N° d’ordre : 2743 THÈSE DE DOCTORAT Présentée par : Mouna FAHR Discipline : Biologie
Spécialité : Physiologie et Biotechnologie Végétale Titre :
Étude physiologique et moléculaire de la tolérance et l’accumulation du Pb chez une population métallicole et une population non métallicole
d’Hirschfeldia incana (Brassicacée récoltée au Maroc) : Perspectives pour la phytoremédiation
Soutenue le : 08 Décembre 2014 Devant le jury
Président
Pr Najib BENDAOU PES, Faculté des Sciences
Université Mohammed V, Rabat Examinateurs
Dr Didier BOGUSZ Directeur de Recherche IRD, Montpellier France
Pr Abdelaziz SMOUNI PA, Faculté des Sciences
Université Mohammed V, Rabat
Pr Rachida HASSIKOU PES, Faculté des Sciences
Université Mohammed V, Rabat
Pr Mohamed ZOUINE Maitre de conférences (HDR)
Dr Mohammed EL MZIBRI
INRA-INPT - Toulouse, France
Chef de l’Unité de Biologie et de Recherche Médicale -CNESTEN- Rabat
Pr Ali BOULARBAH PES, Faculté des Sciences et Techniques
Pr Mondher BOUZAYEN
Université Cadi Ayyad – Marrakech Directeur de Laboratoire Génomique et biotechnologie des fruits. INRA/INPT-ENSAT - Toulouse, France.
FACULTÉ DES SCIENCES
Rabat
2
i
Dédicaces
A l’aide de Dieu tout puissant, qui m’a tracé le chemin de ma vie, j’ai pu réaliser
ce travail que je dédie :
À mon Défunt père qui ne peut partager avec moi ces moments
exceptionnels de ma vie. J’espère que tu es fier de moi.
Tu étais et tu resteras
toujours présent dans mon cœur !
À ma très chère Maman, que j’aime beaucoup, qui n’a cessé de me
combler par son amour et sa tendresse. Puisse Dieu te donner santé, prospérité
et longue vie !
À mon cher frère Brahim qui m’a soutenue et encouragée depuis toujours,
dans mes études et dans la vie en général. Je t’aime et je te souhaite une longue
et heureuse vie !
À ma chère sœur Khadija qui, malgré les distances qui nous séparent,
était toujours proche de moi et suit le déroulement de ma vie jour pour jour. Je
t’aime et je te souhaite le bonheur dans ta vie !
À mon cher frère Mohamed qui m’a beaucoup encouragée et soutenue
aussi bien moralement que matériellement. Merci pour le rire et la joie que tu as
semé dans ma vie. Je t’aime et je te souhaite une longue vie pleine de joie et de
bonheur !
À ma cousine, ma deuxième sœur et mon amie Souad, qui était toujours
présente à mes côtés. Je t’aime et je te souhaite le bonheur dans ta vie !
À mon beau frère Brahim Fillali qui m’a soutenu et encouragé durant ma
thèse. Je te souhaite plein de succès et de joie !
À mes petits neveux que j’aime beaucoup: Adam, Yahya et Adam.
À tous les membres de ma famille sans aucune exception.
ii
Remerciements
Les travaux présentés dans ce manuscrit de thèse ont été réalisés dans le cadre d’une collaboration tripartite entre le Laboratoire de Physiologie et Biotechnologie végétale de la Faculté des Sciences de Rabat sous la direction de Pr Najib BENDAOU, le Laboratoire des biotechnologies des plantes du Centre National de l'Energie, des sciences et des Techniques Nucléaires de Rabat (CNESTEN) sous la direction de Pr Abdelaziz SMOUNI et l’équipe Rhizogenèse de l’UMR DIADE, Institut de Recherche pour le Développement IRD à Montpellier, France sous la direction de Dr Didier BOGUSZ, avec le soutien financier de l’AIRD-France et du Ministère de l’Enseignement Supérieure, de la Recherche Scientifique et de la Formation des Cadres du Maroc.
Je présente mes sincères remerciements à mon directeur de thèse Pr Najib BENDAOU, Professeur de l’Enseignement Supérieur, chef de département de Biologie et chef du laboratoire de physiologie et biotechnologie végétales de la Faculté des Sciences de Rabat, pour l’aide et le support moral qu’il m’a apportés tout au long de ce travail et pour sa gentillesse remarquable. Je le remercie également de m’avoir fait confiance depuis l’année 2006 en m’acceptant en Master « Production végétale » et en veillant à ce que je continuerai aussi bien mon projet de master que celui de thèse dans des très bonnes conditions.
Je tiens à remercier spécialement mon directeur de thèse français Dr Didier BOGUSZ (directeur de recherche à l’IRD). Je vous remercie d'avoir cru en mes capacités, pour le temps et la patience que vous m’avez accordés tout au long de ces années en me fournissant d'excellentes conditions logistiques. Vous étiez pour moi le professeur, le père et l’ami. Je garderai dans mon cœur votre générosité, votre compréhension ainsi que vos qualités humaines. Je n’oublierai jamais mes séjours à Montpellier grâce à vous (nos balades aux Cévennes, les matchs de foot et de Handball auxquels nous avons assisté…). Pour tout ce que vous m'avez donné, je vous remercie très sincèrement. Je vous remercie d’être ici présent parmi les membres de mon jury.
Je remercie mon cher co-encadrant Pr Abdelaziz SMOUNI (Professeur assistant à la Faculté des sciences Rabat) pour son encadrement instructif, ses conseils professionnels, ses orientations pertinentes et sa bienveillance académique exemplaire. Sans votre aide, je n’aurai jamais dû continuer ma thèse et surtout dans des très bonnes conditions. Mon séjour dans votre laboratoire depuis 2007 m’a donné la possibilité de mettre en pratique mes connaissances acquises et d’atteindre les compétences dans mon domaine de spécialité. Je suis aussi reconnaissante pour le temps qu’il a consacré à la correction de ce manuscrit. Je vous remercie infiniment d’avoir accepté de faire parti de mon jury de thèse.
Je remercie infiniment Pr Rachida HASSIKOU Professeur de l’Enseignement Supérieur à la Faculté des Sciences de Rabat d’avoir accepté d’être examinatrice de ce travail de thèse et de faire partie de mon jury.
J’exprime ma gratitude envers Dr Mohamed EL MZIBRI, Chef de l’Unité de Biologie et Recherche Médicale (UBRM), au CNESTEN, à Rabat, je le remercie d’être ici présent parmi les membres de mon jury.
Je le remercie également de m’avoir accordé l’opportunité d’intégrer le laboratoire de Biologie Moléculaire.
Mes remerciements se dirigent vers Dr Mohamed ZOUINE, Maitre de conférences de l’INPT-ENSAT de Toulouse, d’avoir aimablement accepté d’être rapporteur de ce travail de thèse.iii J’adresse également mes sincères remerciements à Pr BOULARBAH, Professeur d’Enseignement supérieure à la Faculté des Sciences et Technique- Université Cadi Ayyad Marrakech, d’avoir aimablement accepté d’être rapporteur de ce travail de thèse.
Je remercie Pr Mondher BOUZAYEN, Directeur de Laboratoire Génomique et biotechnologie des fruits de l’INRA/INPT-ENSAT - Toulouse, France, qui a accepté de juger ce travail en tant qu'examinateur. Je lui adresse mes sentiments les plus respectueux.
Mes profonds remerciements vont aussi au Dr. Claudine FRANCHE, directeur de recherche à l’institut de recherche pour le développement IRD-France de m’avoir gentiment accueilli parmi son équipe, et d’avoir mis à ma disposition tout les moyens pour mener à bien mon projet de thèse. Je la remercie pour sa gentillesse et son amitié.
Je tiens à remercier Dr. Valérie HOCHER, chercheur à l’IRD-France ; Dr. Patrick DOUMAS, chercheur à l’INRA- Montpellier France ; Dr. Laurent LAPLAZE chercheur à l’IRD-France ; Dr. Yves VIGOUROUX, chercheur à l’IRD-France pour leur aide sur certains aspect de ma thèse et pour leurs discussions toujours enrichissantes.
Mes remerciements aux enseignants du laboratoire de physiologie et biotechnologie végétale et également à tous mes professeurs de biologie végétale de la Faculté des Sciences Rabat.
J’exprime tous mes remerciements à l’ensemble de l’Equipe rhizogenèse, pour l’aide précieuse qu’ils m’ont apporté lors de mes stages à Montpellier.
Je remercie tout le personnel de laboratoire UBRM (CNESTEN) qui m’a prêté aide et assistance pour que je puisse effectuer mon stage dans de bonnes conditions. Qu’ils trouvent ici l’expression de ma profonde reconnaissance.
Un remerciement tout particulier au Pr Souad DEBBAGH (Enseignante chercheur à l’IAV Hassan II) qui m’a enseigné pour la première fois la biologie végétale lors de mes classes préparatoires de l’IAV Hassan II. Elle m’a donné envie de continuer dans le domaine de biologie végétale, sachant que j’avais une formation en mathématique durant le secondaire.
