Les Mycotoxines du mil : occurrence et flore fongique
259
0
0
Texte intégral
Figure

+7
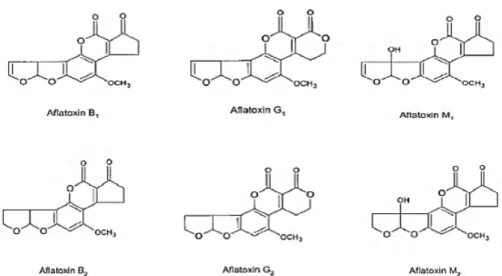

Documents relatifs