L'évolution de la vie sociale chez les insectes
298
0
0
Texte intégral
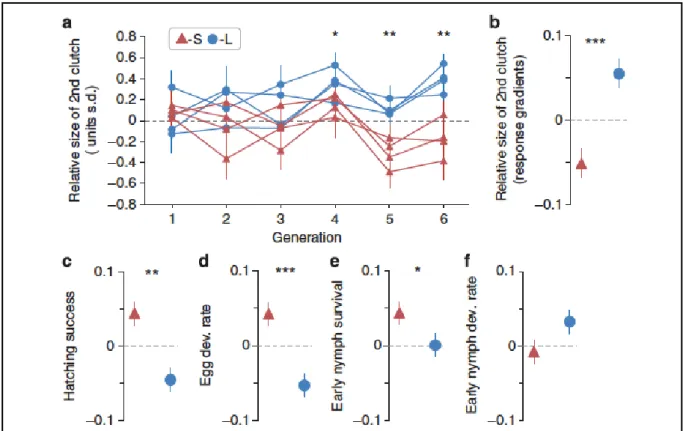
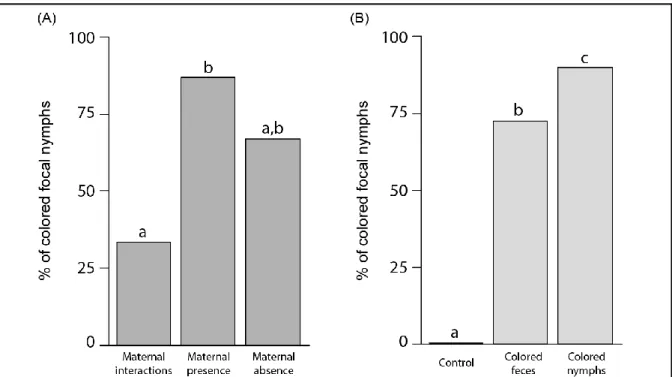
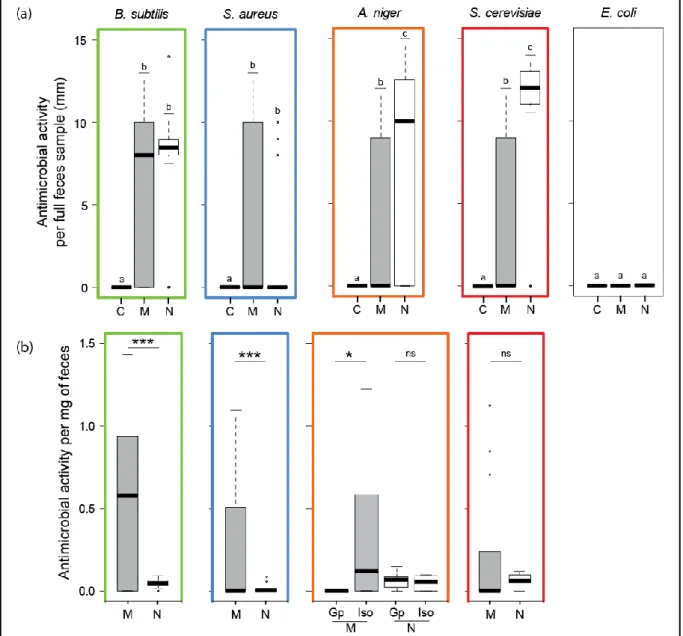
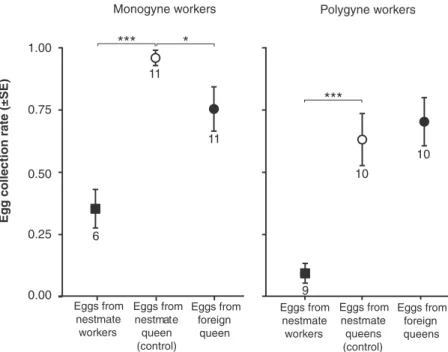
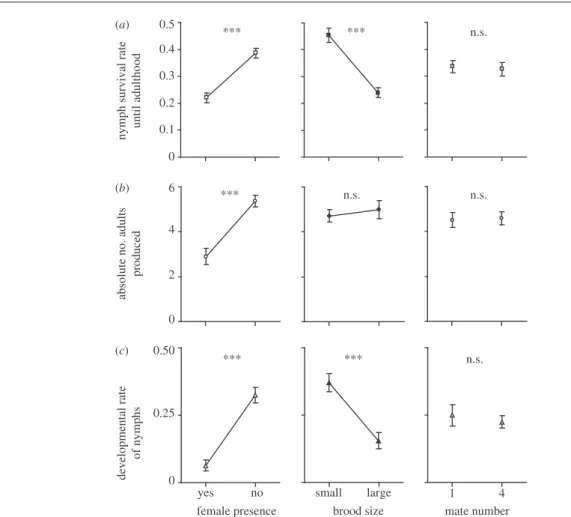
Figure


+7



Documents relatifs