Un grand merci à mes chers collègues et amis de Montpellier : Julien, Issa, Philipe et Daniel. Toujours dans la bonne humeur, nous avons passé des bonnes moments ensemble et j’espère que nos routes se recroiseront un jour.
Ces remerciements ne seraient pas complets sans une pensée pour mes amies Zineb, Noama, Naima et Nawal avec qui j’ai passé des moments inoubliables aussi bien au labo (CNESTEN) qu’ailleurs. Je voudrais également remercier le comité scientifique de l’Institut de Recherche pour le Développement (IRD) France de m’avoir attribué une bourse qui a couvert financièrement trois années de ma période de doctorat (Jan 2011- Déc 2013).
iv
Productions scientifiques
A. Articles scientifiques internationaux
Fahr M., Laplaze L., Hocher V., Bendaou N., EL Mzibri M., Bogusz D., Smouni A. 2015. Assessment of lead tolerance and accumulation in metallicolous and non-metallicolous populations of Hirschfeldia incana. Environmental and Experimental Botany 109: 186–192 Fahr M., Laplaze L., Bendaou N., Hocher V., EL Mzibri M., Bogusz D., Smouni A. 2013. A review: Effect of lead on root growth. Frontiers in Plant Sciences 4: 175. doi: 10.3389/fpls.2013.00175
Auguy F., Fahr M., Moulin P., Brugel A., Laplaze L., El Mzibri M., Filali-Maltouf A., Doumas P. and Smouni A. 2013. Lead tolerance and accumulation in Hirschfeldia incana, a Mediterranean Brassicaceae from metalliferous mine spoils. PLoS One 8(5): e61932. doi: 10.1371/journal.pone.0061932.
B. Communications orales
Fahr M, EL Mamoun I, Laplaze L, EL Mzibri M, Doumas P, Triqui Z, Bendaou N, Hocher V, Bogusz D and Smouni A. Caractérisation et valorisation de plantes hyperaccumulatrice du plomb originaires du Maroc : perspectives pour la phytoremédiation des sites contaminés par les métaux lourds. Conférence Internationale ; Régler le cas des rejets miniers à travers de meilleurs pratiques de gestion. 5-7 Mai 2014. Marrakech, Maroc.
Fahr M, Auguy F, Vigouroux Y, Boguzs D, Laplaze L, Couderc M, Bendaou N, El Mzibri M, Doumas P et Smouni A : Diversity of lead hyperaccumulation in Hirshfeldia
incana : Perspectives for phytoremediation.. 9th International Phytotechnology Society from
11th to 14th September, 2012. Hasselt University in Belgium.
Smouni A, Fahr M, Auguy F, El Mzibri M, Doumas P : Caractérisation et valorisation de plantes hyperaccumulatrices du plomb originaires du Maroc : Perspectives en phytoremédiation. 14, 15 et 16 mai 2012- Montpellier- France.
Fahr M, Auguy F, Vigouroux Y, Couderc M, Moulin P, Laplaze L, Ater M, El Mzibri M, Doumas P et Smouni A : Etude physiologique et moléculaire de la diversité de l’hyperaccumulation du plomb chez Hirschfeldia incana - Perspectives en phytoremédiation. 3ème Colloque International de Biotechnologie Microbienne « Innovation technologique et valorisation des biomolécules » du 15 au 17 Mars 2012- Tanger-Maroc.
Fahr M, Auguy F, Vigouroux Y, Ater M, El Mzibri M, Laplaze L, Doumas P et Smouni A : Approches moléculaires de la diversité de l’hyperaccumulation du plomb chez
v
International, EAU, Déchets et Environnement (EDE3). 21-22 Novembre 2011, Faculté des sciences Dhar EL Mahraz, Fès- Maroc.
Fahr M, Auguy F, Vigouroux Y, Ater M, El Mzibri M, Laplaze L, Doumas P et Smouni A : Diversité de l’hyperaccumulation du plomb chez Hirschfeldia incana (Brassicacée) : Perspectives pour la phytoremédiation. Doctoriales de l'IRD France sud. 11 et 12 Octobre 2011 Montpellier, France.
AuguyF, Fahr M, BrugelA, LaplazeL, El Mzibri M, Filali-Maltouf M,Doumas Pand SmouniA: Hirschfeldia incana (Brassicaceae) is a lead hyperaccumulator and a good model to study Pb tolerance and accumulation. 8th APGC Symposium « Plant Functioning in a Changing Global and Polluted Environment», Groningen, The Netherlands, June 5 - 9, 2011.
C. Communications affichées
Fahr M, Auguy F, Vigouroux Y, Boguzs D, Laplaze L, Couderc M, Bendaou N, El Mzibri M, Doumas P et Smouni A : Diversity of lead hyperaccumulation in Hirshfeldia
incana : Perspectives for phytoremediation.. 9th International Phytotechnology Society from
vi
Résumé
Le plomb (Pb) est l’un des métaux les plus utilisés par l’homme mais aussi l’un des plus toxiques. Depuis plusieurs siècles, sa teneur n'a cessé d'augmenter en raison des activités anthropiques provoquant plusieurs problèmes environnementaux et sanitaires. Le Maroc présente plusieurs sites d’extraction du Pb qui sont des sources de contamination par ce métal. Sur le site d’Oued El Heimer, notre équipe a identifié plusieurs essences végétales tolérantes au Pb dont la brassicacée pseudométallophyte « Hirschfeldia incana ».
L’objectif de ce travail est d’étudier et de comprendre les mécanismes homéostasique qui permettent à H. incana de s’adapter à de fortes concentrations en Pb ce qui permettra d’optimiser son utilisation pour le développement de procédés de phytoremédiation de ce métal. Pour cela, nous avons réalisé une étude comparative des réponses physiologiques et moléculaires entre une population métallicole (PM) et une population non métallicole (PNM) d’H. incana. Une analyse génétique a également été réalisée en vue d’apprécier l’effet du stress métallique sur la spéciation de cette espèce.
La croissance, l'accumulation du Pb et plusieurs paramètres physiologiques ont été comparés entre la PM et la PNM d’H.incana cultivées en hydroponie à 0, 50 et 100 µM du Pb ou sur un sol pollué. Les résultats obtenus montrent une tolérance au Pb plus importante de la PM aussi bien en condition hydroponique que sur sol pollué. En hydroponie, les deux populations d’H. incana accumulent des concentrations élevées du Pb (>1 000mg/Kg de MS) dans leurs parties aériennes. L’accumulation du Pb est donc un caractère constitutif de cette espèce. Toutefois, la PNM accumule 2,5 fois plus que la PM. Chez la PM, le processus de photosynthèse est moins affecté par le Pb et la teneur en anthocyanes est significativement plus faible par rapport à la PNM. La modification des réponses physiologiques permettrait donc à la PM de mieux s’adapter aux fortes concentrations du Pb.
Le niveau d'expression des gènes HiMRP14, HiGCN2 et HiHMA4 a été évalué et comparé entre la PM et la PNM au niveau des parties aériennes par QPCR. Les gènes HiMRP14, HiGCN2 et HiHMA4 présentent une surexpression en réponse au Pb aussi bien chez la PM et la PNM. Ces résultats montrent qu’il y’a une régulation et une éventuelle implication de ces gènes dans l’accumulation du Pb au niveau des parties aériennes. Cependant la PNM exprime deux fois plus les gènes HiMRP14 et HiGCN2 que la PM d’H. incana. Ceci peut expliquer les différences d’accumulation du Pb entre les deux populations et suggère ainsi une possible adaptation de la PM par une régulation de l’accumulation du Pb.
Les gènes HiMT2, HiCCH, HiABC22 et HiGCN2 ont été choisis pour une étude génétique. L'analyse de séquences a montré une variation non significative au niveau intrapopulation. Tandis que la variation interpopulation est importante pour ces gènes. A ce niveau d’analyse, nous n’avons pas détecté d’effet de sélection sur les gènes chez les deux populations. Les différences d’adaptation au Pb observées pourraient être dues à une variation au niveau de la régulation des gènes étudiés.
Nos résultats démontrent que la PM est plus adaptée à tolérer et à accumuler le Pb et a donc un bon potentiel pour son utilisation dans des procédés de phytoremédiation des sites contaminés par le Pb.
Mots clés : Hirschfeldia incana, plomb (Pb), tolérance, accumulation, phytoremediation, population, métallicole.
vii
Abstract
Lead (Pb) is one of the most used metals but also one of the most toxic. For centuries, its content has been increasing due to human activities causing environmental and health problems. Morocco has several Pb extraction sites that are sources of contamination. On Oued El Heimer site, our team has identified several Pb tolerant plant species including the Brassicaceae and pseudométallophyte species "Hirschfeldia incana".
