T
T
H
H
È
È
S
S
E
E
En vue de l'obtention du
D
D
O
O
C
C
T
T
O
O
R
R
A
A
T
T
D
D
E
E
L
L
’
’
U
U
N
N
I
I
V
V
E
E
R
R
S
S
I
I
T
T
É
É
D
D
E
E
T
T
O
O
U
U
L
L
O
O
U
U
S
S
E
E
Délivré par l'Université Toulouse III - Paul SabatierDiscipline ou spécialité : NEUROSCIENCES
JURY
Pr. Jean-Michel LASSALLE Président Dr. Valérie DOYERE Rapporteur
Dr. Bruno POUCET Rapporteur Dr. Alain MARCHAND Directeur de thèse
Ecole doctorale : CLESCO Unité de recherche : CNIC et CRCA
Directeur(s) de Thèse : Alain MARCHAND et Martin GIURFA Rapporteurs : Docteur Valérie DOYERE et Docteur Bruno POUCET
Présentée et soutenue par Frédéric ESCLASSAN Le 19/12/2008
Titre : NATURE ET SUBSTRAT NEUROBIOLOGIQUE DU CONDITIONNEMENT DE TRACE :
par le Pr Jean‐Michel LASSALLE. Ils ont donné de leur temps dans l’évaluation de ce rapport et ont manifesté un intérêt particulier à ce travail notamment le jour de la soutenance à travers un riche débat que j’ai personnellement beaucoup apprécié.
Il serait impardonnable de ma part de ne pas sincèrement remercier la personne qui a sûrement le plus souvent, durant ces quatre dernières années, fait les frais de mon sale caractère et de mon obstination j’ai nommé le Dr Alain MARCHAND aussi connu sous le nom de « Mon directeur de Thèse » ou encore « chef ». A titre personnel Alain je tenais vraiment à te remercier puisque tu as toujours répondu présent aux moments clefs pour m’apporter un soutien aussi bien scientifique que moral.
Je voulais aussi remercier le Dr Georges DI SCALA directeur du CNIC mais aussi chef de l’équipe au sein de laquelle j’ai effectué les travaux qui vont suivre. Une double casquette dure à porter et surement parfois bien ingrate que tu as pourtant su garder fermement attaché sur la tête. Georges, ton « souci », ton scepticisme et ton amour des mots précis m’ont poussé à évoluer et ; ne jouons pas sur la fausse modestie ; à sans cesse m’améliorer. Pour cela, je te suis redevable. Comment ne pas remercier mon collègue de chaque jour, celui sans lequel ce travail ne serait certainement pas ce qu’il est aujourd’hui : le Dr Etienne COUTUREAU. Entre discours scientifique et séries d’opérations il est toujours bon de glisser une bière… ou un barbecue. Merci à toi d’avoir était aussi disponible. Ton humour (non non je ne vais pas donner d’exemple ne t’en fait pas) va énormément me manquer.
Je remercie aussi le dernier arrivé des titulaires de l’équipe le Dr Jean‐Rémi PAPE. Bien que tu sois arrivé tardivement dans l’équipe tu as su t’adapter à des thématiques de recherche pas forcément simple à maitriser lorsqu’on n’a pas une formation ES Psychologie Expérimentale. Pourtant je ne t’ai jamais vu baisser les bras à la vue d’un tableau rempli de X‐/AB+/A‐/Y+. Merci à toi aussi pour ton soutien.
Il est temps à présent que je remercie mes compagnons de galère, qui continuent encore à se réunir, mais à présent sans moi, au sein de notre sacro sainte « réunion d’équipe » du vendredi midi je veux bien sûr parler des thésards de l’équipe « Fonctions exécutives, circuits neuronaux et développement ».
Aurore la pro des X‐/AB+/A‐/Y+ (conf. plus haut). Le petit cœur avec un caractère de cochon. Il est difficile de trouver les mots justes pour te remercier tellement, les mots semblent insuffisants pour dire à quel point ton soutien et ta présence ont été pour moi indispensables durant tout ce temps. Ta force de caractère et de persévérance devrait être un exemple pour tous ceux qui t’entourent puisqu’après tout ce temps et toutes ces galères que tu as traversées c’est grâce à toi que demain nous pourront dire : « bien sûr que les rats sont capables de faire de la ré‐évaluation
sans problème tant votre force de travail et votre attrait pour la science (et la rigueur qui va avec) sont fort. Bon courage et merci pour tout.
