Les neuropilines
Texte intégral
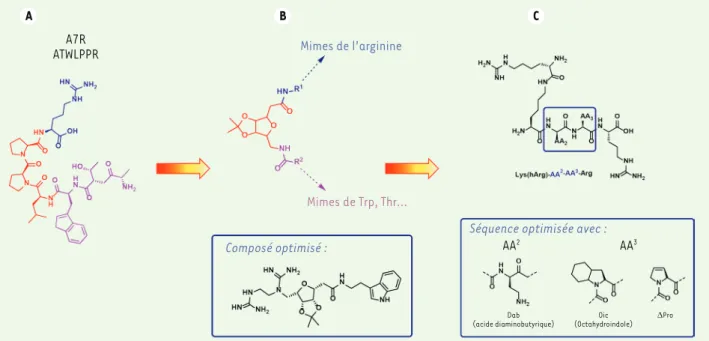
Figure


Documents relatifs
Copyright and moral rights for the publications made accessible in the public portal are retained by the authors and/or other copyright owners and it is a condition of
In the ideal MHD ap- proximation, those are the incompressible Alfv´ en modes (thereafter indexed with A) as well as the fast and slow magnetosonic modes (indexed by F and S), while
Dans les Gaules, les exemples les plus répandus ont été relevés à l'intérieur des habitats, voire à l'extérieur, contre les murs, comme dans la villa de Langeais (Indre-
[r]
We complement our analysis of local ancestry in hybrids by examining fine-scale patterns of genomic differentiation between the parental species, which may capture possible older
Notre recherche se basera donc sur ces deux projets, menés individuellement dans nos classes, et visera à analyser et comparer chez nos élèves l’influence de
Le dessin d’enfant cadre l’exercice, mais sa réinterprétation nécessite une certaine autonomie, une aisance dans le dessin libre que tout le monde n’a pas. En outre, cet
Naturellement, tous les instruments qui permettent à toute personne de transférer des fonds et ce, quelque soit le support ou le procédé technique utilisé. B/- La
