HAL Id: tel-03123055
https://hal.archives-ouvertes.fr/tel-03123055
Submitted on 27 Jan 2021HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Distributed under a Creative Commons Attribution - NonCommercial - NoDerivatives| 4.0
météorologiques : biais et surestimations des estimations
de terrain
Olivier Bouriaud
To cite this version:
Olivier Bouriaud. Réponses de la productivité des forêts aux fluctuations météorologiques : biais et surestimations des estimations de terrain. Biodiversité et Ecologie. Université Paris Sud, 2020. �tel-03123055�
Réponses de la productivité des forêts aux fluctuations
météorologiques : biais et surestimations des estimations de terrain
Olivier Bouriaud
ORCID : http://orcid.org/0000-0002-8046-466X
Google scholar :
https://scholar.google.ro/citations?user=MtTiakoAAAAJ&hl=enResearcherID :
http://www.researcherid.com/rid/C-4700-2011Dossier présenté pour l’obtention de l’Habilitation à Diriger des Recherches
Université Paris-Sud
École doctorale Sciences du Végétal
http://www.ed-sciences-du-vegetal.u-psud.fr
Soutenue le
5 Novembre 2020 devant le jury composé de :
Pr Christopher CARCAILLET, Pr, Directeur des études EPHE, LEHNA, Lyon, rapporteur
Dr Jérôme CHAVE, DR CNRS, UMR 5174 CNRS – Université Paul Sabatier, Toulouse, rapporteur Pr Claire DAMESIN, Pr Univ. Paris-Sud, Laboratoire ESE (UMR 8079), Orsay, rapporteure,
Dr Eric DUFRÊNE, DR CNRS, Laboratoire ESE (UMR 8079), Orsay, examinateur Dr Annabel PORTÉ, DR INRAE, Bordeaux, examinatrice
Mémoire pour obtenir
l’Habilitation à Diriger des Recherches
Contribution à l’étude de la saisonnalité
de l’acquisition et de l’utilisation des ressources
par les arbres forestiers en zone tempérée
par Nicolas Delpierre
Présenté le 03 février 2020
Devant le jury composé de :
— Dr Nathalie Bréda (DR INRA, Silva, Nancy), rapporteure,
— Dr Isabelle Chuine (DR CNRS, CEFE, Montpellier), examinatrice,
— Pr Claire Damesin (Pr Univ. Paris-Sud, ESE, Orsay), examinatrice,
— Dr Hendrik Davi (DR INRA, URFM, Avignon), examinateur,
— Pr Nathalie Frascaria-Lacoste (Pr AgroParisTech, ESE, Orsay), rapporteure,
— Pr Mathieu Jonard (Pr Univ. Louvain, ELI, Louvain), rapporteur.
Sommaire
SOMMAIRE ... 2
1 PRÉAMBULE : OBJETS ET OBJECTIFS DES TRAVAUX DE RECHERCHE ... 3
2 AXE 1. ANALYSE TEMPORELLE DE LA PRODUCTIVITÉ : INTÉGRATION DES DIFFÉRENTS NIVEAUX DE VARIATION PARFOIS IMPLICITES ... 4
2.1 CHOIX DES SITES D’ÉCHANTILLONNAGE : VARIABILITÉ SPATIALE DES CONDITIONS DE MILIEU ... 4
2.2 ECHANTILLONNAGE DES ARBRES : EFFETS CACHÉS DU STATUT SOCIAL ET DE LA TAILLE DES ARBRES ... 5
2.3 DE L’ACCROISSEMENT RADIAL À L’ACCROISSEMENT VOLUMIQUE : VARIABILITÉ AU SEIN DE L’ARBRE ... 7
2.4 EFFETS DE LA DENSITÉ DU BOIS : DIFFÉRENCE ENTRE ACCROISSEMENT VOLUMIQUE ET BIOMASSE ... 7
3 AXE 2. VARIATIONS SPATIALES ET TEMPORELLES DE LA PRODUCTIVITÉ : EFFETS DES MÉLANGES D’ESPÈCES ET SENSIBILITÉ AUX FLUCTUATIONS MÉTÉOROLOGIQUES... 8
4 CONCLUSIONS... 10
1 AMÉLIORATION DE L’ESTIMATION DE LA PRODUCTIVITÉ ... 11
1.1 VARIABILITÉ DE L’ALLOCATION AÉRIENNE DE LA CROISSANCE ... 12
1.1.1 Continuer l’effort de développement des modèles biométriques... 12
1.1.2 Tester des méthodes de calibration locale... 13
1.1.3 Renforcer les aptitudes dynamiques des prédictions de biomasse aérienne : allocation et allométrie .... 15
1.2 APPLICATION DANS LES ESTIMATIONS DE L’INVENTAIRE FORESTIER NATIONAL ... 19
1.2.1 Améliorer les estimations de productivité dans l’inventaire ... 19
1.2.2 Intégrer les erreurs d’estimation dans les estimations sur petits domaines ... 21
2 ANALYSE À TRÈS GRANDE ÉCHELLE DE LA PRODUCTIVITÉ ... 23
2.1 QUANTIFIER LA RÉPONSE DE LA PRODUCTIVITÉ DES FORÊTS AUX ÉVÈNEMENTS MÉTÉOROLOGIQUES À L’ÉCHELLE DE LA RESSOURCE FORESTIÈRE ... 23
2.1.1 Quantifier et incorporer l’autocorrélation temporelle dans les estimations de productivité ... 26
2.1.2 Quantifier la résistance de la productivité à des évènements météo extrêmes ... 28
2.2 INTERACTIONS AVEC LA GESTION ... 32
2.2.1 Relations avec la diversité spécifique et fonctionnelle ... 34
Résumé du parcours de recherche
1 Préambule : objets et objectifs des travaux de recherche
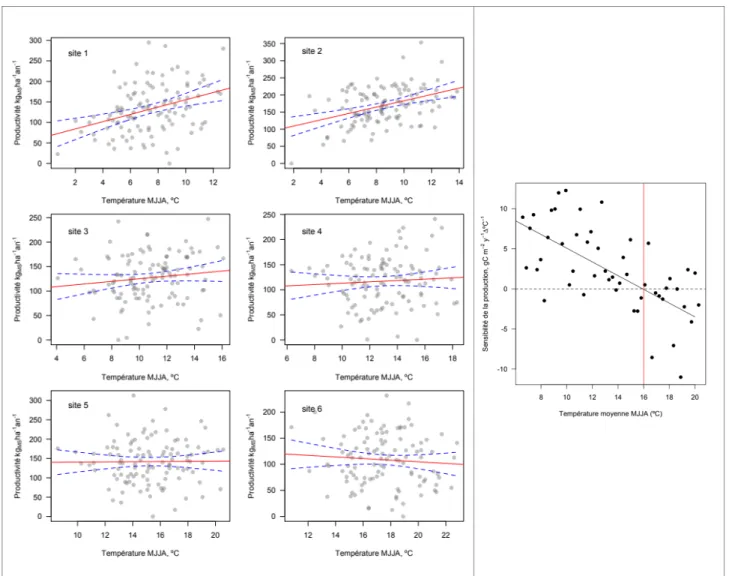
La productivité des forêts est le meilleur indicateur de leur vitalité, et des biens et services en dépendent directement. La productivité est influencée par une grande diversité de facteurs de milieu et anthropiques, à toutes les échelles temporelles. La plupart des études récentes montrent une augmentation globale de la productivité des forêts tempérées et boréales (Hembert et al 2017), mais l’augmentation des évènements météorologiques extrêmes et les dérives climatiques vers des conditions de croissance plus difficiles tracent un avenir beaucoup plus sombre au-delà des prochaines 50 années (Reyer et al. 2017).
Par nature, la productivité est une propriété ou un trait défini à l’échelle d’un peuplement. On peut constater une certaine variabilité dans sa définition. La productivité est la quantité totale de matière végétale produite par une forêt par unité d’espace et de temps (définition déjà présente chez Assman 1970). Cette définition générique convient particulièrement aux études basées sur la mesure ou la simulation des flux de carbone car elle se décline alors en deux termes emboîtés : productivité brute (Gross Primary Productivity) et productivité nette (Net Primary Productivity) telles que NPP = GPP – Ra où Ra est la respiration autotrophe. La productivité nette représente l’entrée autotrophe de matière dans les forêts, qu’elle soit stockée sous forme de bois ou non. Cette définition a donc un caractère très général et proche des processus physiologiques, mais elle est très distante des mesures utiles à la gestion. Il a été acté que l’on ne peut pas estimer la NPP comme la différence entre la GPP et la Ra directement (Waring et Schlesinger 1985). Sur le terrain, on peut l’approcher en estimant la variation du stock de matière organique contenu (aérien et souterrain) dans les arbres et les plantes, moins la quantité perdue : litière, mortalité des racines, exportations et consommations (Clark et al. 2001), pour une surface et une durée données.