Enfin une petite pensée pour une petite espagnole du tonnerre la petite Maria qui a fait quelques temps partie de cette équipe avant de finalement partir réaliser sa thèse dans sa ville natale de Madrid (et oui je sais combien il est difficile de quitter le Sud). Ton accent et ta bonne humeur m’ont manqué et me manquent encore. Merci d’avoir été là. Après avoir remercié les collaborateurs les plus proches de ce travail il me faut maintenant remercier tout les autres qui bien que menant leur propres barque on bien voulu m’accorder de leur temps et partager avec moi les nombreux moments inoubliable que j’ai passé à Bordeaux. N’en déplaise à la bienséance je vais cette fois‐ci commencer par remercier les étudiants du CNIC (passé et présent). Edith. Entre le thym et le Cumin se dégage une douce odeur de gâteau sucré attestant de la présence d’un « sexy dolup » qui reste toujours « fraiche comme un lardon ». Ce soir avec didit nous irons surement cocher les cases d’une grille d’euro million avant bien sûr d’aller descendre une Carlsberg. Nous gagnerons c’est sûr puisque nous avons garé la voiture sur notre place rituelle. Nous pourrons donc acheter une île sur laquelle nous bâtirons ZANAX ® notre maison de 50 pièces. Edith pourra alors enfin passer ses journées à faire le ménage pendant que je serai dans un hamac. Ed viendra me voir tout sourire je lui dirai alors «tu sais ma puce faudrait vraiment que tu fasses quelques chose pour tes cheveux»… Elle fera un sourire crispé et partira à la recherche d’une répartie cuisante qui arrivera… trop tard… Edith tu comprendras potentiellement tout ça et tout ce que ça veut dire pour moi/toi/nous quoi.
Au tour des trois anciens qui pour moi restent indissociables, Stéphane, Ludo et Pierre. Vous avez su nous montrer la voie à suivre et avez réussi à supporter nos plaintes et nos sauts d’humeur (j’écris « nos » par honte d’écrire « mes »). Les années passent et vous restez toujours pour moi l’objectif à atteindre. Pierre merci de m’avoir fait comprendre que la biologie moléculaire c’est l’ADN tout ça tout ça... Stéphane merci de m’avoir laissé croire que je pouvais dire des trucs pas trop con parfois. Ludo merci d’avoir été mon modèle tout simplement.
Je demande aux personnes dont le nom va suivre de s’avancer d’un pas : François (mon basque, ça serait le pied de pouvoir partager à nouveau mon bureau avec toi grand), Azza (ma azzazou, tes blagues et ta bonne humeur me manquent terriblement), Seb (le mec le plus spontané au monde, c’est quand que tu viens me voir parigot ???), Aurélie (la pile électriques aux 120 casquettes mais toujours disponible pour une bière), Stephanie G.B. (la petite maman, je te souhaite tout le bonheur du monde), Stephanie A.Q. (l’autre maman, je suis lié à toi pour la vie par la date du 19 Décembre 2008), Yves (le seigneur des terres de Castillon la bataille) et Maude (l’autre toulousaine, celle qui a partagé avec moi les affres du DEA).
course) ou plus simplement d’un pichet de bière !
Je tiens aussi à remercier du fond du cœur l’ensemble du personnel technique qui facilite et rend possible le travail au quotidien que cela soit dans les animaleries, les salles d’histologie, les locaux techniques et informatiques ainsi que l’intendance et le secrétariat. Un remerciement particulier à Maïté ROY qui avant de nous quitter a fortement contribué à la formation technique de nombreux étudiants dont j’ai eu la chance de faire partie.
Enfin merci à l’ensemble des chercheurs titulaires du CNIC trop nombreux pour être nommé individuellement mais qui par le biais de nombreuses discutions formelles et informelles ont participé a la construction de mon esprit critique durant ces 4 année.
