ix em bryons sont produits par chac un d es 16 ovarioles , pe rm ettant ainsi la prod uction d’enviro n 100 e mbryo ns par une fe m elle asexuée ad ulte. Le dévelo pp em ent des e m bryons a été fine me nt cara ctérisé en 20 stades e mb ryonnaires [Miura et al., 2003]. D ès les pre miers s tades em bryonnaires, des cellules germ inales s e développe nt au sein d es em bryons. Ainsi, au sein d ’un e mère d e génération 1, se trouve des e m bryons d e génération 2 , et e ux-mêm es comporte nt des cellules germ inales ainsi q ue des ovocytes en cours de différenciation, consituant la génératio n 3 et montrant bie n le phénomène de téléscop age des générations caractéristique chez les p ucerons parthénogénétique s [Miura et al., 2003]. C eci a été dém ontré par l’utilisation d’a nticorps ciblant d eux protéines codées par des g ènes m a rqueurs des cellules germinales de la D rosophile et conservés chez le p uceron, à savoir vasa et nanos. C es anticorps marquent en effet d es cellules germ inales primordiales se trouvant au s ein d’e mbryo ns, et ce à partir du 5
èm e
s tade d e dévelo ppeme nt [Cha ng et al., 2004]. D urant les stades tardifs de d évelop pem e nt, à partir d u stade 16, les dive rses parties du corps comm e les m us cles, les ye ux e t les appendices s ont déjà observables. En fonction de la photopério de, les ovocytes form és au sein d’e m bryons peuvent soit correspondre à de futures fem elles parthénogénétiq ues, à de futures fem elles sexuées ou à de futurs m âles (Figure 7). Les ovocytes diploïdes à l’origine d ’individ us m âle s sont obtenus par élimination d ’u n chro m osom e X via un méca nism e encore mal caractérisé. Les cellules som atiq ues d es mâles comportent ainsi trois paires d’autoso m es et un se ul chromosom e X, tandis q ue les fem elles (sexuées ou asexuées ) comportent trois paires d ’a utosomes et d eux chro m osomes X par cellule som atique. Il exis te donc un déséquilibre du nom bre d’allèles as sociés au chrom osome X entre m âles et femelles, vis-à-vis des autosomes. D es disfonctionne me nts cellulaires pe uve nt survenir s uite à un tel déséq uilibre q u’il convient de contrebala ncer via des m éca nism es dits de com p ensation d e dose. Avant de rentre r plus dans le détail de ces m éca nism es , nous présenterons les spécificités du chrom osom e X des pucero ns.
II. Synthèses b ibliographiq ues A. Biologie de s puce rons
Fig ure 7 | Cellules germinales et ovocytes des différents m orp hes d’A. pisum e n relation avec le polyphénism e de rep rod uction et le chromosom e X. Les batonnets noirs correspondent aux trois autosom es tandis que les b âtonnets verts correspo nd ent au chromosome X. En photop ériod e longue , les femelles asexuées pro duise nt dans leurs ovaires des ovocytes diploïdes (futurs em bryo ns). En photop ériod e courte , les fem e lles asex uées p rod uisent dans leurs ovaires des ovocytes diploïdes (em bryo ns) d e fe melles s exuées et de m âles. L es femelles sexuées , m êm e à l’état d ’e m b ryons , com porte nt des lignées ge rm inales prod uisant d es gam ètes ha ploïd es (œ ufs). Les pucero ns mâles sont obtenus par élimination d’un chromosom e X lors de la formation de le ur ovocyte diploïde. Les mâles com portent d es germaria pro duisa nt des ga mètes haploïd es (s permatozoïdes ).
