Pollen-based climate reconstruction techniques for late Quaternary studies
Texte intégral
Figure
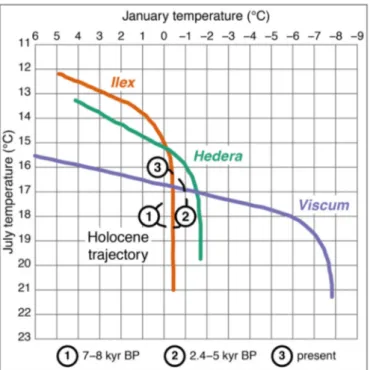



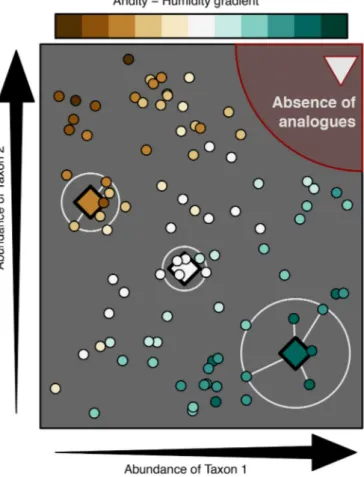


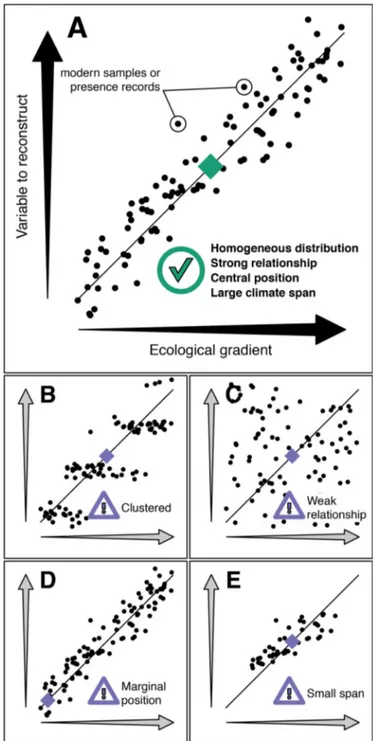

Documents relatifs
Study of uranium(VI) and radium(II) sorption at trace level on kaolinite using a multi-site ion exchange model.. Jacques Ly,
Here we report mitochondrial DNA identification and haplotype richness estimation, using 239 pangolin scale samples from two confiscations in Hong Kong.. We found a total of
When we replace the pushers with high-friction pushers (pushers with rubber coating), the planner estimates that the desired horizontal object motion lies inside the motion cone for
Department of Mathematics, Massachusetts Institute of Technology, Cambridge, Massachusetts 02139, USA Department of Electrical and Computer Engineering, National University
Mots clés: Mangue (Mangifera indica L.), arôme, composés volatils, glycosides, précurseurs d'arôme, traitement thermique, traitement
Under the reference plant invasion scenario, the population sensitized to ragweed at the European level is estimated to increase from 33 million (range: 27 million–38 million) to
An important concept in regional analgesia is that the duration of the applied stimulus (temporal summation) and the area where the stimulus is applied (spatial summation)
In contrast, in a study, a daily dose of 16 g of inulin – oligofructose (50:50, v/v) in obese women led to changes in the gut microbiota composition that were of a magnitude similar