Les variations autour des éléments pris en comptes, par exemple aérien seulement, ont conduit à la grande disparité des définitions de la productivité, qu’elles soient explicites ou non d’ailleurs (et trop souvent implicites dans les publications) peut être à cause de la différence du turn-over des compartiments. Ces variations impliquent aussi que les unités soient elles-mêmes très variables -sauf l’unité de temps de référence qui tend à toujours être l’année lorsque l’on ne prend en compte des compartiments à turn-over lent comme c’est le cas dans ce manuscrit, donc excluant les feuilles, fruits et racines.
Dans son usage forestier, la productivité était surtout exprimée en accroissement volumique aérien, du tronc seulement ou de la partie commerciale (en France, tout élément aérien ayant un diamètre supérieur à 7 cm) et a mué en accroissement en biomasse, que l’on convertit au besoin en quantités de carbone. Les estimations basées sur les mesures de terrain se concentrent parfois seulement sur une fraction du compartiment aérien, par exemple le bois fort ou même le bois fort tige, de fait correspondant à ce que l’on peut désigner comme aérien boisé (woody aerial NPP). Les pertes sont souvent négligées, ne subsistant que la partie d’accroissement : litière, exports et consommations. Elles sont particulièrement difficiles à mesurer et quantifier en pratique.
La plupart des connaissances acquises sur la productivité des forêts repose sur des sites expérimentaux bien équipés offrant des connaissances précises dans le domaine de la physiologie tandis que les études à grande échelle sont plus orientées vers la détection de patrons et l’assemblage de lois de comportement phénoménologiques (Bertalanffy 1927). On peut facilement opposer l’approche biométrique à une approche basée sur la télédétection ou l’écophysiologie, même si toutes convergent et que des efforts sont effectués pour inclure des modèles biométriques dans les modèles écophysiologiques (cf Castanea, Orchidee, Sainte-Marie et al. 2014). Toutefois, comme cela sera détaillé plus loin, les analyses biométriques peuvent aider à quantifier les mécanismes à l’œuvre sous-tendus par la productivité.
L’objectif de mes travaux de recherche est d’améliorer les estimations de productivité à différentes échelles spatiales et temporelles, surtout concentrée sur la partie boisée, et approfondir la compréhension des effets de la gestion sur la productivité des forêts. En particulier, déterminer la part du forçage climatique et des effets directs de la gestion sont des objectifs déterminants compte tenu des défis futurs (Reyer et al. 2017, Schulze et al. 2020) : pour nommer les principaux, au plan climatique, la transition climatique, et au plan de la gestion, l’antagonisme entre écologie politique, conservation de la nature et bioéconomie.
2 Axe 1. Analyse temporelle de la productivité : intégration des différents niveaux
de variation parfois implicites
La productivité est définie comme l’accroissement annuel en volume ou en biomasse d’un peuplement donné. L’analyse de la productivité passe par son estimation, ce qui pose des questions méthodologiques importantes malgré les apparences simples du problème. La croissance radiale des arbres est un des éléments les mieux décrits dans la littérature et étudié depuis de très nombreuses années (domaines de la dendroclimatologie et de la dendroécologie notamment), mais qui n’est qu’assez indirectement lié à la productivité lorsque celle-ci est exprimée en termes de biomasse ou de quantités de carbone fixés par unités de temps et de surface. La raison de la perte de proportionnalité directe entre croissance radiale et productivité se structure en plusieurs termes :
• le manque de proportionnalité entre la croissance secondaire et la croissance primaire (celle-ci ne contribuant cependant que très peu à la productivité à l’instant t), et la variabilité de la croissance secondaire au long du tronc puis dans la couronne
• Le découplage existant entre croissance individuelle et production totale dans des communautés végétales fermées (loi de production des constantes, ou density-biomass relationships, Enquist 2002)
• le découplage entre la croissance radiale et la variation de la densité du bois, pas uniforme selon les classes botaniques d’espèces d’arbre.
• l’échantillonnage, qui renvoie directement aux questions typiques des programmes d’inventaire forestier nationaux et qui tient donc à un axe de recherche spécifique, et qui tient à la diversité entre arbres si l’on raisonne à l’échelle d’une placette, à la diversité entre placettes à plus grande échelle.
Chacun de ces termes contribue à introduire des incertitudes et possiblement des biais d’estimation, lesquels peuvent inverser une tendance ou masquer un des effets à analyser. Leur analyse a commencé lors de mes travaux de thèse, et ont continué sur cet axe directeur, et sont détaillés ici.
2.1 Choix des sites d’échantillonnage : variabilité spatiale des conditions de milieu
La dendrochronologie reste influencée dans ses méthodes par l’idée d’un déterminisme climatique prédominant, du moins le recherche-t-elle de façon privilégiée. Rapidement les séries dendrochronologiques seront utilisées pour réaliser des reconstructions des températures ou des précipitations (Briffa et al. 1992, 2004) voire même de la sécheresse (Stahle et al. 1985). Les échantillonnages ont été très influencés par ces objectifs, qui tendent à maximiser la corrélation entre croissance et climat. L’idée sous-jacente est celle d’un facteur climatique très dominant, suivant la loi de Liebig.
Le choix des sites d’échantillonnage était traditionnellement orienté vers la maximisation du signal climatique et a conduit à choisir des placettes peu représentatives de la ressource forestière (Babst et al. 2018, 2019, Zhao et al. 2019). Tant que le sujet principal de l’étude était le climat, dont on voulait une reconstruction, cette stratégie était acceptable. La divergence de la sensibilité de la croissance aux températures a fait apparaitre des doutes sur la stabilité des relations croissance-climat (D’Arrigo et al. 2008). En abordant la représentativité des sites d’étude ou à la population d’arbres échantillonnés, c’est aussi
la reconnaissance d’un déterminisme complexe et multi-factoriel de la productivité qui a été reconnue (hypothèse de limitation multiple, par opposition à l’hypothèse de Liebig. Cette idée m’est apparue en partant des analyses dendrochronologiques réalisées dans un site atelier donné, dont la représentativité géographique est très difficile à quantifier (par exemple, partant du site de Hesse, support de ma thèse, et généralisant aux hêtraies du Nord Est). Par opposition, les inventaires forestiers obligent à installer des placettes d’une manière représentative issue de l’échantillonnage lui-même.
Les placettes constituées pour les mesures de croissance rétrospective ont longtemps favorisé des conditions de croissance extrêmes, soit très froides, soit très sèches (Charney et al. 2016). Dans tous les cas, la sensibilité à un facteur climatique donné était maximisée. Ce fait a été récemment démontré par une comparaison avec les analyses climatiques issues de dispositifs d’inventaire forestier (Klesse et al. 2018). Dans le cas des placettes permanentes au contraire, ce sont les peuplements productifs qui ont eu tendance à être échantillonnés (Gillis et al. 2005). Elles ne sont donc pas utilisables pour une estimation à grande échelle de la productivité. Les estimations dérivées de télédétection suggèrent des variations régionales marquées, mais dont les limites sont floues et fluctuantes (Guay et al. 2014). La variabilité spatiale est très claire, et développer des prédictions régionales fiables nécessite ainsi une réplication suffisante.
Continuité de mon travail au Centre Canadien des Forêts, Girardin et al. 2016 montre l’intérêt d’avoir une base empirique très large et aléatoire dans laquelle le positionnement des placettes n’est pas lié à un critère donné. Le réseau de placettes, qui est celui de l’inventaire forestier national du Canada, permet d’obtenir des estimations de tendances et de niveaux de croissance régionaux précis. Des tests de ré-échantillonnage ont permis de démontrer qu’il n’y avait pas de biais apparent dans les estimations, même pour les espèces peu échantillonnées. La puissance de ce dispositif a par ailleurs été démontrée par son aptitude à saisir l’influence de l’enrichissement de l’air en CO2 sur la croissance. Les résultats montrent en effet que l’assemblage de
séries individuelles de croissance au niveau régional ne permet pas de passer le seuil de détection de signaux communs très faibles comme celui de la variation de concentration en CO2 de l’air, lequel ne devient
significatif que lorsque l’ensemble des placettes sont incluses, indiquant que l’effectif à avoir pour détecter un signal aussi faible est très important.