The main aim of this work is to study and understand the homeostatic mechanisms that allow adaptation of H. incana to high Pb concentrations. Thus, we carried out a comparative study of physiological and molecular responses between metallicolous population (PM) and a non metallicolous population (PNM) of H. incana. Genetic analysis was also performed to assess the effect of metal stress on the speciation of this species.
Growth, Pb accumulation and different physiological characteristics were compared in a MP and a NMP of H.incana grown in hydroponic at 0, 50 and 100µM of Pb or in polluted soil. Our results show that MP appears more tolerant than NMP in both hydroponic and soil culture. In hydroponic culture, the two populations of H. incana accumulate high Pb concentrations (> 1 000mg / kg DM) in their aboveground parts. Thus, the Pb accumulation is a constitutive character of this species. Nevertheless, Pb accumulation in the aboveground parts of NMP was 2.5 times higher than MP. The MP had significantly lower concentrations of anthocyanins and higher concentrations of chlorophyll a than NMP. Therefore, modification of physiological responses would allow a good adaptation of MP to high Pb concentrations.
MRP14, GCN2 and HMA4 genes expression was evaluated and compared between MP and NMP in the aboveground parts by QPCR analysis. The results showed that HMA4, GCN2 and MRP14 genes are regulated by Pb in both MP and NMP. This can suggest a functional role of the 3 genes in Pb accumulation. Interestingly, MRP14 and GCN2 genes were expressed constitutively higher in NMP. These results can explain the difference in accumulation between the two populations and suggest a possible adaptation of the MP by a regulation of Pb accumulation.
MT2, CCH, ABC22, GCN2 and HMA4 genes were selected for genetic comparison between the MP and NMP of H.incana. Sequences analysis didn’t show significant intrapopulation variation. While, interpopulation variation is significant for these genes. At this stage of analysis and based on these genes, we did not detect any effect of lead stress on the speciation of H. incana. The differences observed on Pb adaptation might be due to a variation at genes regulation level.
Our results demonstrate that the MP has adapted to tolerate and accumulate Pb and thus has a good potential for phytoremediation of Pb contaminated site.
Key words: Hirschfeldia incana, lead (Pb), tolerance, accumulation, phytoremediation, population, metallicoulous.
viii
Table des matières :
Dédicaces ... i
Remerciements ... ii
Productions scientifiques ... iv
Résumé ... vi
Abstract ... vii
Table des matières : ... viii
Liste des figures... xiii
Liste des tableaux ... xv
Liste des abréviations ... xvi
Introduction générale ... 1
Objectifs ... 5
Chapitre I : Revue bibliographique ... 7
I. Le plomb: propriétés physico-chimiques, effets sur la santé et l’environnement ... 8
1. Propriétés physico-chimiques du Pb ... 8
2. Origine naturelle du Pb ... 8
3. Sources anthropiques du plomb ... 9
4. Toxicité du Pb ... 10
II. Dépollution des sols contaminés par le plomb ... 11
1. Les techniques physico-chimiques ... 11
2. La bioremédiation ... 12
2.1. La bioremédiation par les microorganismes ... 12
2.2. La bioremédiation par les plantes « Phytoremédiation » ... 12
2.2.1. La phytostabilisation ... 13
2.2.2. La rhizofiltration ... 14
2.2.3. La phytoextraction ... 15
III. Le Plomb et la plante ... 16
1. Biodisponibilité du plomb pour les plantes ... 17
1.1. Effet de pH sur la biodisponibilité du Pb ... 18
1.2. Effet du potentiel redox sur la biodisponibilité du Pb ... 19
1.3. Effet de l’activité biologique sur la biodisponibilité du Pb ... 19
2. Le Pb et l’activité racinaire ... 20
2.1. Réponse racinaire au Pb ... 20
ix
3. Le Pb et l’activité de la partie aérienne des plantes ... 24
3.1. Assimilation du Pb par absorption foliaire ... 25
3.2. Assimilation du Pb par translocation depuis les racines vers les parties aériennes ... 25
3.3. Effet du Pb sur l’activité de la partie aérienne ... 26
4. Mécanismes de la tolérance et de l’accumulation du Pb ... 27
4.1. Absorption et transport des ions Pb2+ ... 27
4.1.1. Les chélateurs externes ... 28
4.1.2. Les chélateurs internes ... 28
4.2. Le transport transmembranaire ... 30
4.2.1. Les ATPases de types P ... 32
4.2.2. La famille des CNGC ... 32
4.2.3. Les transporteurs de type ZIP ... 32
4.2.4. Les transporteurs ABC ... 33
4.2.5. Les protéines NRAMP ... 33
4.2.6. Les antiports cation/H+ ... 34
Chapitre II: Site d’étude ... 45
I. Introduction ... 46
II. Cadre géographique de la région d'étude ... 47
III. Conditions climatiques et bioclimatiques de la région d'étude ... 47
IV. Description du site ... 48
V. Caractérisation du sol ... 50
1. Le pH ... 50
2. Evaluation de la pollution du sol aux ETMs ... 50
VI. Caractérisation floristique du site d’Oued El Heimer ... 51
1. La flore ... 51
2. Teneur en éléments-traces métalliques des plantes ... 52
3. Choix de la plante d’étude... 52
3.1. Description botanique de la plante « Hirschfeldia incana » ... 54
3.2. Cycle de vie ... 55
3.3. Origine et habitat ... 55
Chapitre III : Matériel & méthodes ... 56
A. Matériel ... 57
I. Matériel végétal ... 57
II. Echantillons de sol ... 57
x
B. Méthodes ... 57
I. Etude physiologique de la tolérance et l’accumulation du Pb des populations d’Hirschfeldia inacana ... 57
1. Culture des plantes ... 57
1.1. En condition hydroponique ... 58
1.2. En sol contaminé ... 58
2. Récolte et conditionnement du matériel ... 58
3. Extraction des anthocyanes ... 59
4. Extraction et dosage des chlorophylles ... 59
5. Dosage du Pb ... 60
5.1. Hydrolyse acide ... 60
5.1.1. Parties aériennes et racines ... 60
5.1.2. Sols ... 60
5.2. Détermination de la concentration en Pb ... 61
5.3. Analyse statistique ... 61
II. Analyse des séquences des gènes candidats ... 61
1. Extraction de l’ADN génomique ... 61
2. PCR (Polymerase Chain Reaction) ... 62
3. Clonage ... 63
4. Transformation d’E.coli ... 64
5. PCR sur colonies et purification de l’amplifiat ... 64
6. Electrophorèse sur gel d’agarose d’ADN ... 65
7. Séquençage de l’ADN ... 65
7.1. Principe ... 65
7.2. Mode opératoire ... 66
8. Purification des produits de réaction de séquence ... 68
8.1. Principe ... 69
8.2. Mode opératoire ... 69
9. Analyse des produits de séquençage ... 69
III. Analyse d’expression des gènes candidats par QPCR ... 70
1. Extraction d’ARNs ... 70
2. Traitement des ARNs à la DNase ... 70
3. Réaction de transcription inverse ... 71
4. Réaction de PCR quantitative (QPCR)... 71
xi
4.2. Mode opératoire ... 73
5. Electrophorèse en gel d’agarose ... 74
6. Analyse statistique ... 74
IV. Analyse bio-informatique ... 75
Chapitre IV : Etude physiologique de la tolérance et l’accumulation du plomb chez une population métallicole et une population non métallicole d’Hirschfeldia inacana en conditions contrôlées ... 76
I. Introduction ... 77
II. Résultats ... 79
1. Effet du Pb sur la croissance d’H. incana ... 79
1.1. Plante cultivées en hydroponie ... 79
1.2. Plante cultivées en sol pollué par le Pb ... 79
2. Accumulation du Pb chez la PM et la PNM ... 81
2.1. Plante cultivées en hydroponie ... 81
2.2. Plante cultivées en sol pollué par le Pb ... 81
3. Effet du Pb sur les concentrations d'anthocyanines ... 83
4. Effet du Pb sur les teneurs en pigments photosynthétiques ... 83
III. Discussion ... 85
1. H. incana est une pseudométallophyte qui tolère des concentrations élevées du Pb. ... 85
2. Chez H. incana, la PM est plus tolérante au Pb que la PNM. ... 86
3. L’accumulation du Pb est un caractère constitutif chez H. incana. ... 86
4. En conditions hydroponiques, la PNM d’H.incana accumule des teneurs plus élevées en Pb au niveau des parties aériennes par rapport à la PM. ... 87
5. La faible biodisponibilité du Pb en sol pollué affecte son accumulation chez H. incana. ... 88
6. Le Pb induit la synthèse et l’accumulation des anthocyanes au niveau les feuilles chez la PM et la PNM d’H.incana. ... 88
7. Le Pb affecte l'activité photosynthétique de la PNM d’H. incana. ... 89
IV. Conclusion ... 91
Chapitre V : Etude de l’expression des gènes HiMRP14, HiHMA4, HiGCN2 et HiMT2a chez la population métallicole et la population non métallicole d’H. incana ... 93
I. Introduction ... 94
II. Choix des gènes candidats ... 96
III. Résultats ... 99
1. Analyse de l’expression du gène HiMRP14 ... 100
2. Analyse de l’expression du gène HiGCN2 ... 101
3. Analyse de l’expression du gène HiHMA4 ... 102
xii 1. Le gène HiMRP14 pourrait être impliqué dans le stockage vacuolaire du Pb chez H. incana 103
2. Le gène HiGCN2 est plus surexprimé chez la PNM que chez la PM d’H. incana. ... 104
3. HiHMA4 est surexprimé en réponse au Pb chez les deux populations d’ H. incana. ... 104
V. Conclusion ... 105
Chapitre VI : Analyse intra et inter population des gènes HiABCG22, HiCCH, HiGCN2 et HiMT2a chez H. incana ... 107
I. Introduction ... 108
II. Choix des régions de gènes candidats ... 109
III. Résultats et discussion ... 112
1. Analyse intra-population des gènes candidats ... 112
2. Analyse inter-population des gènes candidats ... 114
IV. Conclusion ... 119
Conclusion générale & Perspectives ... 120
Références bibliographiques ... 125 Annexes ... 142 Annexe 1 ... 143 Annexe 2 ... 144 Annexe 3 ... 145 Annexe 4 ... 145 Annexe 5 ... 145 Annexe 6 ... 145 Articles ... Erreur ! Signet non défini.
