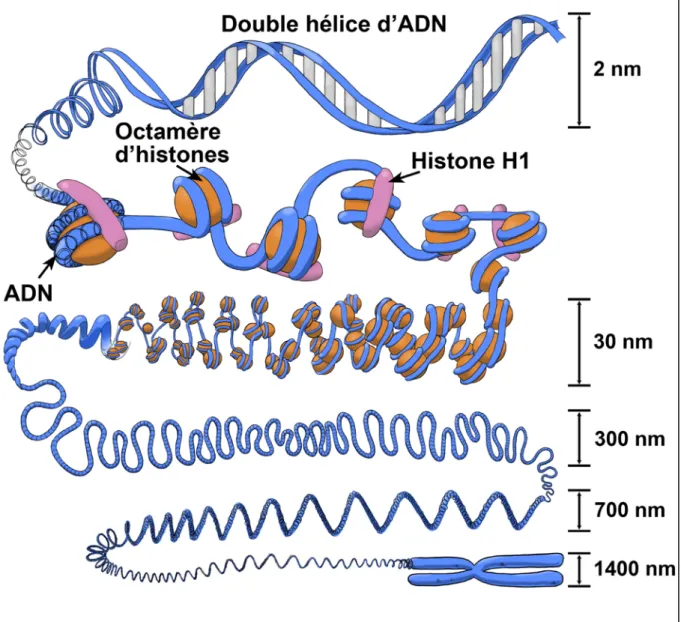
Méthylation de l'ADN, phyto-oestrogènes et cancer du sein et de l'ovaire
Texte intégral
Figure






Outline
Documents relatifs
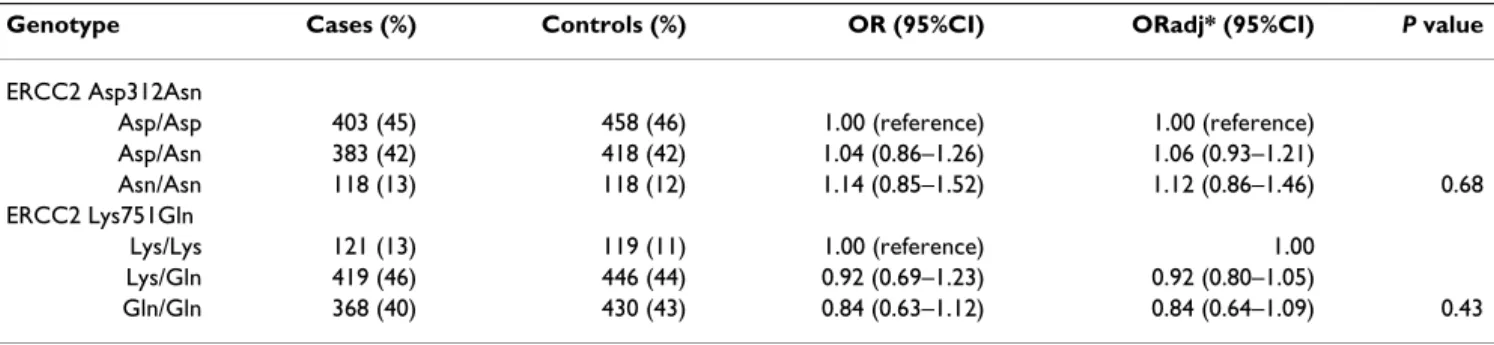
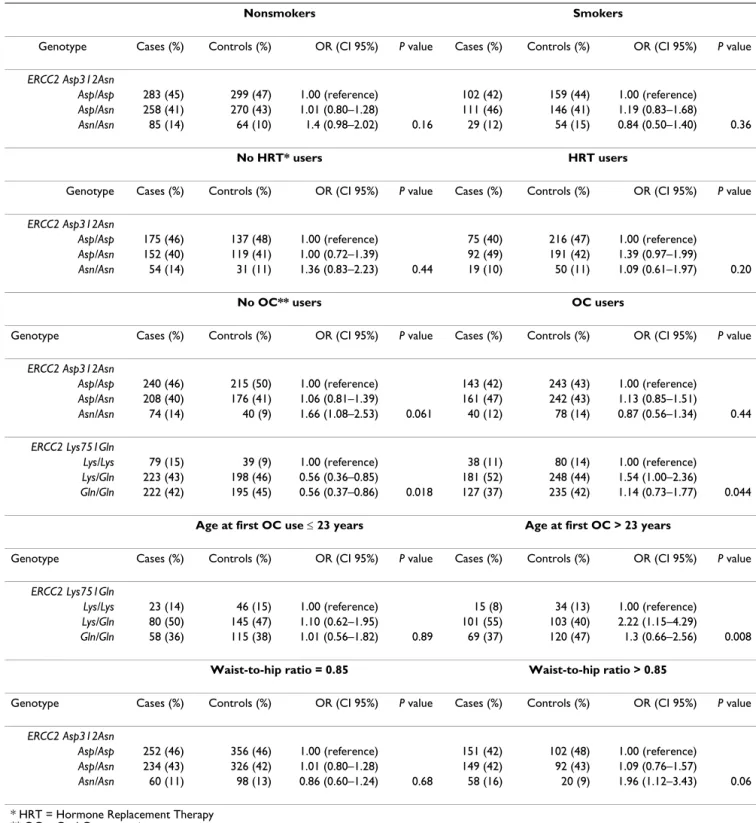
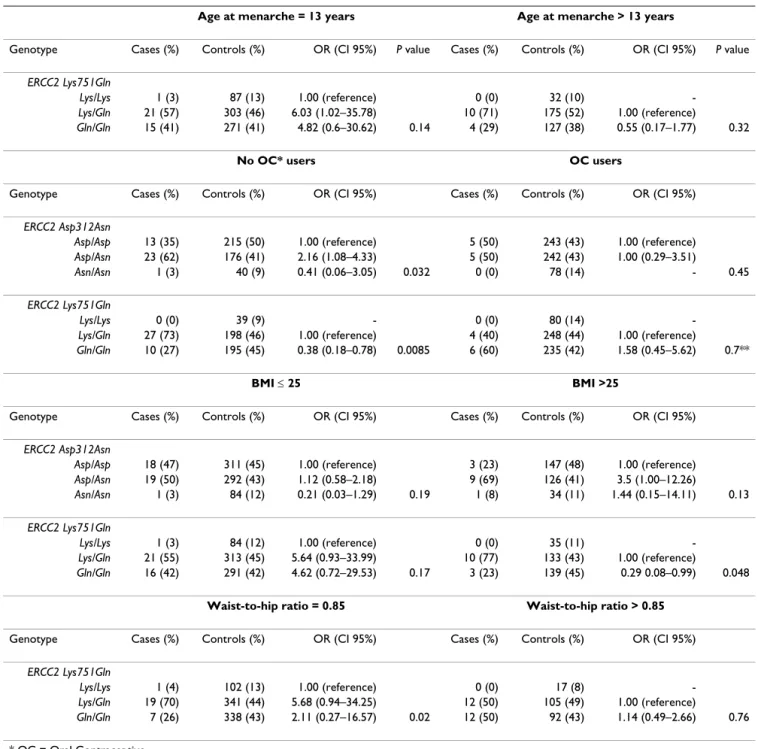
Dans certaines situations la présence simultanée de deux variants de classe 1 (POLYMORPHISME) des gènes BRCA1 et ou BRCA2, sont responsables d’une augmentation du
(Tumorectomie monobloc jusqu’au plan pectoral et décollement profond de la glande
Notons que certaines des mutations des gènes BRCA1 et BRCA2 sont uniques dans la population maghrébine (Afrique du Nord) et ont déjà été signalées dans d'autres
Une surveillance continuelle de la fonction cardiaque chez les survivantes d’un cancer du sein asymptoma- tiques n’est actuellement pas recommandée 16 ; toutefois, les
Liane à fr. charnu formé de n drupéoles.. Gynécée à n C libres, uniovulés et ovaire infère non adhérent au réceptacle floral, après fécondation chaque C se
Tableau B-3 Recommandations des guides de pratique clinique sur l’utilisation de la TEP-TDM pour l’orientation thérapeutique et l’évaluation de la réponse au traitement du
les jeunes filles âgées de 11 à 13 ans révolus à la première dose, pour lesquelles le vaccin GARDASIL a été choisi pour la vaccination HPV, soient vaccinées selon un schéma à
- Les gènes de classe B, associés aux gènes de classe A contrôlent la mise en place des pétales, ici la diminution de l’expression de RhAP3 induit la présence de plus de pétales.