L’utilisation des séries dendrochronologiques comme source d’estimation à grande échelle de la productivité, par exemple pour fournir des références aux estimations issues de télédétection ou modélisation, demande des efforts d’intégration. Marchand et al. 2018, continuant la réflexion de Girardin et al. 2016, insiste sur la nécessité d’intégration des mesures dendrochronologiques et d’inventaire, et souligne la possibilité de biais dans l’échantillonnage des placettes lorsque sont exclus les peuplements ayant fait l’objet de perturbations, naturelles ou anthropiques.
2.2 Echantillonnage des arbres : effets cachés du statut social et de la taille des arbres
Les arbres échantillonnés (y compris souvent dans mes recherches) ont très globalement toujours été des arbres dominants. Ils ont en effet deux avantages principaux : i) ils sont gros et contribuent avec beaucoup de cernes, bien lisibles ; ii) la variation de la croissance radiale est supposée moins influencée par la dynamique du peuplement que pour les autres arbres (Fritts 1976). C’est cette propriété de relative indifférence des arbres dominants à la compétition qui est à la base de l’indice de fertilité et de la relation entre hauteur dominante et productivité, par exemple telle que résumée dans la loi de Eichhorn élargie (Decourt 1973) mais à laquelle ont été trouvés depuis bon nombre de contre-exemples (Cieszewski et Bella 1993, Pardé et Bouchon 1988, Meredieu et al. 2003).
Choisir des arbres dominants procédait du même effort d’élimination des variations de croissance non liées aux facteurs recherchés, essentiellement climatiques, et servait donc à la facilitation de l’analyse des données. Implicitement il est admis que le statut social des arbres a une influence sur les séries de croissance, mais ici c’est plutôt la variabilité inter-annuelle et la présence de variations non climatiques que l’on
cherchait à éliminer, plutôt que le niveau de croissance. Or, dans l’analyse de la productivité, le niveau absolu de croissance est lui aussi très important.
Les sauts de croissance liés aux éclaircies ou tout changement de statut social créent des ruptures problématiques pour l’analyse des données, car les méthodes statistiques utilisées pour estimer la relation entre la croissance radiale et un facteur donné se basent sur des séries temporelles normalisées (supposées) stables (Wilmking et al. 2020). En toute rigueur il faut non seulement une moyenne constante mais aussi une variance constante sur la période d’analyse. Or des fluctuations soudaines de croissance compromettent l’un et l’autre, et corriger une série présentant un saut -en fait même détecter un saut ou toute anomalie dans la série temporelle- pose son lot de difficultés méthodologiques.
L’importance du choix des arbres à échantillonner a surgi dans le contexte des études des tendances à long terme (Brienen et al. 2012, Nehrbass-Ahles et al. 2014, Marchand et al. 2018). Deux sources de biais tenant à l’échantillonnage sont évoquées : la première tient au fait que les arbres maintenant dominants ont pu être dominés autrefois et leur changement de niveau de croissance tient, entre autres, à l’amélioration de leur position sociale (Cherubini et al. 1998). La deuxième source tient au fait que les arbres les plus vieux sont globalement ceux qui croissent le plus lentement (Brienen et al. 2012, Melvin 2004), par exemple parce qu’ils seraient sur un site de faible fertilité. Dans les deux cas les changements de croissance, positifs, ne sont pas liés à des tendances climatiques ou de fertilité et constituent ainsi des artéfacts ou sources de biais dans les analyses (voir par exemple : Bontemps et Esper 2011). On observe que c’est ici le niveau de croissance, et non la variabilité inter-annuelle, qui est au cœur du problème, nous recentrant sur la problématique de productivité. La question de la représentativité de l’échantillon utilisé pour détecter des tendances de croissance s’est posée aussi grâce aux réflexions méthodologiques autour de la méthode de standardisation et d’extraction des tendances, méthode dite de standardisation des effets ontogéniques par une courbe de référence régionale (Regional Curve standardisation, ou RCS). Ces réflexions ont en effet rapidement pointé vers le choix des arbres et l’étendue géographique associée (Esper et al. 2012).
Lors de l’analyse des données dendrochronologiques prélevées dans le cadre de l’inventaire forestier du Canada, la question de l’échantillonnage des arbres a émergé fortement, les arbres carottés ayant un diamètre supérieur à la moyenne de la placette, sans élément expérimental pour y répondre. Les discussions que j’ai conduites avec les collègues de WSL spécialistes des tendances à long terme (Jan Esper et David Frank) ont ainsi fait émerger un thème de recherches dédié aux effets de l’échantillonnage, avec deux résultats principaux. Le premier, obtenu par la thèse de Flurin Babst que j’ai codirigée, a abouti à définir un
protocole d’échantillonnage dédié à la reconstruction de la productivité combinant des mesures dendrométriques et dendrochronologiques (Babst et al. 2013, 2014a,b). Ces publications se basent sur un
jeu d’inventaires et de mesures dendrochronologiques constitué dans le cadre du projet CarboExtreme (FP7). Petit clin d’œil à l’inventaire national roumain dont j’étais déjà directeur technique, le seuil de précomptage dans ces publications est de 5,6 cm, défini comme la base de la classe de diamètre allant de 6 à 7 cm. Le deuxième est un test de sensibilité destiné à quantifier l’importance de l’échantillonnage des arbres sur l’estimation des tendances à long terme de la productivité et de sa sensibilité au climat (Nehrbass-Ahles et al. 2014). Il s’agit du premier article méthodologique réellement axé sur les biais que peuvent provoquer un échantillonnage. L’analyse fut réalisée en simulant les diverses procédures d’échantillonnage dendrochronologiques classiques d’une placette de 0,5 ha complètement inventoriée. Cette étude a mis en évidence le fait que la méthode la plus couramment utilisée jusqu’alors, qui se concentre sur les arbres
dominants, génère des biais d’estimation de la croissance allant jusqu’à 459% et des biais d’estimation des tendances dépassant les 200%. Les analyses à courte fréquence de la réponse de la croissance au climat
restent par contre très peu affectées par les méthodes d’échantillonnage.
Comme montré par Babst et al. 2014a, l’échantillonnage doit tenir compte de contraintes pour permettre de réaliser le passage (implicite mais fondamental) de l’échelle de l’arbre à celui du peuplement. Les méthodes développées ont depuis été mises en œuvre dans d’autres projets tels que dans FundivEurope (FP7), où les
inventaires de tous les sites d’étude et les mesures dendrochronologiques ont été réalisées par mon équipe à l’Université de Suceava.
2.3 De l’accroissement radial à l’accroissement volumique : variabilité au sein de l’arbre
La loi de Pressler (publiée en 1864) prédit que la surface d’un cerne annuel est constante de la base du tronc jusqu’à la base du houppier, puis décroit linéairement jusqu’au sommet de l’arbre. Cette loi a un fondement physiologique basé sur le transport de l’eau, si l’on prend comme fonction principale du cerne le transport de la sève : la surface conductrice à une hauteur donnée est contrainte par la quantité de feuilles positionnées au-dessus de ce point. C’est en somme ce que soutient le modèle de conduction de l’eau dit pipe-model (Shinozaki et al. 1964) qui a eu le succès qu’on lui connait, même s’il a connu des révisions substantielles (Enquist 2002).
La proportionnalité entre la croissance radiale et l’accroissement volumique du tronc ou de l’arbre entier est une hypothèse implicite dans beaucoup d’études, surtout dendrochronologiques (ex : Bergès et al. 2002), mais n’a reçu que peu d’attention. Une analyse de tige réalisée sur 30 hêtres de grande taille pendant ma thèse a montré que la croissance radiale à 1,3 m n’est pas strictement proportionnelle à l’accroissement volumique du tronc (Bouriaud et al. 2005). La corrélation entre la croissance observée à 1,3 m et celle plus en haut dans le tronc baisse continument le long du tronc. Cette analyse a montré davantage encore que la
réaction à la sécheresse était beaucoup plus marquée à 1,3 que plus en haut dans le tronc -et donc que l’accroissement volumique : un début de croissance plus tardif vers la base du tronc, et une distance plus
grande aux feuilles résulte dans une moindre croissance lors d’une sécheresse. Cette analyse a été répétée sur des épicéas en Allemagne dans la thèse de Marieke van der Maaten-Theunissen à laquelle j’ai été associé, et qui confirme et généralise les résultats obtenus sur le hêtre (van der Maaten-Theunissen et Bouriaud 2012). On peut probablement généraliser ces constats à la discrimination isotopique des éléments chimiques : l’étude récente de Jucker et al. 2017 en effet montre que le signal isotopique δ13C est un très bon indicateur
de sécheresse (définie comme il se doit par rapport à une limite de 40% une réserve d’eau extractible du sol, Granier et al. 1999) mais qui exagère la réponse de l’accroissement en surface terrière aux sécheresses. Il n’y a en effet pas de proportionnalité dans les séries respectives, la discrimination marquant les sécheresses
plus fortement que l’accroissement et témoigne ainsi des capacités des arbres à limiter les effets de la
sécheresse dans une certaine mesure.