xiii
Liste des figures
Figure 1 : Réserves mondiales en plomb en millions de tonnes (1990) (Laperche et al., 2004) ... 9
Figure 2 : Les principales de techniques de phytoremédiation (Moussavou, 2010). ... 13
Figure 3 : Interactions au niveau de la rhizosphère entre le sol, la plante et les microorganismes, influençant la phytodisponibilité des ETM (Pourrut, 2008). ... 18
Figure 4 : Schéma des différent réponses racinaires au Pb chez les plantes: (a) sequestration du Pb par les plaques Fe/Mn; (b) Fixation du Pb par la callose ; (c) l’efflux du Pb à travers la membrane plasmique ; (d) formation du complexe Pb-phytochélatine au niveau du cytoplasme ; (e) Transport du Pb-phytochelatine vers la vacuole (Fahr et al., 2013). ... 23
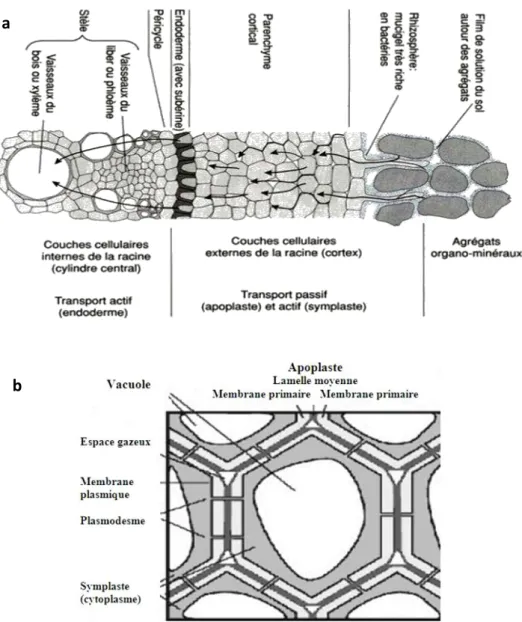
Figure 5 : Représentation schématique : (a) d’une coupe transversale de racine, indiquant le transport apoplastique et symplastique des ions, (b) des cellules végétales (Gobat et al., 1998) ... 31
Figure 6 : Schéma récapitulatif des transporteurs des métaux lourds dans une cellule végétale ... 35
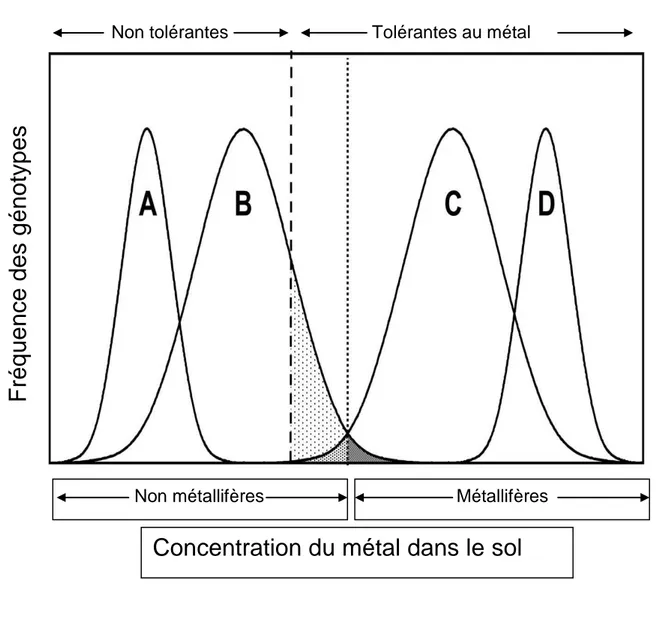
Figure 7 : Schéma de la répartition théorique des espèces végétale : A, B, C, D. Fréquence des populations métallicole (M) et non métallicole (NM) ainsi que la distribution (fréquence) des génotypes tolérants et non tolérants (Pollard et al., 2002). ... 38
Figure 8 : Représentation schématique des stratégies rencontrées chez les plantes en réponse à une augmentation des concentrations en métal dans le sol (Baker, 1981). ... 43
Figure 9 : Carte montrant la situation géographique de la région étudiée et la localisation des stations de prélèvements (Smouni et al., 2010). ... 47
Figure 10 : Description du site d’Oued El Heimer. A) vue d’ensemble de la fonderie et de la zone dénudée de végétation située dans le passage des émanations de gaz et de poussière émises par la cheminée ; B) scories plombifères déposées sur de grandes surfaces aux abords de la fonderie et du cours d’eau (Smouni et al., 2010). ... 49
Figure 11 : Site Oued el Heimer avec détail des émanations de gaz émises par la fonderie. Ces gaz sont essentiellement formés de SO2. ... 49
Figure 12 : Hirschfeldia incana poussant sur des scories plombifères du site d’Oued El Heimer. ... 55
Figure 13 : Etapes de la réaction de séquence ... 68
Figure 14 : Suivi en temps réel d'une réaction PCR. ... 73
Figure 15 : La biomasse racinaire (A), la biomasse des parties aériennes (B) des populations PM et PNM d’H. incana cultivées pendant 3 semaines à différentes concentrations du Pb et la biomasse des parties aériennes (C) des deux populations PM et PNM d’H. incana cultivées sur un sol pollué de Oued el Heimer pendant 40 jours. ... 80
Figure 16 : Concentrations du Pb au niveau des racines (A), des parties aériennes (B) de la PM et la PNM d’H. incana cultivées à différentes concentrations du Pb pendant 3 semaines et La concentration du Pb au niveau des parties aériennes (C) de la PM et la PNM d’H. incana cultivées sur un sol pollué d’Oued el Heimer pendant 40 jours... 82
xiv Figure 17 : Concentration d’anthocyane au niveau des feuilles de la PM et la PNM d’H. incana cultivées à différentes concentrations du Pb pendant 3 semaines. ... 83 Figure 18 : Concentration de chlorophylle a (A) et b (B) au niveau des feuilles de la PM et la PNM d’H. incana cultivées à différentes concentrations du Pb pendant 3 semaines. ... 84 Figure 19 : Expression relative du gène MRP14 mesurée par QPCR au niveau des parties aériennes (PA) d’H.incana chez la PM et la PNM. ... 100 Figure 20 : L’expression relative du gène HiGCN2 mesurée par QPCR au niveau des parties aériennes (PA) d’H.incana chez la PM et la PNM. ... 101 Figure 21 : L’expression relative du gène HiHMA4 mesurée par QPCR au niveau des parties aériennes (PA) d’H.incana chez la PM et la PNM. ... 102 Figure 22 : Résumé des blasts effectué sur le site www.tair.com sur les quatre gènes étudiés chez A. thaliana. ... 111 Figure 23 : Résultat d’alignement des séquences du gène HiABCG22 chez différent individus de la PM (bleu) et la PNM (rouge) d'H. incana. ... 116 Figure 24 : Résultat d’alignement des séquences du gène HiGCN2 chez différent individus de la PM (bleu) et la PNM (rouge) d'H. incana. ... 117 Figure 25 : Résultat d’alignement des séquences du gène HiMt2a chez différent individus de la PM (bleu) et la PNM (rouge) d'H. incana. ... 118 Figure 26 : Résultat d’alignement des séquences du gène HiCCH chez différent individus de la PM (bleu) et la PNM (rouge) d'H. incana. ... 118
xv
Liste des tableaux
Tableau 1 : Caractéristiques physico-chimiques du Pb ... 8
Tableau 2 : Les techniques utilisées pour la remédiation d’une contamination métallique du sol ... 12
Tableau 3 : Plantes proposées pour la rhizofiltration du Pb (Fahr et al., 2013) ... 15
Tableau 4 : Production minière et flux d’exportation du Plomb (2002) ... 46
Tableau 5 : Concentration en ETM (As, Cd, Cu, Ni, Pb et Zn) (mg/kg) et indice de pollution ... 51
Tableau 6 : Teneurs en ETM (en mg/Kg de MS) dans les feuilles des plantes prélevées sur les différents sites d’étude. Les teneurs en ETM ont été déterminées par ICP-AES. ... 52
Tableau 7 : Les métaux et les concentrations que tolère Hirschfeldia incana : Les concentrations totales et biodisponibles (DTPA extractible) en métaux lourds au niveau des sols contaminés. (Gisbert et al., 2006) ... 55
Tableau 8 : Amorces basées sur les séquences d’A. thaliana utilisées pour amplifier les gènes candidats chez H. incana. ... 62
Tableau 9 : Amorces spécifiques d’Hirschfeldia incana... 63
Tableau 10 : Mélange réactionnel pour le séquençage. ... 67
Tableau 11 : Amorces utilisées pour la Q-PCR à partir des ADNc d’H.incana. ... 73
Tableau 12 : Liste des gènes candidats chez H.incana, classés par catégorie fonctionnelle, au niveau des parties aériennes (PA) et des racines (R) (Auguy et al non publié). ... 97
Tableau 13 : Liste des gènes candidats... 99
Tableau 14 : Résultat de l’analyse intrapopulation : diversité nucléotidique et tests de neutralité pour les 4 gènes candidats au niveau de la PM et de la PNM d’H. incana. ... 113
xvi
Liste des abréviations
A : Absorbance
ABC : ATP-Binding Cassette ADN : Acide désoxyribonucléique ADNc : ADN complémentaire Ag : Argent
Alt : Altitude
ARN : Acide ribonucléique As : Arsenic
ATM : ABC transporter mitochondrial ATP : Adénosine tri-phosphate BD : Broughton and Dilworth Ca : Calcium
CAX : Cation exchangers
CBP : Calmodulin-Binding Protein CCH : Copper chaperone