II. Synthèses b ibliographiq ues A. Biologie de s puce rons
4. Le chro mosom e X chez les puce rons : cytologie et com pens ation
Au cours du siècle dernier, de nombre uses es pèces d e pucerons ont été karyotypées , montra nt une grande variabilité d u nom bre et d e la morp hologie de leurs chro mosom es de ma nière intra et inters pécifiq ue [K uznetsova et Shaposhnikov, 1973 ; Black ma n, 1980]. Les puce rons possède nt des chrom osom es holocentriq ues q ui ne présentent do nc pas de vérita bles ce ntromères, comportant ainsi une activité transcrip tomiq ue le long de l’ens e mble de leu rs chromosom es [Blackm a n, 1987]. Alors que chez les organis mes à chrom osom es m onocentriq ues la position d u centromère est une caractéristique d'ide ntific ation importa nte, l'a bsence d e centro mère des chromoso m es holocentriques chez les pucerons les re ndent difficilem ent ide ntifiables s’ils prése ntent d es tailles similaires. Par conséquent, la cytogénétiq ue chez les p ucerons s’est longte m ps limitée au compta ge et au tri des chromosomes s elon leur taille, à l'ide ntification des chrom osomes X grâce à la différence du nom bre de ce chro m osom e entre mâles et fe m elles , et à la localisation de la région orga nisatrice d u nucléole (NO R) par marq uage à l'arge nt. Le NO R correspond à un clus ter de gènes codant po ur les ARN ribosomiq ues 18S, 5,8S et 28 S, et ces régio ns ont été étudiées che z les puce rons e n tant q ue marqueurs chro m osomiq ues [Novotná et al., 201 1]. Les puce rons s ont e n effet des esp èces hétéroga m étiq ues m âles d e type XX /X0, co mme par exem p le le nématod e Caenorhabditis elegans. Les puce rons m âles ne possèdent donc q u’un uniq ue chromosom e X et sont prod uits par d es fem elles parthéno génétiques X X (Figure 7). Au cours d e ce processus de pro duction d’indiv idus m âles, l'un d es chrom osom es X est éliminé afin de générer des ovocytes diploïdes co mpo rtant de ux sets d ’autoso m es et un seul ch romosom e X (Figure 7).
Alors que les autosomes se sépare nt de m anière indép enda nte, les deux chromosomes X sont ici reliés e ntre e ux p ar une e x trémité, contraire m e nt à une mitose ord inaire. Un des deux chromosom es X se sépare d e son complém e nt et sa chrom atide sœur ségrége dans les cellules filles [O rlando , 1974 ; Blackm a n et H ales, 1986]. Il a été d émo ntré q ue les ch romosom es X d e pro méta p hase sont reliés pa r un étroit pont AD N-protéine q ui est forte me nt m a rq ué a u nitrate d 'arg ent et q ui corresp o nd rait au NO R [Mandrioli et al., 1999]. Par conséquent, le NO R est supposé jouer un rôle dans la ség régation des chro m osomes X dans les ovocytes destinés à donner des individ us mâles. Il est égale me nt importa nt d e noter qu e chez les p ucero ns, le chro mosom e X ten d à rep rése nter une propo rtion importa nte du génom e co mm e c’est le cas che z D iuraphis no xia (35%) [Nov otnáet al., 2011] et Acyrthosipho n pisum (31%) [Richard et al., 2017 ; Jaq uiery et al., 2017].
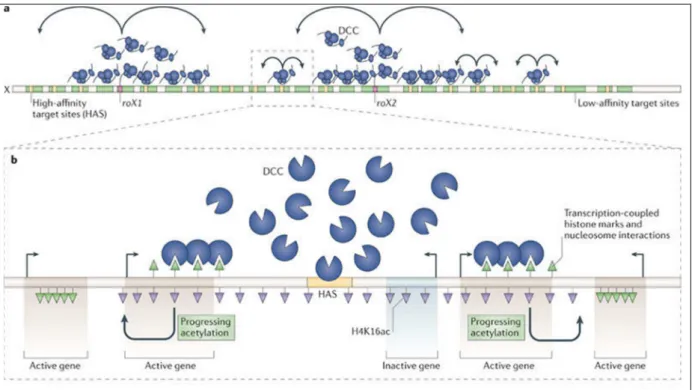
T out comm e c hez de nom b reux orga nism es h étéro ga m étiques mâles , il e xiste un d éséq uilib re dans le nom bre d’allèles ass ociés au chrom osom e X entre mâles et fem elles, induisant d e ce fait un différe ntiel d u nivea u d’expression d es gènes liés au X . Au co urs de l’évolution, certaines espèce s hétéroga m étiq ues ont acq uis des m éca nism es de comp ensation de dose (détaillés en II.C.) permetta nt d’ass urer un nivea u égal de transcription des chromo s omes X de chaque sexe com parative me nt aux autosom es. Ils impliq uent généralem ent d es m écanis mes épigénétiq ues , augm e ntant ou dim inuant l’accessibilité d e la chro m a tine ainsi q ue l’expression des gènes du chromoso m e X chez les m âles ou les fem elles. Ces m écanis mes ont notam me nt été étudiés chez certains orga nis mes m o dèles, do nt la D rosophile chez q ui un c omple xe protéiq ue (le D CC, ou com plexe d e com pe nsation d e dose )
II. Synthèses b ibliographiq ues A. Biologie de s puce rons
orchestre la co mpensation de dose chez les mâles [Conrad et Ak htar, 2012]. Afin d’étudier la prés ence potentielle de te ls m éca nismes chez les p ucero ns, des étud es visant à caractériser les profils d’e xpression d u chro mosom e X entre mâle et fe melles (sexuée et asexuée) ont été effectuées [Jaquiéry et al., 2013 ; Pal et Vicoso, 2015 ; Richard et al., 2017] (voir V.). Il a été identifié q ue le se ul chrom osom e X des mâles est en moye nne plus ex primé q ue les d eux chro m osome X des fem elles sexuées et asexuées, témoignant d’un potentiel méca nisme de com pe nsation de dose sous-jacent, ou tout du moins d ’u n méca nism e global d e rég ulation de la transcription d u X chez un des de ux s exes [Jaquiéry et al., 201 3].