2.4 Effets de la densité du bois : différence entre accroissement volumique et biomasse
Entre le volume et la biomasse intervient la densité du bois. Souvent implicitement constante, comme par exemple dans les modèles de biomasse, elle varie elle aussi sensiblement à toutes les échelles : dans le cerne, dans l’arbre, et entre arbres d’un peuplement (Guilley et al. 1999, etc) et peut être corrélée à la largeur de cerne. Généralement la corrélation est négative chez les résineux : à un cerne large correspond une densité annuelle faible, ce qui peut conduire à une compensation, au moins partielle, d’une baisse de croissance. Mais il semble exister des différences entre espèces, pas seulement en amplitude mais aussi en signe, avec une corrélation positive chez les chênes.
Deux situations distinctes peuvent être différenciées : le cas d’une corrélation claire entre largeur de cerne et densité (annuelle), et le cas où il n’y a pas de relation claire. Ce dernier cas est celui du hêtre, où sont observables plutôt des tendances que des relations stables, s’agissant d’un bois à pores diffus. Toutefois, Bouriaud et al. 2004 montrent que la densité du bois est moins variable que la largeur de cerne, bien que corrélée aux fluctuations climatiques sur une fenêtre temporelle plus large. Les facteurs climatiques sont d’ailleurs différents de ceux influant sur la largeur de cerne. Il s’agit donc d’un découplage dans les
fluctuations de ces séries temporelles, qui vont dans le sens d’une plus forte variation de la largeur de cerne.
Dans le cas de l’existence d’une corrélation entre largeur de cerne et densité, comme chez l’épicéa, le découplage est encore plus fort. Comme nous l’avons montré (Bouriaud et al. 2005a), la densité du bois peut être expliquée par un phénomène de « dilution » dans la deuxième moitié de la saison de croissance, où il existe un rapport direct négatif entre la croissance radiale et la densité. Cette dilution se manifeste par le fait que la densité diminue de manière inversement proportionnelle à la vitesse de croissance instantanée, et signifie que la quantité de matière produite reste constante tandis que le volume occupé varie.
Quantifier les erreurs d’estimation de l’accroissement en biomasse générées par la négligence de la corrélation entre largeur de cerne et densité du bois est difficile. A base de mesures et d’analyse de tige, nous avons montré (Bouriaud et al. 2015) que l’effet de compensation (une moindre densité lorsque la largeur de cerne augmente, et inversement) se traduit par une erreur sur l’accroissement en biomasse allant jusqu’à 20%. Dans une étude de plus grande ampleur (5 sites de mesures de flux Eddy-covariance) mais basée sur le même principe et portant sur plusieurs espèces, nous avons montré (Babst et al. 2014a) que tenir compte des variations inter-annuelles de densité du bois, y compris des effets de compensation (densité plus grande lors de croissances faibles) augmente sensiblement la corrélation entre les estimations de productivité nette de l’écosystème (NEP) issues des mesures biométriques et les estimations issues des mesures de flux. Cette
étude souligne donc l’influence significative des compensations dans les estimations de productivité.
Un troisième facteur non négligeable vient de la variation entre arbres. Pour le moment le déterminisme de cet effet individuel reste très largement inconnu et est couramment traité comme un effet aléatoire (donc effectivement imprévisible). Il a d’ailleurs été démontré sur l’inventaire forestier national français que la densité du bois d’un arbre localement proche d’un autre, de même taille et de même espèce n’est pas un meilleur prédicteur de la densité du bois de son voisin que la densité de l’arbre moyen de même espèce dans une région (projet XyloDensMap, Bontemps, comm. pers.) La deuxième partie de ce mémoire aborde ce sujet et des pistes. Les effets aléatoires se manifestent de deux manières : la première en influençant le niveau moyen (la densité moyenne), le deuxième en influençant la relation entre la densité et la largeur de cerne. Les deux effets sont effectivement observés (Bouriaud et al. 2004, 2015).
3 Axe 2. Variations spatiales et temporelles de la productivité : effets des
mélanges d’espèces et sensibilité aux fluctuations météorologiques
Les recherches présentées dans l’axe 1 ont un caractère méthodologique assez marqué, même si elles apportent des éclairages forts sur les mécanismes physiologiques de la croissance (par exemple, la répartition de la croissance, le découplage entre assimilation et croissance), ce que l’on peut voir en examinant le profil des articles qui citent ces travaux. L’axe 2 a une orientation beaucoup plus écologique. Dans cet axe les estimations rétrospectives de la productivité sont analysées en relation avec les facteurs de milieu ou du peuplement tel que la diversité des espèces. Il s’agit d’une évolution assez naturelle du premier axe car basé sur ses acquis méthodologiques, et mis en œuvre essentiellement grâce à la participation à des projets internationaux très actifs, et le suivi de thèses en doctorat conduites en Italie (Université de Florence, Laboratoire de botanique théorique et appliquée). D’un point de vue fonctionnel, cet axe de recherche prolonge la question de la stabilité de la productivité, mais en prenant en compte les différences
inter-spécifiques dans le niveau de productivité et dans leur sensibilité aux fluctuations climatiques. La
comparaison de la sensibilité de la productivité aux fluctuations météorologiques était un objectif de l’étude démarrée en 2007 en Roumanie, qui se basait sur la comparaison de la croissance et de la réponse au climat de trois espèces participant de manière comparable au couvert dominant d’une parcelle donnée, donc croissant toutes dans des conditions identiques. Les résultats encourageants de cette étude, consistant en
une sensibilité très différente des espèces étudiées toutes conditions égales par ailleurs (Bouriaud et Popa 2009), ont également montré l’importance du contexte et soulignent la nécessité d’être opposés à
d’autres espèces et d’autres conditions de milieu, en particulier de sortir du contexte du site d’étude pour s’étendre à un réseau à plus grande échelle.
La diversité (ici le nombre d’espèces d’arbres) a une influence positive sur la stabilité temporelle de la productivité. Ce résultat, connu pour les communautés végétales herbacées, devait être confirmé pour les forêts tempérées. Dans le cadre du projet FP-7 FundivEurope a été mise en place une infrastructure qui consiste en placettes de mesure dont le mélange d’espèces est contrôlé, et installées sur six sites d’étude régionaux allant de l’Espagne, l’Italie à la Finlande (Baeten et al. 2013). Les sites et les réseaux de placettes optimisent le découplage des facteurs d’ordinaire confondus, tels que les facteurs de milieu principaux (type de sol et profondeur, exposition, altitude) tout en offrant un contraste de conditions climatiques très important. Se basant sur ces mesures, nous avons pu calculer (Jucker et al. 2014) la productivité rétrospective (croissance aérienne annuelle en biomasse) en utilisant les méthodes présentées dans Babst et al. 2014b qui associe des mesures dendrométriques et dendrochronologiques. Deux indices ont été utilisés pour analyser la productivité : selon Gross et al. (2014), la productivité d’un peuplement mélangé est comparée à celle de peuplements purs ce qui permet de déceler si la moyenne (la surproduction : overyielding) ou l’écart-type (stabilité) de la productivité est la plus affectée par le mélange. En plus de ces indices, a été calculé un indice de synchronicité selon Loreau et de Mazancourt (2008), afin d’apporter des éléments quantitatifs d’explication des effets de la diversité sur la productivité. Les résultats ont montré une moindre variation
inter-annuelle (donc plus grande stabilité) des peuplements mélangés, et que cette plus grande stabilité pouvait s’expliquer par un découplage (asynchrony) de la réponse des espèces aux fluctuations du climat.
En 2017, dans le cadre du projet SoilForEurope (H2020), mon équipe de l’Université de Suceava a procédé à la remesure de toutes les placettes du réseau du projet FundivEurope. Ces nouveaux inventaires, réalisés par les mêmes personnes et avec les mêmes méthodes, vont fournir des estimations de la productivité à base de comparaison d’inventaire. Ils fourniront enfin les informations démographiques que les protocoles de reconstruction ne peuvent fournir : recrutement, mortalité. Ils offrent ainsi une perspective temporelle longue qui manquait jusqu’ici aux études portant sur la diversité. Les recherches prévues dans ce sens sont développées dans la partie suivante.