Cd : Cadmium
CDTA : trans-1, 2-Cyclohexylene Dinitrilo Tetraacetic Acid Chl a : Chlorophylle a
Chl b : Chlorophylle b
CNGC : Cyclic nucleotide-gated ion channel Co : Cobalt
Cr : Chrome
CSH : Cross Species Hybridisation Cys : Cystéines
dNTP : désoxyribonucléotide DEPC : Diéthyl pyrocarboate
DTPA : Diethylene triamino pentaacetic acid
EDDHA : Ethylenediamine-N,N0bis(o-hydroxyphenyl) Acetic Acid EDTA : Acide éthylène-diamine-tétraacétique
EGTA : Ethylene bis[oxyethylenetrinitrilo] Tetraacetic Acid ERO : Espèces Réactives de l’Oxygène
ET :
ETMs : Eléments traces métalliques FBC : Facteur de bioconcentration FC : Fold change
Fe : Fer
GFP : Green Fluorescent Protein GCN : General Control Non-repressible GSH : Glutathion
GSHS : Glutathion Synthétase GST : Glutathion-S-Transférase H : hydrogène
HBED : N, N0-di(2-hydroybenzyl) Ethylene Diamine N, N0-diacetic acid HCl : Acide chlorhydrique
xvii Hi: Hirschfeldia incana
HMA : Heavy metal ATPase
ICP-AES : Inductively Coupled Plasma-Atomic Emission Spectrometry IP : Indice de Pollution
IRT : Iron-Regulated Transporter ISSR : Inter Simple Sequence Repeat ITS : Internal transcribed spacer K : Potassium
Mg : Magnésium Mn : Manganèse
MRP : Multidrug Resistance Protein MRE : Metal Response Element MS : Masse Sèche MT : Métallothionéine N : Nord NADPH : nicotinamide-adénine-dinucléotide-phosphate NE : Nord Est Ni : Nickel
NRAMP : Natural Resistance-Associated Macrophage Protein P : Phosphore
Pb : Plomb pb : Paire de base PbS : Galène PC : Phytochélatine
PCR : Polymerase chain reaction PDR : Pleitropic Drug Resistance
PGPR : Plant Growth Promoting Rhizobacteria pH : Potentiel d’hydrogène
PM : Population Métallicole PNM : Population Non Métallicole PTFE : Polytétrafluoréthylène
RAPD : Random Amplified Polymorphic DNA RFLP : Restrictio Fragment Length Polymorphism Rpm : Rotation par minute
RT : Reverse Transcription
RUBISCO : Ribulose-1,5-bisphosphate caboxylase QPCR : PCR quantitative
QTL : Quantitatif Trait Loci SOD : Superoxyde Dismutase TBE : Tris Borate EDTA Tm : Température de fusion UV : Ultraviolets
v/v : Volume/volume W : Ouest
ZIP : Zinc and Iron regulated transporters Protein Zn : Zinc
1
2
Introduction générale
Les éléments traces métalliques (ETMs) ou « métaux lourds » sont des composants essentiels des écosystèmes terrestres car ils participent aux cycles biogéochimiques. Ils sont pour certains d’entre eux essentiels au bon fonctionnement de la biosphère. A faible dose, certains métaux comme le fer, le zinc, le chrome, le cuivre,... sont indispensables à la survie des formes vivantes. Cependant, à forte dose, ils sont toxiques et peuvent être à l'origine de disfonctionnements physiologiques majeurs et de maladies graves (Borovik, 1990). Par ailleurs, d’autres métaux qui n’ont pas de fonctions biologiques, sont classés parmi les polluants dits "polluants atmosphériques dangereux". Il s'agit en particulier du plomb (Pb), du mercure (Hg), de l’arsenic (As) et du cadmium (Cd).
En absence d’activités humaines, les métaux lourds, par nature non biodégradables (Traunfeld et Clement, 2001) sont relâchés dans l’environnement au rythme de l’érosion ou des activités biologiques (Gadd, 2010). Cependant, ce rythme est considérablement perturbé par l’anthropisation. Depuis le siècle dernier, les activités industrielles ont connu un développement considérable et ont entrainé une accélération de l’industrie minière. Après l’arrêt des activités et la fermeture des sites miniers, la plupart des déchets sont laissés à l’abandon. Ces déblais fortement chargés en métaux lourds sont alors facilement érodés. Ces activités extractives très polluantes engendrent des perturbations des cycles biogéochimiques modifiant ainsi la dynamique et le fonctionnement des écosystèmes avec des conséquences parfois irréversibles sur la biodiversité et peuvent aller jusqu’à des changements climatiques (Sala et al., 2000).
Lorsque le risque pour les écosystèmes est très élevé, la dépollution s’impose. Il est important donc d’élaborer des stratégies efficaces de gestion des environnements pollués par les métaux. Une bonne gestion permet ainsi de limiter les effets délétères des matériaux issus d’anciennes activités minières et mieux prendre en compte les problèmes environnementaux et la santé publique. Dans ce contexte, plusieurs approches ont été mises en œuvre pour réhabiliter ces milieux pollués. La majorité de ces approches sont basées sur des procédés physico-chimiques (Pierzynski, 1997; Maenpaa et al., 2002 ; Mench et al., 2005). Ces techniques restent, malgré leur efficacité, très coûteuses et laborieuses avec un impact non négligeable sur l’environnement. Afin de pallier les principales limitations de ces méthodes, les recherches
3
s’orientent depuis quelques années vers l’utilisation des organismes vivants : des microorganismes ou des plantes (phytoremédiation), comme système biologique (Pilon-Smits, 2005).
Actuellement, le terme de phytoremédiation est largement accepté et regroupe toutes les méthodes basées sur l’utilisation de végétaux dans un but de minimiser l’effet néfaste des métaux sur l’environnement. Malgré leur développement récent, ces méthodes sont variées et des résultats encourageants ont d’ores et déjà été obtenus pour des problématiques diverses. Cependant, à l’heure actuelle, le principal obstacle au développement de la phytoremédiation réside dans le faible nombre d’espèces potentiellement utilisables à l’échelle industrielle, la faible biomasse des plantes utilisées, ainsi que le manque en connaissances des mécanismes physiologiques et du déterminisme génétique de la tolérance et de l’hyperaccumulation des métaux.
Certaines plantes, dites métallophytes, sont capables de se développer normalement sur les sites fortement pollués par divers métaux. Ces plantes métallophytes ont la particularité d’être soit métallophytes « strictes » : se développant uniquement sur les sites à fortes concentrations en métaux, soit des métallophytes facultative ou pseudométallophyte : se développant aussi bien sur des sites métallifères (population métallicole) que sur des sites non métallifères (population non métallicole). Ces populations présentent des différences au niveau physiologique et moléculaire qui leur permettent de s’adapter chacune à son environnement. Certains de ces végétaux, qualifiés d’hyperaccumulateurs (Brooks, 1998), sont capables de stocker massivement les métaux dans leurs parties aériennes (Remon, 2006).
