Régulation des variants d'épissage du cotransporteur rénal Na+-K+-Cl- de type 2 (NKCC2) : implication de la voie des kinases with no lysine (WNK)
188
0
0
Texte intégral
Figure
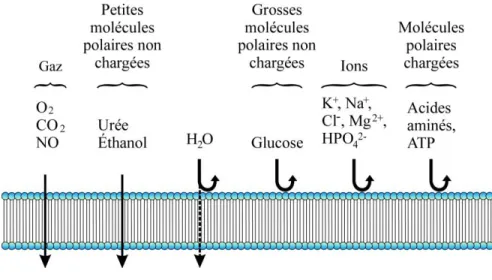
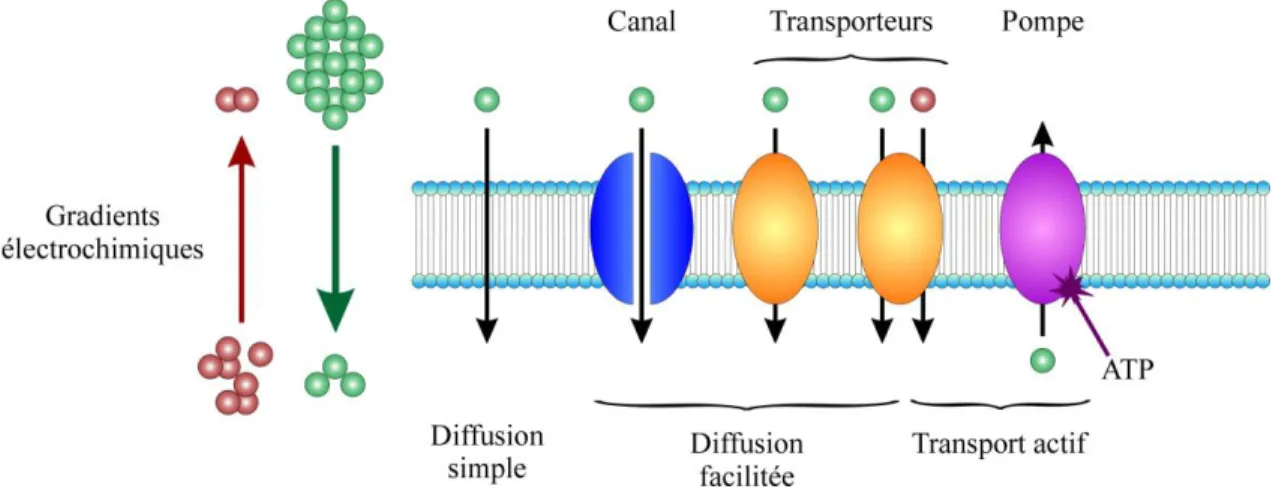
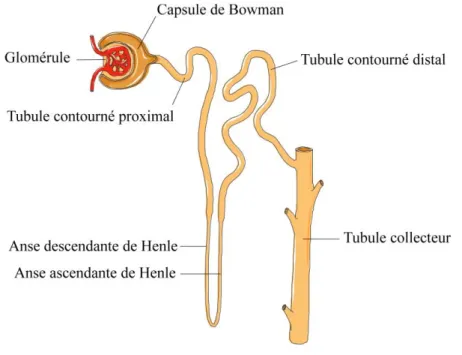

+7
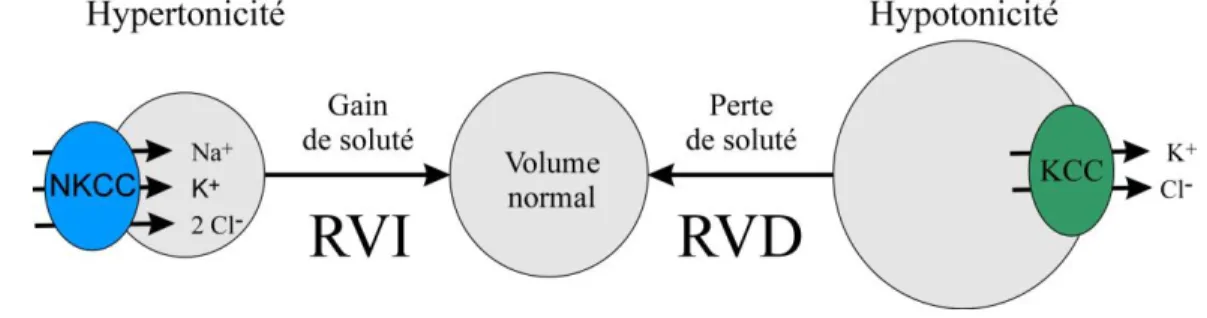


![Figure 1.8. Modèle d’activation de NKCC2. La diminution de la [Cl − ] i , le stress hypertonique et d’autres agonistes engendrent l’autophosphorylation des kinases WNK, la phosphorylation de SPAK/OSR1 puis de NKCC2](https://thumb-eu.123doks.com/thumbv2/123doknet/3184577.90926/39.918.325.629.177.505/modèle-activation-diminution-hypertonique-agonistes-engendrent-autophosphorylation-phosphorylation.webp)


Documents relatifs