La seconde échelle posant problème dans les analyses portant sur les interactions diversité-productivité est l’échelle spatiale. Les relations entre la biodiversité et le fonctionnement des écosystèmes
commencent à être bien appréhendées (exposés en détail dans la partie suivante), mais la plupart des preuves de terrain proviennent d'études à petite échelle. La multiplicité des facteurs qui déterminent le fonctionnement des écosystèmes et les éléments de la biodiversité sont un obstacle qui limite drastiquement l'étendue spatiale des études. En outre, l'extrapolation des structures de la biodiversité-écosystème (B-EF) demeure difficile, en partie parce que l'importance des facteurs environnementaux dans la modélisation des relations B-EF est mal comprise. Par exemple, pour étudier l'ampleur et les moteurs potentiels de la dépendance au contexte des relations B-EF, 26 fonctions écosystémiques ont été mesurées le long du gradient de diversité composite des essences dans six régions d'Europe au sein de la plateforme de recherche FundivEurope. Les analyses effectuées au sein de cette plateforme ont montré que, malgré des variations considérables de la richesse en espèces à travers le continent, il existe une tendance à des relations B-EF
plus fortes dans les régions au climat plus sec et dans les zones avec des saisons de croissance plus longues et des types d'espèces plus diversifiés (van der Plas et al. 2016, Ratcliffe et al. 2017). L'importance
de la disponibilité en eau suggère que lorsque les contraintes hydriques augmentent, par exemple dans le contexte du changement climatique, la biodiversité peut avoir une influence encore plus importante dans le soutien des niveaux élevés de productivité des forêts européennes.
La situation des forêts méditerranéennes, avec leur déterminant climatique dominant est particulièrement intéressante. D’une part parce qu’elle met en jeu des espèces déjà très bien adaptées aux rigueurs climatiques, a priori très résistantes (résistance et résilience sont mieux définis dans la section suivante), d’autre part, hélas, à cause de la dynamique des changements climatiques. La collaboration commencée dans le cadre du projet FundivEurope avec l’Université de Florence s’est continuée par le co-encadrement de la thèse de Giovanni Iacopetti (commencée en fin 2016, directeur Filippo Bussotti et Feredico Selvi) sur le sujet de diversité des arbres et fonctions des forêts, analyse intégrée basée sur les bases de données nationales et
inventaires de terrain. Un des objectifs de cette thèse à laquelle je suis associé est d’examiner les effets de la diversité sur la résistance à la sécheresse dans les forêts thermophiles de Toscane. Il s’agit pour l’essentiel de taillis abandonnés offrant une forme de stabilité temporelle à la fois dans la composition des espèces et une absence de facteurs perturbateurs. Dans le cadre de ces analyses, nous avons utilisé les acquis méthodologiques passés mais aussi utilisé de nouvelles méthodes d’analyse des séries temporelles dédiées aux évènements climatiques extrêmes qui constituent des ruptures dans les séries. Ces nouvelles méthodes sont un point de vue très statistique de la question des anomalies de croissance et s’écartent de la méthode traditionnelle de détection des années caractéristiques, et ont l’avantage double de fournir un test avec une signification et de détecter beaucoup plus que juste une anomalie (détails fournis dans la partie projet) (Druckenbrod 2005). Ces analyses ont très rapidement apporté des résultats nouveaux donnant une autre perspective à des séries temporelles souvent vues comme continues. Afin d’approfondir les analyses portant sur la réaction des espèces face à ces anomalies, et d’aborder leur capacité de résistance, cette thèse va être poursuivie par une seconde, à démarrer au mois d’Août 2020, avec pour titre Impact des changements climatiques sur la résilience de la croissance des peuplements de chêne chevelu (Quercus cerris L.) à base des données à long-terme des réseaux de monitoring, directeur Filippo Bussotti). Cette nouvelle thèse, concentrée sur une espèce importante des zones méditerranéennes, aura une approche beaucoup plus mécaniste et physiologique car elle évaluera la résilience et la vulnérabilité de Q. cerris sur les sites d'étude en combinant l’analyse de l'anatomie du bois, de la réserve de glucides dans la tige et la branche, ainsi que la teneur en amidon dans les rayons du parenchyme et les isotopes du carbone.
4 Conclusions
Les travaux conduits ont permis d’obtenir des résultats sur les deux dimensions de la productivité, le temps et l’espace. Certains sont essentiellement d’ordre méthodologique, s’agissant d’une meilleure quantification de la productivité, mais ont apporté des éléments fondamentaux dans la compréhension de la productivité et de sa stabilité (Tableau 1). D’autres résultats constituent déjà des applications dans le champ de l’écologie et dessinent les directions de recherches futures, décrites et développées dans le projet.
Tableau 1. Tableau récapitulatif et synoptique des résultats portant sur l’analyse de la productivité
Question de recherche Résultat obtenu Publication
Variabilité de la croissance secondaire au long de la tige
Réaction à la sécheresse beaucoup plus marquée à 1,3 m que plus en haut dans le tronc, non proportionnalité croissance radiale/volumique
Bouriaud et al. 2005
van der Maaten-Theunissen et Bouriaud 2012
Découplage entre croissance radiale et densité du bois
Découplage entre la croissance radiale et la variation de la densité du bois, pas
uniforme selon les classes botaniques d’espèces d’arbre Bouriaud et al. 2004 Bouriaud et al. 2005 Babst et al. 2014 Nehrbass-Ahles et al. 2014 Bouriaud et al. 2015 Découplage entre croissance
individuelle et production totale dans des communautés végétales fermées
Nécessité d’un protocole d’échantillonnage dédié à la reconstruction de la productivité, sinon biais importants (20-200%)
Babst et al. 2013 Babst et al. 2014
Nehrbass-Ahles et al. 2014 Bouriaud et al. 2015 Divergence sensibilité spécifiques Différences de sensibilité des espèces
toutes choses égales par ailleurs Bouriaud et Popa 2009 Effet des mélanges sur la stabilité de
la productivité Moindre variation inter-annuelle (donc plus grande stabilité) des peuplements mélangés
Jucker et al. 2014
Iacopetti et al., en évaluation Les nombres entre crochets font référence aux articles de la liste d’articles dans des revues à comité de lecture.
Partie 2. Projet scientifique
1 Amélioration de l’estimation de la productivité
Les efforts d’estimation de la productivité à base de mesures biométriques demeurent très dépendants de l’estimation du stock, car, dans la très grande majorité des situations, la productivité biologique est estimée par différence de stock entre deux observations. L’aspect dynamique paraît plus difficile à prendre en compte que la variabilité spatiale, essentiellement parce que les mesures destructives sont statiques : elles permettent de décrire la distribution, mais pas l’allocation. Les modèles allométriques qui en découlent sont donc statiques aussi. S’ils permettent de modéliser efficacement la variabilité statique telle que représentée dans l’échantillon, leur capacité de prédiction de la dynamique n’est pas garantie. Depuis Prodan (1965) on sait que la relation hauteur-diamètre d’un échantillon n’est pas une bonne représentation de la courbe de croissance que suivra un arbre lors de son développement (ex. figure 1). C’est pourtant bien de cette manière-ci qu’ils sont utilisés lorsque l’on veut estimer l’accroissement en biomasse d’un arbre donné.
Figure 1. Relations hauteur-diamètre pour des épicéas en
fonction de leur âge, d’après Prodan (1965).
Rien n’indique que l’allocation soit une forme différentielle de la distribution de ces valeurs, car différents compartiments de l’arbre peuvent avoir des dynamiques différentes et qui se compensent. Les changements dans l’allométrie avec l’âge sont connus depuis longtemps : Niklas (1995) conclut à un changement de la relation hauteur-diamètre avec le développement. Plus récemment, Genet et al. (2011) démontrent que l’allométrie du tronc et de la couronne varient avec l’âge des arbres. Une des mesures essentielles pour limiter cet effet est d’ajuster la dérivée du modèle statique. Les modèles de volume et de biomasse développés dans le cadre du projet H2020 «DIABOLO» sont précisément basés sur des formes mathématiques dédiées aux accroissements et intègrent les contraintes sur les relations et les valeurs des paramètres liées à la croissance. On peut donc considérer que la partie théorique qui devrait permettre de produire des modèles plus adaptatifs est bien connue, mais qu’elle n’a pas été mise en œuvre. Une nouvelle génération de modèles est donc à concevoir mais la disponibilité des mesures de terrain demeure très limitante.