Ces problèmes de déséq uilib re du nom bre d’allèle s associés au chromosom e X entre m âles et femelles découle d u méca nisme perm ettant la pro d uction des mâles par élimination d ’un d es deux chrom osom es X. Ceci s’effectue au cours du basculem ent d u deve nir d es em bryo ns et de leur lignée germinale dans le cadre d u polyphénis me de re prod uction (Figure 7). Les mécanismes moléculaires découla nt de la p erce ptio n de la diminutio n de la photop ériod e et p ermetta nt le bas culem ent d u deve nir des e mb ryons ont été étudiés chez les p ucerons, et principale me nt chez le puceron d u pois Acyrthosipho n pisum. Nous allons à présent décrire ces m écanism es m oléculaires .
5. Base s m oléculaire s du p olyp hénis m e d e reprod uction
Le polyp hénisme de re production chez le puce ron peut se déco mposer en plus ie urs p has es. La pre mière p hase consiste en une p erce ptio n du signal photop ériodiq ue et plus précisém e nt de la longueur de la nuit (sco tophase ) [Lees, 1973]. Le mécanism e précis de l’horloge circadienne permettant potentielle me nt une mesure de la photop ériod e et de la scotophase es t encore inconnu chez les puce rons, mais il s em ble q ue les gènes tim eless (tim) et perio d (per) y soient im pliq ués chez A. pisum puis que leur expre ss ion est co rrélée positiveme nt et oscille de m anière rob uste avec la
luminosité sur une périod e de 24 heures [Barberà et al., 2017]. Ces deux gènes sont des acteurs maje urs de l’horloge circad ienne chez D . melanogas ter en form ant un hétérodim ère T IM/PER dont la concentratio n est corrélée avec un rythm e circadien de 24 heures [G eka kis et a l., 1995 ; Z eng et al., 1996].
La température influe éga lem ent sur le bas culem e nt du mode de repro duction en accentuant le polyphénis me de re prod uction lorsque les tem pératures sont moyennes (15-20° C) comparativ em ent aux faibles tem pératures (11° C ) ou aux tem pérature s trop élevées (25° C ), ces dernières sup primant com plète m ent le basculem ent du mod e de repro duction [Lees, 1959 ; Chapma n, 1998]. Cep endant le signal photop ériodiq ue – en conditions contrôlées – est le seul param ètre néce ssaire et suffisant à l’initiatio n du polyphénis me de reprod uction. Le décle nchem ent de ce processus s’effectue selon de ux critères : i) une longue ur de nuit critique (CNL), consistant e n une durée m inim um d’obsc urité ininterro mp ue, et ii) un nombre de jours minim um consécutifs (« req uired day numb er », ou RD N) com portant cette CNL. Par exe m ple, chez Megoura viciae, la C NL est d e 7 he ure s et la RD N est de 5 jours à 15° C [Vaz N unes M et Hardie , 2000]. Les durée s respectives d e ces d eux facteurs varie nt d ’une espèce –voire d’un clone –de puceron à l’autre . D e plus , ce s param ètre s dépendent de la tem pérature, avec par exe mple chez Mego ura viciae des CNL de 6h, 7h ou 8h chez des femelles
II. Synthèses b ibliographiq ues A. Biologie de s puce rons
asexuées placées resp ectivem e nt à 12.5° C , 15° C ou 17.5° C, et une RD N de 5 jours ou 4.5 jours à 15° C et 17.5° C , res pectivem ent [ Vaz Nunes M et H ardie, 2000].