Une des approches qui permettent d’étudier ces mécanismes et mieux comprendre les réponses des plantes vis-à-vis des métaux, est l’étude des populations des espèces pseudométallophytes, afin de cibler l’écotype le mieux adapté. Ces études consistent en une analyse phénotypique et physiologique puis au niveau génétique de la distribution des polymorphismes pour ces caractères. Il est donc essentiel de connaître le mieux possible les populations naturelles des espèces cibles afin d’en estimer les ressources génétiques pour être en mesure de les exploiter le plus efficacement possible tout en les conservant.
Contexte d’étude
Depuis le début de XXe siècle, la partie orientale du Maroc (région d’Oujda, Figuig et Er Rachidia) a été le siège d’importantes activités d’extraction et d’exploitation de métaux
4
lourds avec en tout premier lieu du plomb car ce métal était très demandé de part ses nombreuses et diverses applications. Le pic d’exploitation du Pb a eu lieu dans les années 80 où la production marocaine représentait jusqu’à 3,5 % de la production mondiale (Wadjini, 1998). Aujourd’hui, Cette exploitation se poursuit pour un certain nombre de ces sites alors que d’autres sont abandonnés depuis plusieurs décennies suite à l’épuisement des filons. En absence d’une politique d’après mine, ces sites représentent des cas exemplaires de pollution élevée en métaux lourds, notamment en Pb, zinc (Zn) et cadmium (Cd). Ces déblais miniers rejetés, très riches en métaux lourds toxiques, sont à l'origine de plusieurs problèmes environnementaux et sanitaires dont soufrent ces régions. De plus, en raison du couvert végétal épars et peu abondant et d’une érosion très active dans ces régions semi-arides à arides, les métaux sont disséminées par le ruissellement des eaux et par le vent vers les régions cultivées et les zones habitées avoisinantes.
Notre équipe mène des recherches, sur les sites miniers de la région d’Oujda, très fortement contaminés en métaux lourds en particulier le Pb (Smouni et al., 2010), en vue de développer des stratégies de phytoremédiation. Ces stratégies s’appuient sur l’utilisation des espèces végétales locales capables de se développer sur ces milieux afin de limiter la dispersion par voies aériennes ou hydriques des particules métalliques.
Malgré les conditions extrêmes, plusieurs essences ont été identifiées par notre équipe sur ces sites à forte contaminations en Pb. En particulier, il y a une dominance et une végétalisation naturelle importante par la brassicacée « Hirschfeldia incana ». H. incana tolère bien le Pb et l’accumule dans ses parties aériennes. L’accumulation du Pb chez H. incana a été également confirmée au laboratoire en conditions contrôlées. Cette plante est un modèle idéal pour l’étude moléculaire et physiologique de la tolérance au Pb chez les plantes. Il est à noter qu’à nos jours, aucune plante-modèle pour l’étude de la tolérance et l’accumulation du Pb n’a été décrite. H. incana présente l’intérêt d’appartenir à la famille des Brassicacées qui comprend plusieurs plantes modèles pour la tolérance aux métaux (Noccaea caerulescens pour le Cd et
Arabidopsis halleri pour le Zn). Cette proximité va nous permettre également d’utiliser les
données existantes chez le modèle Arabidopsis thaliana.
H. incana est une plante caractéristique de la région méditerranéenne avec une aire de
répartition très large. C’est une pseudométallophyte qui se développe aussi bien sur des sols pollués que sur des sites naturels non contaminés. Cette particularité fait de cet espèce un très bon modèle pour étudier les différences entre les deux types de populations, métallicole PM et
5
non métallicole PNM. Les différences entre populations M et NM permettent d’une part d’apprécier le degré de spéciation en réponse à la pression de sélection exercée par les métaux et d’identifier les locus impliqués dans la tolérance et l’adaptation aux fortes concentrations en métaux. D’autre part de choisir, chez une espèce, les écotypes les mieux appropriés pour l’élaboration de stratégies de phytoremédiation.
Ce projet de thèse a pour objectif d’étudier et de comparer la capacité des deux populations d’H.incana à tolérer et accumuler le plomb. Un autre objectif est d’identifier les bases moléculaires de la tolérance et de l’hyperaccumulation du Pb ainsi que d’explorer ce pouvoir accumulateur du Pb en choisissant la population la mieux adapté au Pb pour développer des procédés de phytoremediation.
Objectifs
La thèse se déclinera en trois parties complémentaires :
1- Caractérisation physiologique de la tolérance et l’accumulation du Pb chez des populations M et NM d’H. incana :
Le but est d’étudier la variabilité de la tolérance et de l’accumulation du Pb d’individus issus d’une population métallicole ou d’une population non métallicole d’H. incana provenant de différent sites : contaminés (site minier d’Oujda) ou non (zone agricole de Tetouan) par le Pb. Ainsi, les individus de chaque population ont été soumis à différentes concentrations du Pb soit sur un sol contaminé ou en condition hydroponique. Après traitement avec ou sans Pb, des individus issus de populations métallicoles (PM) et non métallicoles (PNM) d’H. incana, les différences en biomasse fraiche et le taux d’accumulation du Pb dans les parties aériennes et les racines ont été évalués. Les teneurs en certains pigments au niveau des feuilles ont été également exprimées.
2- Analyse d’expression des gènes candidats :
En s’appuyant sur les données de la littérature et les résultats d’une approche transcriptomique qui a été réalisée par notre équipe, nous avons identifié plusieurs gènes candidats pour l’étude des mécanismes moléculaires de la tolérance au Pb chez H incana. Les trois gènes étudiés codent pour des transporteurs : deux transporteurs membranaires HMA4 et GCN2 et un transporteur vacuolaire (du tonoplast) MRP14. Le niveau de régulation de ces gènes a été
6
mesurée dans les feuilles des plantes exposées ou pas au Pb et comparée entre les deux populations d’H. incana.
3- Analyse intra et inter population des séquences de gènes candidats impliqués dans la tolérance et l’accumulation des métaux chez les plantes.
Les gènes GCN2, ABC22, CCH et MT2a, qui ont été également identifiés par l’approche transcriptomique, ont été choisis pour cette étude. Les polynucléotides correspondant à chacun des gènes candidats (trois transporteurs et un chélateur) ont été isolés et séquencés sur une vingtaine d’individus différents d’une population métallicole (M) et d’une population non métallicole (NM). Une analyse intra et inter-population est réalisée. Ainsi, la comparaison des séquences et la détection de signature de sélection permettra d’identifier les gènes associés à ces phénomènes d’adaptation.
7
Chapitre I :
8
I. Le plomb: propriétés physico-chimiques, effets sur la santé et
l’environnement
1. Propriétés physico-chimiques du Pb
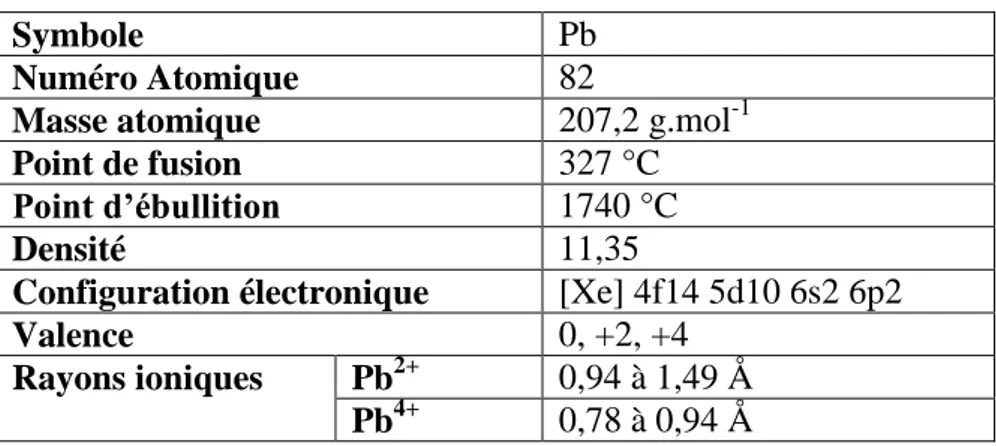
Le plomb (Pb) est connu depuis la haute antiquité. Son nom vient du latin plumbum signifiant liquide argenté. Parmi les métaux lourds dont le numéro atomique est supérieur à 60, le Pb est le plus abondant dans la croûte terrestre (Adriano, 2001). C’est un métal bleuté brillant, mou, très malléable et ductile. En raison de son caractère chalcophile, le Pb natif est rare. Il est associé au gisement de sulfures ; on l'extrait de sa source minérale principale, la galène (PbS) qui en contient 86.6% en poids (Cheng et Hu, 2010), mais aussi des minerais associés au zinc (Zn) (la sphalérite), à l'argent et le plus abondamment au cuivre (Cu) (Hurlbut et Klein, 1982). D'autres variétés communes sont la cérusite (PbCO3) et l’anglésite (PbSO4). Le tableau 1
résume les caractères physicochimiques du Pb.