Obtenir des mesures de la croissance reste ainsi très difficile, particulièrement pour les couronnes. En comparant la distribution pour des années successives, on peut déduire l’allocation (la partie ligneuse de celle-ci). Deux types de mesures complémentaires sont possibles : les mesures micro-dendrométriques, et les télé-mesures. Les micro-dendromètres ont montré leur potentiel pour le suivi de la croissance radiale des organes ligneux. Leur niveau de précision est tel qu’ils permettent même de quantifier les variations intra-annuelles de la croissance en dépit du bruit causé par les variations de diamètre liées au potentiel hydrique du bois et l’humidité de l’écorce. Leur précision diminue sur des organes de petit diamètre, et leur mise en place
nécessite des moyens techniques qui en limitent l’applicabilité. Ils n’en demeurent pas moins une référence, surtout s’agissant de mesurer la croissance primaire des branches principales. Les télémesures englobent deux méthodes distinctes : les mesures optiques et les mesures par scanner laser terrestre. Les mesures optiques (à l’aide d’un théodolite) ont longtemps fait office de référence et leur précision est suffisante pour mesurer l’accroissement en diamètre sur une période de plusieurs années, ou l’accroissement en longueur (croissance primaire).
Les éléments suivants ont pour objet de présenter les recherches visant à apporter un saut quantitatif dans les estimations de la productivité, ayant en tête ces contraintes et se focalisant sur deux aspects complémentaires : d’une part, la modélisation du volume et de la biomasse, aspect spatial de la biométrie des arbres, d’autre part, l’allocation de la croissance, représentant son aspect temporel.
1.1 Variabilité de l’allocation aérienne de la croissance
1.1.1 Continuer l’effort de développement des modèles biométriques
Pratiquement toutes les estimations de volume et de biomasse font appel à des modèles de biomasse ou de volume. Mais les erreurs de prédiction des modèles ont une amplitude représentant environ 10 à 40% de l’estimation elle-même. Toute amélioration des modèles offrirait donc un gain appréciable sur les prédictions. La multiplicité des sources de variation de l’allométrie impose l’utilisation de formes de modèles assez souples pour les absorber. C’est ce qui a été démontré dans le cadre du projet DIABOLO. L’objectif de la tâche T2.1 de ce projet était justement de développer des équations compatibles de volume et de biomasse, en vue d’une application à tous les pays européens pour les principales espèces forestières (Work Package 2 Tâche 2.1, leader Olivier Bouriaud, http://diabolo-project.eu/about/estimations/). Ces modèles sont conçus pour être appliqués aux données collectées dans les inventaires forestiers européens. Ils sont basés sur une nouvelle formule mathématique d'allométrie des arbres qui, en essence, prend en compte le rôle fondamental des processus d’ontogenèse (Gatsuk et al. 1980). Ils tiennent compte explicitement de la possible divergence de la dynamique de la forme du tronc et de celle de la densité du bois : la forme du tronc évolue dans un sens dont les effets sont contraires à ceux de la densité du bois. Formaliser conjointement ces deux dynamiques permet d’augmenter sensiblement le pouvoir explicatif et la robustesse du modèle.
Les paramètres du modèle ne sont pas des valeurs ajustées constantes, mais sont des fonctions elles-mêmes des variables de base (diamètre, la hauteur totale). Ainsi, les paramètres varient d'un arbre à l'autre et reflètent la variabilité spatiale et temporelle de la forme des arbres. Les ajustements font appel à la modélisation bayésienne. Ces modèles sont génériques à l’échelle européenne, cherchant à minimiser l’erreur sur un important gradient de structure, composition et mode de gestion forestière. Ils bénéficient d’une très grande couverture géographique : France, Allemagne, Danemark, Autriche, Roumanie, République Tchèque. La pertinence locale de ces modèles reste à analyser. L'analyse des déviations locales fournira une nouvelle perspective sur la variabilité de l’allométrie et à terme, des facteurs qui la contrôlent ou qui lui sont le mieux corrélés : la densité (relative), la hauteur totale, le rapport moyen de longueur de couronne sur la hauteur totale.
La variabilité spatiale de la densité du bois reste mal connue en dépit de son intérêt évident pour les estimations de biomasse et pour les aspects concernant la qualité du bois. La densité du bois est désormais considérée comme l’un des principaux traits des essences, avec par exemple la hauteur maximale, la longévité, la surface spécifique des feuilles (Szenson et Enquist 2007, Kattge et al. 2009). La variabilité spatiale reste en échange à la fois très importante quantitativement, et se double d’une variabilité individuelle locale considérable demeurant non expliquée. Connue depuis longtemps, elle a motivé l’introduction d’un effet individuel aléatoire dans les modèles de densité (ex. Guilley et al. 1999, Bouriaud et al. 2004 et 2015). Si la quantification de la variabilité spatiale à grande échelle de la densité du bois va devenir possible grâce aux mesures réalisées dans le projet XyloDensMap, en revanche, elle n’apporte pas de réponse à l’échelle locale. Combler le manque de connaissances sur cette échelle intermédiaire apporterait d’une part une
amélioration à la modélisation de la densité, à l’estimation de la productivité, et d’autre part des informations sur la physiologie de la croissance.
1.1.2 Tester des méthodes de calibration locale
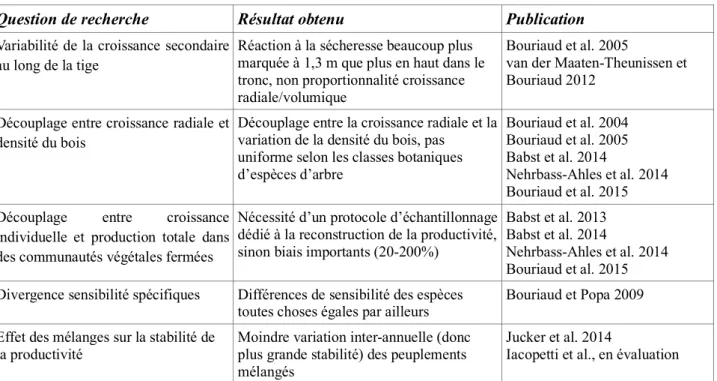
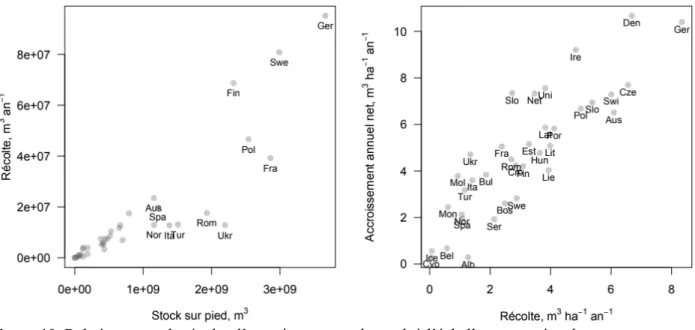
La volonté d’augmenter la pertinence locale des mesures d’inventaire, faisant écho aux demandes toujours plus nombreuses des acteurs du secteur forestier pour des estimations à l’échelle de la ressource, peut se faire par la calibration locale des modèles de manière à minimiser leurs biais. A cette échelle les variations locales de l’allométrie peuvent compromettre la validité des modèles. Les modèles globaux développés sur des jeux de données nationaux peuvent en effet ne pas avoir de biais global mais se trouver trop loin des paramètres moyens d’une zone particulière (Zhang et Shi 2004), donc générer des biais locaux importants (Figure 2). Parmi les facteurs en cause pourrait figurer la diversité spécifique et la très probable influence du voisinage spécifique (Forrester et Bauhus 2016), les mélanges d’espèces ayant une influence certaine sur le développement des couronnes, l’élagage et donc la forme du tronc (Forrester et al. 2017).
Les erreurs de prédiction du volume liées à l’utilisation de modèles allométriques ont longtemps été négligées car supposées s’annuler à l’échelle de la placette mais leurs effets sont en réalité non négligeables (McRoberts et Westfall 2014, Bouriaud et al. 2015). La calibration locale des modèles allométriques paraît être de plus en plus réaliste et efficace, à mesure que les covariables les plus influentes (ex diamètre du tronc à une hauteur fixe ou variable) sont de plus en plus faciles à acquérir.
Figure 2. Biomasse aérienne totale fonction du diamètre à 1,3 m pour différents groupes de données (ici, des zones de provenance régionales) pour l’épicéa en Roumanie.
Les modèles basés sur de grands jeux de données tiennent compte de la variation régionale, formant des groupes de données (correspondant aux sites de provenance des données). La courbe globale peut être localement très biaisée (droite) pour un groupe donné, d’où l’intérêt de calibrer localement.