Plusieurs études ont perm is de mie ux com pre nd re le s mécanism es cellulaires e t moléculaires sous-jacents au polyphénis m e d e re prod uction, en com m e nç ant pa r s’intéress er à la pre m ière phase du polyphénis me de re prod uction : la perception et la tra nsductio n d u sig nal p hotop ériodique aux tiss us cibles (Figure 8). Le sig nal photopériodique est tout d’abord p erç u par le cerv eau d u p ucero n (les photorécepte urs photop ériodiques ne sont pas connus) puis il est trans m is aux tissus cibles, les ovarioles de la femelle ase xuée. Le g roupe d e cellule s neuro -sécrétrices I (NSC G roup I) se mble être impliqué da ns la perce ptio n du sig nal p hotop ériodique ou da ns la transm ission de ce signal ve rs les tiss us cibles, car une fois le NSC G roup I micro-cau térisé, il s ’est avéré q ue les pucero ns étaient incapables de répondre aux change m e nts photopériodiqu es [Steel et Lees, 1977]. La nature de la molécule permetta nt la transductio n du signal photo pério dique est encore inco nnue, m ais il est fait l’hypothèse q u’elle po urrait circuler par les projections axonales des cellules d u NSC G roup I jusq u’aux ovarioles où des réce pteurs pourraie nt être localisés. Une étude compa rant les transcripto mes de têtes d’A. pisum placés en conditions de jours co urts ou jours longs a perm is d’ide ntifier enviro n 600 transcrits différe ntielle me nt exprim és [Le T rionnaire et al., 2007]. Parmi ces transcrits se trouvent des gènes co dant des protéine s impliquées dans la modification de la structure de la cuticule, dans la signalisation nerve use et dans la signalisation neuro-endocrine, ainsi q ue des enzymes im pliquées dans les voies de signalisation de la dopam ine et de l’insuline [Le T rionnaire et al., 2009]. En outre, il a été m ontré q ue le génom e du puce ron contie nt 10 g ènes codant l’insuline , annotés Insuline Related Protein (IRP) 1 à 10, ce s dernie rs étant particulièrem e nt exp rim és au niveau d e la tête de M. persicae et A. pisum [H uyb rechts et al., 2010].
L’horm one juvénile est égalem ent im pliq uée dans la signalisation de la photopériod e. L’app lication ectopique d e K ino prène, un analogue d e l’ho rmo ne juvénile, sur la cuticule de puce rons placés en jours courts pro duisa nt do nc norm ale me nt des individus sexués , réve rs e le deve nir d es em bryo ns, ces derniers se dévelo ppa nt a lors en individ us asexués [Corbitt et Hardie , 1985 ; Hardie, 1987]. Ceci sugg ère une im plication d e l’hormone juvénile da ns la transduction d u signal photop ériodique. La mélatonine s erait aussi impliquée d e m a nière potentie lle da ns la transd uction du sig nal photop ériodique, puisque les gènes arylalkylam ine N-acetyltransferas e (AANAT ) 1 et 3 ont été identifiés comm e surexprimés au nivea u de têtes de pucerons placés en jours courts par rapport à des têtes de p ucero ns placés en jours longs chez A. pisum [Barb erà et al., 2013].
II. Synthèses b ibliographiq ues A. Biologie de s puce rons
Fig ure 8 | Modèle hypothétiq ue pour la rég ulation d u polyphénism e de rep rod uction chez les p uce rons. D es analyses transcripto miq ues à l’échelle d u génome co mbinées à des études physiologiq ues ont permis d'ide ntifier d es progra m me s génétiques pouvant joue r un rôle clé dans la rég ulation de la réponse à la photop ériod e chez les p ucerons. L es étap es initiale s de d étection et de transd uction d u signal p hotopériodiq ue se m blent ass ociées à une modification de la structure d e la cuticule, pouvant être liée à une réd uction d es nivea ux de do pamine dans les têtes des pucerons. Les systèmes nerve ux visuels et cérébra ux pourraie nt égalem e nt joue r un rôle d ans ce tte éta pe d e signalisation. L ’horm o ne juvénile a égalem ent été m ontrée com me impliquée dans la transduction endocrine de ce signal d u cerveau aux tissus cibles, les embryo ns. Les étapes ultérie ures à ce tte signalisation endocrine au niveau des e mbryo ns correspondent à un pass age d’une ovogénès e sexuée à une ovogénès e asexuée en condition d e p hotopériode courte. C ette modifica tion du deve nir des cellules germinales est accompag née d’une e xpression différentielle d e tra nscrits impliq ués da ns l’ovogénèse, dans le deve nir de la lignée germ inale, d ans des régulations transcriptionnelles et post-transcriptio nnelles, ainsi q ue dans des régulatio ns épig énétiq ues. D ’après [Le T rionnaire et al., 201 3].