Tableau 1 : Caractéristiques physico-chimiques du Pb
Symbole Pb
Numéro Atomique 82
Masse atomique 207,2 g.mol-1
Point de fusion 327 °C
Point d’ébullition 1740 °C
Densité 11,35
Configuration électronique [Xe] 4f14 5d10 6s2 6p2
Valence 0, +2, +4
Rayons ioniques Pb2+ 0,94 à 1,49 Å
Pb4+ 0,78 à 0,94 Å
2. Origine naturelle du Pb
Le Pb est présent de façon naturelle dans la croûte terrestre et dans tous les compartiments de la biosphère. Les gisements associés au volcanisme forment les plus grosses concentrations de sulfures massifs. Ces gisements du Pb sont en grand nombre et répartis dans le monde entier. Plus de 150 mines sont réparties dans 35 pays (Laperche et al., 2004). Les réserves dans le monde étaient de 130 millions de tonnes en 1990 (figure 1), alors qu’en 2011, le stock était estimé à 79 millions de tonnes dans les gisements exploitables (USGS, 2011). Dans l'air, les émissions du Pb provenant de poussières volcaniques véhiculées par le vent sont reconnues d'une importance mineure. D’autres processus naturels, comme la dégradation et l'érosion des
9
roches mères (Gadd, 2010) et les feux de forêt, contribuent de façon significative à la libération du Pb. Mais généralement, ces processus naturels ne conduisent que rarement à des concentrations élevées du Pb dans l'environnement.
Figure 1 : Réserves mondiales en plomb en millions de tonnes (1990) (Laperche et al., 2004)
3. Sources anthropiques du plomb
Les contaminations par le Pb sont généralement anthropiques. Elles sont à l’origine d’un flux du Pb très important pouvant conduire à des contaminations locales aiguës. Ce flux représentait, en 1980, 332 000 t.an-1 contre 12 000 t.an-1 d’origine naturelle (Nriagu and Pacyna, 1988).
L'histoire du Pb remonte à des temps très anciens. Il est beaucoup utilisé en raison de sa diffusion, de sa grande malléabilité et son bas point de fusion. Il était employé lors de l'âge du bronze avec l'antimoine et l'arsenic. Il a été utilisé abondamment dans diverses applications, principalement en raison de son excellente résistance à la corrosion dans l’environnement (ATSDR, 2007). Les éventuelles sources sont les pratiques agricoles due aux épandages d'insecticides, de fongicides, d'engrais, de boues résiduaires urbaines ou de composts d'ordures ménagères,…etc. De nombreux pesticides contiennent également des éléments métalliques ou métalloïdes tels que le Pb, Hg, Cu, Mn, Cd ou As (Atafar et al., 2010). Il y a également les retombées atmosphériques telle que l'industrie extractive et métallurgique, pratiques liées à l'incinération, utilisation des énergies fossiles, essence au Pb, etc.
10
Dans la plupart des cas, les contaminations de l’environnement trouvent leur origine dans des pratiques du passé (extractions et fonderies) qui ne tenaient pas compte des impacts environnementaux, que ce soit en matière d’exploitation industrielle ou minière (infiltration ou déversement de substances polluantes, stockage sans mesure de protection des sols, retombées de poussières…), ou en matière d’élimination des déchets (décharges, dépôts de résidus miniers ou industriels utilisés comme remblais…).
L’activité minière perturbe énormément l’environnement dès le début de l’exploration des sites avec la destruction de l’habitat naturel. Ensuite, l’exploitation se poursuit par l’extraction du minerai. Avant d'être transporté à la surface, le minerai abattu est soumis à un premier triage, en fonction de son poids (seuls les morceaux les plus lourds sont conservés, car cela signalait la présence du Pb argentifère, plus dense) et de sa couleur (la présence de grains noirs, caractéristiques de la galène pure, est discriminante). Ce qui est jugé exploitable est remonté à la surface, aux abords des mines dans les laveries, pour y subir plusieurs opérations de tri et de concassage en particules très fines puis l’enrichissement en utilisant, comme procédé, la gravimétrie et/ou la flottation (selon la taille des minéraux) qui utilise les différences d’hydrophobicité des métaux à séparer. Ces traitements génèrent des déchets qui représentent 88 à 99 % de la roche extraite et qui contiennent encore des quantités non négligeables du Pb (1 à 12%). Ils sont déposés en couches successives via des tuyaux et forment après assèchement de gigantesques digues (haldes) à proximité des sites d’exploitation.
4. Toxicité du Pb
Le Pb est un élément trace métallique non essentiel aux métabolismes des organismes vivants. Il est classé au premier rang des éléments les plus dangereux (Duker et al., 2005). Les sources d’exposition sont diverses telles que l’inhalation ou l’absorption du Pb dans des situations accidentelles. L’exposition au Pb constitue un risque pour la santé humaine. Une fois dans l’organisme, le Pb se concentre, via la circulation sanguine, dans le cerveau et les os. En cas d’intoxication chronique, le Pb induit le saturnisme. Il est réputé également par son pouvoir mutagène et cancérogène (Robert et al., 2007).
Par conséquent son accumulation dans l’environnement (sol, eau, air) présente un danger permanant pour les populations, si les mesures nécessaires ne sont pas prises par les autorités concernées. Dans des pays où la législation est inexistante, la pollution par le Pb va perdurer
11
pendant des centaines d’années si aucune action n’est envisagée. Ainsi les populations les plus fragiles courent le risque d’être exposées à des quantités importantes de Pb, d’où la nécessité de restaurer et dépolluer ces sites.
II. Dépollution des sols contaminés par le plomb
La réhabilitation des sites pollués représentent aujourd'hui un des enjeux majeurs des états, dans le cadre de développement durable. D’où l’orientation des recherches, ces dernières décennies, au développement de techniques de dépollution de plus en plus efficaces et écologiques.
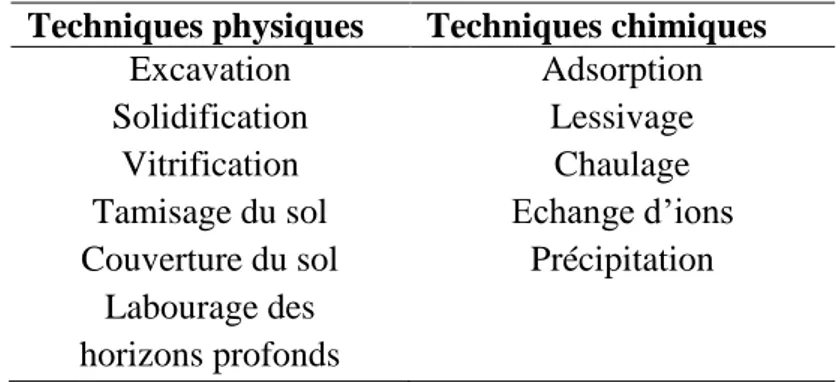
1. Les techniques physico-chimiques
Les techniques les plus utilisées pour décontaminer les sols pollués par les métaux lourds sont les techniques physico-chimiques (Tableau 2). Elles se font in situ ou ex situ. Parmi ces techniques, l’excavation consiste à enlever la couche de sol contaminé et à la déplacer (Mench et al., 2005). Cette technique pose problème car le sol contaminé n’est pas éliminé et il faut aussi disposer de sol non contaminé pour remplacer le sol enlevé (Maenpaa et al., 2002). La solidification est l’ajout au sol d’un élément de cimentation pour former un matériel non poreux, non lessivable et solide (Pierzynski, 1997). Le même résultat peut être obtenu en chauffant le sol pour le transformer en verre (vitrification) (Maenpaa et al., 2002). L’encapsulation ou la couverture du sol (capping) consiste à recouvrir le sol contaminé avec une couche imperméable (Pierzynski, 1997). Dans certains cas, le labourage des horizons profonds est effectué car cela permet de mélanger les couches superficielles contaminées avec les couches profondes non contaminées mais cela ne résout pas le problème du lessivage (Mench et al., 2005). Il existe plusieurs types d’extraction des métaux dont l’extraction électrique pour laquelle des électrodes poreuses sont implantées dans le sol et le courant généré fait migrer les ions métalliques vers la charge opposée à la leur, les polluants sont ensuite récupérés par pompage. L’extraction utilisant une solution acide, afin de solubiliser les éléments métalliques, est aussi possible. Si les sols ne peuvent pas être traités sur le site ils sont excavés et sont par la suite soit stockés dans des décharges soit traités, par exemple par un lavage sous haute pression d’eau ou par lavage avec de l’eau additionnée de tensioactifs (Paff et Bosilovich, 1995 ; Chandra Sekhar et al., 2005).