Dans les inventaires forestiers, les modèles actuels ne sont pas calibrés localement. Les arbres dits tarif utilisés dans certains échantillonnages (tels que l’inventaire forestier Suisse) sont une forme simple de calibration dans laquelle les paramètres des fonctions sont estimés à partir d’un nombre restreint d’arbres. La généralisation au peuplement de la placette peut faire appel à des méthodes en plusieurs stades. Mais les
méthodes de mesure des caractéristiques complémentaires ont beaucoup évolué lors des dernières années et permettent désormais d’obtenir des mesures utiles à des calibrations plus poussées.
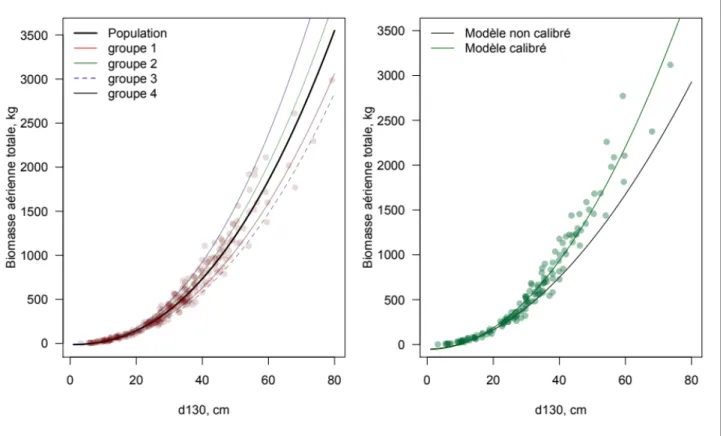
La calibration locale des équations allométriques est une solution efficace pour tenir compte et compenser les variations spatiales de l’allométrie (figure 2). Le projet DIABOLO a montré les perspectives et limites des bases de données actuelles qui semblent déficitaires en ce qui concerne la biomasse des branches ou des parties inférieures au seuil de comptage, ainsi que la densité du bois. Bouriaud et al. 2019a montrent le potentiel de la calibration dans le cas de modèles complexes ayant plusieurs effets aléatoires, et soulignent en particulier l’efficacité de la calibration bayésienne. La méthodologie proposée introduit des changements dans les approches de l’allométrie à trois niveaux différents : i) dans la philosophie et la méthode de l’ajustement du modèle global, lequel doit offrir les paramètres nécessaires à sa calibration, ii) dans la méthode de calibration locale et iii) dans les protocoles de mesures de terrain nécessaires à l’obtention des jeux de calibration et de validation. Ce dernier point aura donc à terme un retentissement probable sur les mesures réalisées sur le terrain par des équipes d’inventaire.
Figure 3. Principe de l’ajustement et de la calibration locale des équations allométriques.
L’ajustement implique un modèle (ici, le volume du tronc, Vstem) ayant des effets aléatoires (groupes, notés u) correspondant aux sites de provenance des données. Deux modalités de calibration sont possibles : le calcul local des effets aléatoires ou la calibration Bayésienne (cf. Bouriaud et al. 2019b)
Le scanner terrestre laser (noté TLS) pourrait ici trouver une application particulièrement efficace en offrant des jeux de données permettant à la fois de quantifier les erreurs de prédictions mais aussi de réaliser une calibration locale des modèles (base de données calibration, figure 3). Ces jeux de données pourraient être limités à une partie des compartiments (les branches fines demeurent un compartiment très difficile à mesurer) en raison de difficultés liés à cette approche.
En conclusion : les nouvelles méthodes d’ajustement des modèles allométriques permettent de produire des formes plus adaptatives et qui permettent une calibration locale. Les données nécessaires à l’ajustement de tels modèles sont encore déficitaires, malgré la relative parcimonie de ces modèles. Des méthodologies pour obtenir des jeux de calibration et de validation doivent être développées, et mises en œuvre. Le TLS devrait permettre d’obtenir non seulement un complément de données pour l’ajustement mais aussi, ou surtout, des données permettant de procéder à des calibrations locales des modèles. Les travaux proposés portent à la quantification des erreurs des modèles existants, à relier les déviations à des caractéristiques de peuplement faciles à mesurer, à déterminer l’importance de la calibration et l’approfondissement des connaissances portant sur la variabilité spatiale des allométries. Après la contribution pionnière de l’inventaire forestier français depuis 2010, d’autres pays s’engagent désormais de façon très pro-active dans l’utilisation du TLS en inventaire forestier (Finlande, Annika Kangas ; Suisse, Urs-Beate Brandli), dans la logique collaborative des inventaires européens. On peut donc envisager une contribution décisive de cette technologie dans un horizon de 5 à 10 ans, au champ des allométries de volume et de biomasse aériens des forêts. Compte tenu de l’investissement de collègues européens et Nord-Américains un projet post-doctoral réunissant plusieurs partenaires serait souhaitable.
Les mesures destructives (nécessaires pour fournir les données de référence) entreprises avec le laboratoire BEF de l’INRAE (Laurent Saint-André) en continuité de DIABOLO devraient permettre d’en apercevoir le potentiel dans le cas d’applications locales. Une thèse de doctorat en collaboration pourrait permettre de déterminer la pertinence locale de ces modèles et d’avoir la base empirique nécessaire pour développer localement une série de modèles de volume et de biomasse, et de les décliner y compris pour des espèces pour lesquelles il existe très peu de données à l’heure actuelle. L’intérêt pour la diversité des espèces met en effet l’accent sur des espèces qui étaient jusqu’ici très largement délaissées.
Proposition pour une thèse de doctorat
Sujet : Variabilité spatiale de l’allométrie des essences principales : quantification, déterminisme, conséquences pour les estimations de volume et de biomasse. Compensation par la calibration locale.
Hypothèses : La variabilité spatiale de l’allométrie provoque des biais d’estimation. Le développement des arbres est influencé par une multitude de facteurs mais les plus importants peuvent être appréhendés par une liste courte de caractéristiques mesurables (ex. h/d moyen). La calibration locale permet d’obtenir des modèles pour des essences pour lesquelles les données sont très rares.
1.1.3 Renforcer les aptitudes dynamiques des prédictions de biomasse aérienne : allocation et allométrie
L’allocation est définie comme la répartition de la croissance, et par conséquent des ressources nécessaires à la croissance : eau, nutriments et photosynthétats, incluant ainsi les coûts de construction et d’entretien (Poorter et al. 2012). Elle représente le résultat de transferts et déplacements de matière, à distinguer de la distribution, laquelle représente les proportions de biomasse observées à un moment donné.
Comme montré en introduction de ce premier axe, la croissance se déduit essentiellement par différence de stock à deux moments, et les modèles de prédiction représentent mal la dynamique, et ont une forme implicite de constance de l’allocation, laquelle reste très peu étudiée.
Distribution et allocation entre les parties aériennes des arbres sont mal connues : d’une manière générale le compartiment aérien le mieux documenté est le tronc. Sur un pas de temps court, la croissance du tronc peut être approchée avec un niveau de précision satisfaisant par des mesures simples de diamètre. Mais mesurer la
croissance primaire et secondaire des branches reste problématique. Le modèle allométrique le plus discuté est le modèle WBE (West Brown Enquist, Brown et West 2000, Enquist 2002) du fait de son universalité théorique, qui est d’ailleurs réellement allométrique par conception : la croissance relative des branches est représentée comme une fonction de la croissance totale de l’arbre, et implique une proportion stricte entre le rayon d’un segment et sa longueur quel que soit le niveau d’observation dans l’arbre. De nombreuses observations montrent que ces contraintes ne sont pas respectées en pratique (Fehrmann et Kleinn 2006, Petit et Anfodilo 2009). Les observations disponibles sont très mal distribuées spatialement, généralement concentrées sur peu d’arbres et peu de sites, et il est impossible d’en tirer plus que la contradiction par ailleurs attendue du modèle WBE. Continuer l’effort de mesures parait donc indispensable pour aller plus loin dans la connaissance de l’allocation de croissance des parties aériennes des arbres sur de grands domaines.
Le scanner terrestre laser apparait ici comme étant potentiellement une alternative très intéressante pour produire des informations sur tous les éléments de manière non destructive, et parce qu’il permet de quantifier à la fois la croissance primaire et secondaire. Deux types de difficultés apparaissent d’emblée toutefois : d’une part l’obstruction stéréologique des différents compartiments ou arbres, d’autre part la perte de précision des mesures avec la hauteur dans l’arbre. Ces deux défauts des mesures TLS pourraient être compensés par un protocole spécifique : l’obstruction demande d’avoir un nombre de scans plus élevé, de manière à minimiser les fractions des branches non scannées. Des mesures directes de dimensions sur les nuages de point, évitant la complication de la reconstruction, laquelle a un niveau d’incertitude trop élevé, devraient permettre de quantifier l’accroissement des organes par comparaison de scans faits sur des périodes différentes.
Des protocoles de terrain adaptés devraient permettre de surmonter le problème de l’obstruction même pour des arbres de grande taille (Fig. 4). Le deuxième problème est lié au fait que la partie supérieure de la couronne est scannée avec une moindre densité de points que le bas à cause la distance au capteur. Un angle constant entre visées se traduit par une distance entre points de mesure augmentant de manière linéaire avec la distance au capteur. A nouveau, il existe une forme de compensation tirant profit du fait que les lignes de tir successives se recoupent en formant une calotte très dense, qui permet de très bien échantillonner les branches fines du moment qu’elles sont situées à la verticale du capteur (Fig. 5).
Pour le moment les efforts d’estimation d’accroissement à partir de données TLS sont très rares (Sheppard et al. 2016) bien que certains tests comme le suivi FACE (Free-Air CO2 Enrichment) donne des résultats
satisfaisants (Eric Casella, com. pers). Les difficultés liées à l’exploitation des nuages de points et à la reconstruction des arbres et de leur volume semblent encore limiter fortement la précision, donc l’utilité, de cette approche. En attendant une percée méthodologique autour des algorithmes de traitement des données TLS, les nuages de point de qualité demeurant le facteur le plus limitant dans l’effort de d’analyse de l’allocation, un projet visant à regrouper les acteurs Européens (ex. E. Casella, UK ; T. Pitkänen & A. Kangas, FI ; F. Babst, CH) ou Nord-Américan (B. Strimbu, USA) avec des protocoles standardisés pourrait permettre de constituer une base de données orientée vers la quantification de l’accroissement et sa répartition.
Figure 4. Nuages de points TLS. Ce nuage montre (à gauche) un très grand hêtre (hauteur totale d’environ
40 m) dont on voit la couronne très complexe et (à droit droite) un zoom sur une partie de la couronne située à environ 30 m de hauteur.
Cet effort va aussi dans le sens d’une meilleure intégration avec les cycles biogéochimiques, la partie croissance ayant un rôle central dans les cycles (Janssens et al. 2001). Pousser les estimations de croissance vers la minéralomasse (Legout et al. 2020), pour un sous-échantillon dans un premier temps, permettrait de dimensionner l’effort nécessaire pour élargir les perspectives de ces études aborder le champ des cycles biogéochimiques.
Figure 5. Représentation schématique de la densité des points de mesure d’un TLS.
La calotte étant à la croisée des transects d’échantillonnage, la densité locale est beaucoup plus forte au-dessus de l’appareil comme le montre l’histogramme de densité.
En conclusion :
L’estimation de la productivité nécessite d’utiliser en différentiel des modèles ajustés sur des données statiques. Sur des périodes de temps courtes cette approximation est sans doute justifiée, mais en pratique, pas du tout documentée ou testée. Toutefois il existe des indices suggérant une dynamique de l’allocation aux compartiments aériens boisés non négligeable.Le manque de connaissances quantitatives sur l’allocation pourrait être compensé par le couplage de différents types de mesures de suivi. Ces mesures fourniraient non seulement des données nécessaires pour la modélisation, elles apporteraient aussi des connaissances fondamentales sur l’allocation et la plasticité des arbres. L’effort de mesure est très important, doit mettre en jeux plusieurs méthodes, et des protocoles doivent être conçus et testés afin d’obtenir des jeux de données standards de qualité suffisante pour permettre d’alimenter des travaux de modélisation. L’IFN étant construit pour une revisite des placettes à 5 années, l’allongement de la fenêtre temporelle ne peut se faire qu’en mesurant en dehors de l’échantillon annuel standard de l’IFN mais doit se baser sur son réseau et toutes les mesures passées pour constituer une collection de placettes. Cet effort de mesures accompagnera celui développement des méthodes TLS, s’agissant de l’individualisation des arbres, du filtrage des nuages de points et de la reconstruction géométrique.
Proposition pour une thèse de doctorat
Sujet : Variations dans l’allocation aérienne des arbres, plasticité et réaction aux conditions de croissance.
Hypothèses : Les erreurs de prédiction des accroissements sont corrélées aux conditions de croissance, notamment météorologiques. Les arbres procèdent à des ajustements dans l’allocation qui permettent d’atténuer leurs effets négatifs.
L’objectif principal est de quantifier la magnitude des ajustements de l’allocation pour quelques essences principales feuillues. Comme suggéré un assemblage de technologies complémentaires permettrait à la fois de limiter les risques : micro-dendromètres pour l’accroissement radial des branches sur des arbres mesurés de manière répétitive à l’aide du TLS. Les sites d’étude pourraient être choisis à base d’un examen des scans récents des placettes IFN, et s’efforcer de trouver des sites représentant des conditions de croissance contrastées, notamment en matière de réserve hydrique du sol.
1.2 Application dans les estimations de l’inventaire forestier national 1.2.1 Améliorer les estimations de productivité dans l’inventaire
Les aspects suivants, portant sur l’amélioration des estimations de productivité, sont au premier abord assez techniques mais très riches d’enseignements car ils documentent et quantifient les mécanismes fondamentaux du développement des forêts : recrutement, croissance, mortalité et récolte. En tant que petite résultante de composantes beaucoup plus grandes, la productivité demeure un objet difficile à inventorier et quantifier. Compte tenu de l’augmentation de la pression exercée sur les forêts en vue de la mobilisation des ressources forestières, l’estimation des flux par l’inventaire est un thème de recherche important. Appuyés sur les mesures de l’inventaire forestier national (IFN) ces travaux ont pour objectif de contribuer à l’effort permanent d’amélioration ou optimisation de l’enquête. Le positionnement et le contenu du projet sont appuyés largement sur l’IFN et les axes de recherches du LIF, présentés dans la section Synthèse en fin de document.
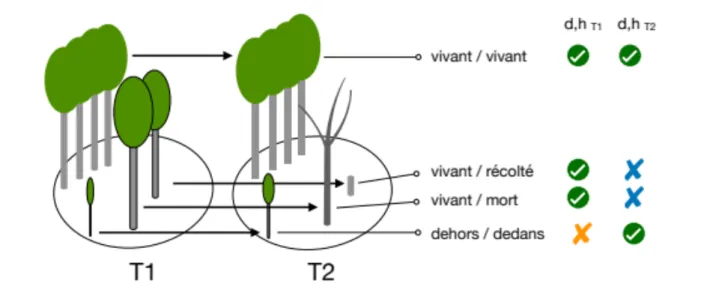
L’échantillonnage terrestre de l’inventaire forestier français se base sur un échantillon de placettes semi-permanentes totalement renouvelé. En effet, pour une année donnée, environ la moitié des placettes sont mesurées pour la première fois tandis que l’autre moitié représente les placettes installées cinq ans auparavant. Les flux sont estimés à partir des mesures effectuées sur les placettes re-visitées, dites placettes de retour. Une comparaison d’inventaire permet alors d’estimer les flux principaux du bilan de matière : accroissement, recrutement, coupes et mortalité. Ces composantes ne sont pas mesurables directement mais recèlent des estimations indirectes, le calcul de la production simple direct n’étant possible que pour les seuls arbres vivants aux deux moments de l’inventaire. Pour tous les arbres mesurés à une seule occasion, soit lors du premier (arbres morts ou récoltés) soit lors du deuxième (recruts, oublis, fraction devenue accessible), l’accroissement ne peut être obtenu que par modélisation (voir schéma en fig. 6). Pour une fraction des arbres mesurés aux deux occasions, un saut dans les données indique une erreur de mesure. Pour tous ces arbres, le calcul de l’accroissement se base sur des estimations et non sur des mesures. Il existe pour toutes ces situations deux solutions de calcul : une modélisation faisant appel à un déterminisme (i.e. des régressions) et qui sont presque toujours paramétriques, et des imputations qui n’impliquent pas de causalité et ne sont pas paramétriques. Le problème du modèle est qu’il est développé à base de jeux de données complets, et que donc d’emblée se pose la question de la représentativité des sous-domaines utilisés pour leur ajustement (van Deusen 1997).
Les modèles de croissance sont a priori les moins adaptés aux inventaires (pas seulement nationaux) à cause du biais qu’ils introduisent. En effet, une prédiction d’un modèle de croissance sera strictement positive (c’est d’ailleurs aussi le cas des largeurs de cerne), ce qui est logique mais n’est pas compatible avec les estimations de croissance réalisées par comparaison d’inventaire et pour lesquelles les croissances apparentes peuvent être négatives (essentiellement à cause des imprécisions de mesure). Les imputations en revanche sont capables de prédire des estimations négatives, qui respectent ainsi la distribution des mesures réelles.