Les méthodes physico-chimiques de dépollution de sites in situ et ex situ présentent l’inconvénient d’être coûteuses et lourdes à mettre en œuvre. De plus, elles perturbent
12
fortement l’activité biologique des sols et altèrent leur structure physique. Le besoin de nouvelles techniques économiquement compétitives et pouvant préserver les caractéristiques du sol s’est fait sentir et l’utilisation des biotechnologies s’est avérée être une alternative intéressante. C’est pourquoi actuellement de nombreuses recherches sont axées sur de nouvelles méthodes plus écologiques comme la bioremédiation.
Tableau 2 : Les techniques utilisées pour la remédiation d’une contamination métallique du sol
Techniques physiques Techniques chimiques
Excavation Adsorption
Solidification Lessivage
Vitrification Chaulage
Tamisage du sol Echange d’ions Couverture du sol Précipitation
Labourage des horizons profonds
2. La bioremédiation
La bioremédiation fait appel soit à des microorganismes et aux produits de leur métabolisme, soit à des végétaux supérieurs et aux bactéries de leur rhizosphère pour aboutir à la phytorestauration du milieu pollué (Pilon-Smits, 2005).
2.1. La bioremédiation par les microorganismes
Dans les nombreuses formes de bioremédiation, les microorganismes sont utilisés et gérés par le contrôle des facteurs environnementaux pour réduire la pollution de l'environnement. Les processus de bioremédiation utilisent des communautés microbiennes indigènes, y compris les PGPR (Plant Growth Promoting Rhizobacteria), les champignons ou les actinomycètes afin de réduire, éliminer, contenir ou transformer les métaux en produits inoffensifs. Alcanivorax
borkumensis a été utilisé pour éliminer les nappes de pétrole (Gertler et al., 2009). Des
bactéries telluriques du genre Clostridium ont également été utilisées pour la bio-immobilisation du sélénium par réduction indirecte (Le Cloirec and Andrès, 2005).
2.2. La bioremédiation par les plantes « Phytoremédiation »
La phytoremédiation est un cas particulier de la « bioremédiation » qui désigne l’ensemble de techniques qui vise à exploiter la capacité de certaines plantes pour dépolluer et stabiliser des
13
sites (sols, eaux) (Pilon-Smits, 2005) contaminés par divers polluants (organiques et inorganiques) (Chaney et al., 1997).
Chez les plantes, il existe une variété de réponses à un environnement contaminé. Ces réponses correspondent à une tolérance ou non et sont liées ou non à une capacité d’accumulation du métal (Clemens, 2006). Différentes technologies de décontamination basées chacune sur une réponse particulière ont pu être développées. Elles sont classées en cinq catégories (figure 2) : la phytodégradation, la phytoextraction, la phytostabilisation, la phytovolatilisation et la rhizofiltration. Toutes ces technologies sont non exclusives donc complémentaires (Yang et al., 2005).
Figure 2 : Les principales de techniques de phytoremédiation (Moussavou, 2010).
Les ETMs représentés en rouge peuvent s’accumuler dans la plante en conservant la même forme ou être transformés (symboles roses) de différentes manières : dégradation dans le sol ou dans la plante (symboles en croissant), volatilisation dans l’atmosphère (ronds ailés) ou stabilisation dans le sol (carrés).
Dans le cas de la pollution par le Pb, les techniques envisageables pour les sols sont la phytoextraction et la phytostabilisation et pour les eaux, la rhizofiltration.
2.2.1. La phytostabilisation
C’est une technique qui permet de réduire la fraction disponible des polluants dans le sol rhizosphérique, de manière chimique ou biologique, en les stockant dans le système racinaire ou en favorisant son insolubilisation dans la rhizosphère. L'implantation d'un couvert végétal sur un sol pollué limite les risques d'érosion et de lessivage du sol et le transfert des polluants
14
vers d'autres milieux (Kumpiene et al., 2009). La phytostabilisation nécessite des végétaux qui réunissent un certain nombre de propriétés notamment une croissance rapide avec un système racinaire important afin d’augmenter les surfaces de contact sol-plante, une capacité élevée de colonisation du site, être tolérantes aux contaminants, une faible translocation du métal vers les parties aériennes, un cycle de vie long, un fort taux de reproduction et des avantages commerciaux/économiques pour rentabiliser le site contaminé (Mench et al., 2005).
Cette technique peut être améliorée par un apport d'amendements aux sols dans le but de réduire la mobilité et la disponibilité des ETMs. L'ajout des amendements va ainsi augmenter l’adsorption des contaminants sur les phases porteuses solides du sol et favoriser des réactions physico-chimiques (augmentation du pH) ou biologiques qui font diminuer le pool labile en contaminants notamment dans la solution du sol (Gupta and Sinha, 2006). L’ajout de phosphate par exemple dans le sol entraine la formation de précipités stables avec le Pb qui se traduit par une diminution de la biodisponibilté de cet ETM.
2.2.2. La rhizofiltration
La rhizofiltration est une technique qui met en jeu les systèmes racinaires de certaines espèces terrestres ou aquatiques qui absorbent, concentrent et stockent les contaminants des eaux polluées (Ghosh and Spingh, 2005 ; Pilon-Smits, 2005). La rhizofiltration est utilisée pour le traitement des rejets industriels et des effluents agricoles. Elle est particulièrement efficace pour l’élimination du Pb, Cd, Cu, Ni, Zn et Cr, éléments retenus dans les racines (Chaudhry et
al., 1998). Cette méthode présente l’avantage de pouvoir être appliquée in-situ ou ex-situ avec
des espèces végétales pas nécessairement hyperaccumulatrices. Les plantes terrestres sont les plus appropriées pour la rhizofiltration parce qu'elles produisent des racines plus longues et plus importantes et souvent fibreuses avec de grandes surfaces d’absorption de métaux (tableau 3).
On peut citer Helianthus annuus qui est utilisée depuis de nombreuses années pour son potentiel à assainir les eaux polluées par l’uranium et le Pb (Dushenkov et al., 1995 ; Dushenkov et al., 1997) et la fougère Pteris vittata, qui est capable, en culture hydroponique, d’accumuler de grandes quantités d’arsenic (As) (Wang et al., 2002). Le jonc, le roseau et l’iris sont couramment utilisés dans les bassins filtrants. D’autres espèces sont citées dans le tableau 3.
15
Tableau 3 : Plantes proposées pour la rhizofiltration du Pb (Fahr et al., 2013)
Espèce Aire d’application Références
Carex pendula Système aquatique (Yadav et al., 2011)
Pistia stratiotes Eau contaminé (Baharudin, 2008 ; Vesely et al., 2012) Eichhornia crassipes Système aquatique (Tiwari et al., 2007)
Scripus americanus Phaseolus vulgaris
Système aquatique Eau contaminé
(Santos-Díaz and Barrón-Cruz, 2011) (Piechalak et al., 2002)
2.2.3. La phytoextraction
La phytoextraction est une des stratégies de la phytoremédiation basée sur l’utilisation de végétaux (hyper)accumulateurs qui vont absorber les métaux ou métalloïdes du sol et les accumuler dans les organes faciles à récolter (Remon, 2006). C’est la voie dans laquelle les chercheurs se sont le plus investis depuis le début de cette décennie. C’est l’approche qui parait la meilleure pour isoler et éliminer les contaminants du sol sans détruire sa structure et sa fertilité (Ghosh and Spingh, 2005 ; Pilon-Smits, 2005). Les espèces végétales concernées absorbent, concentrent et stockent les métaux toxiques du sol contaminé dans leurs organes aériens (tiges, feuilles, etc.) (Henry, 2000 ; Gisbert et al., 2003 ; Eapen and D'Souza, 2005). Les plantes le plus souvent utilisées pour la phytoextraction des métaux appartiennent aux familles des Brassicaceae et des Fabaceae (Pilon-Smits, 2005). On trouve d’une part, la phytoextraction assistée par des chélateurs de métaux qui est désignée également par la phytoextraction induite et d’autre part la phytoextraction continue.
Une décontamination par la phytoextraction nécessite une répétition de cycles de culture avec récolte de la biomasse à la fin de chaque cycle. Ces cycles se succèdent jusqu'à ce que la concentration de métal dans le sol atteigne une valeur acceptable, ce qui conduit à l’accumulation de biomasse végétale potentiellement dangereuse. Celle-ci doit être stockée de manière à ne présenter aucun risque de contamination pour l’environnement (Blaylock and Huang, 2000). La biomasse peut être traitée par compostage, par compaction, par combustion, par gazéification ou par pyrolyse (Blaylock and Huang, 2000 ; Garbisu and Alkorta, 2001).
La phytoremédiation est une « technologie verte » qui est aussi écologique qu’esthétique, lorsqu’elle est convenablement implantée. Son avantage majeur reste son faible coût par