Enfin je remercie aussi mes amis castrais, albigeois, toulousains, ruthénois, mazamétains, montpelliérains et autres qui malgré la distance restent auprès de moi.
Et bien entendu merci à ma famille qui n’a eu de cesse de croire en moi, je vous dédie ce manuscrit car sans vous il n’aurait jamais pu voir le jour. MERCI pour votre présence et pour vos encouragements. Merci tout d’abord à mes parents qui ont toujours cru en moi, même dans les moments où je perdais confiance. Merci à Mes sœurs Kinou et Zaza qui m’ont longtemps poussé vers l’avant. Sans vous je serais certainement encore en train de bosser mes verbes irréguliers. Merci a mes beaux‐fréres Dada et Jéjé, Médecins et Docteurs Je n’ai pas eu l’envie de vous surpasser mais votre proximité m’a poussé à me dépasser. « L'envie veut abaisser et l'émulation égaler. L'une
s'afflige des succès, l'autre y aspire. Celle‐là est jalouse de tout mérite et l'autre en est ambitieuse. ».
Enfin Merci à mes six petits anges Lena, Emma, Elise, Colin, Cloé et Milan. Vous observer grandir est une de mes plus grandes joies. Merci pour ces moments passés avec vous et merci pour toutes vos preuves d’amour.
SOMMAIRE
INTRODUCTION GENERALE
I- Cadre général du travail de thèse.
1
II- Conditionnement de Trace : Définition et aspects cognitifs. 3
II- Aspects théoriques du conditionnement de Trace.
10
III- Aspects neurobiologiques.
18
IV- Principaux objectifs de ce travail de thèse.
26
CHAPITRE I :
Sélection des paramètres du protocole expérimentaux
INTRODUCTION
I- Effets de manipulations de l’intensité du SI.
27
II- Les critères pris en comptes pour la mise en place du protocole
de Trace.
28
III- Approche expérimentale.
30
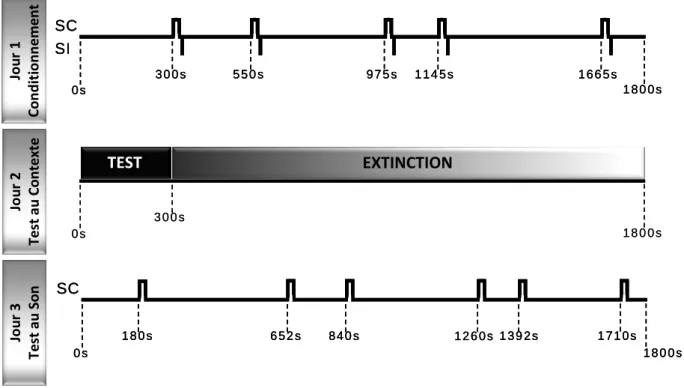
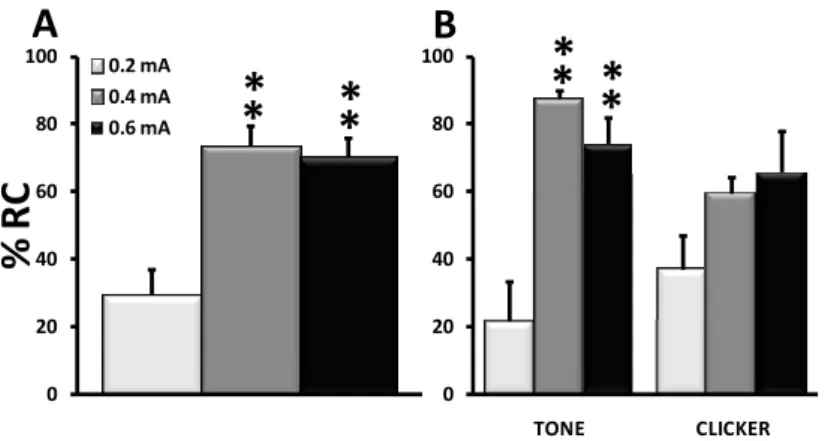
Expérience 1: Effet de l’intensité du SI sur
le conditionnement à un son.
31
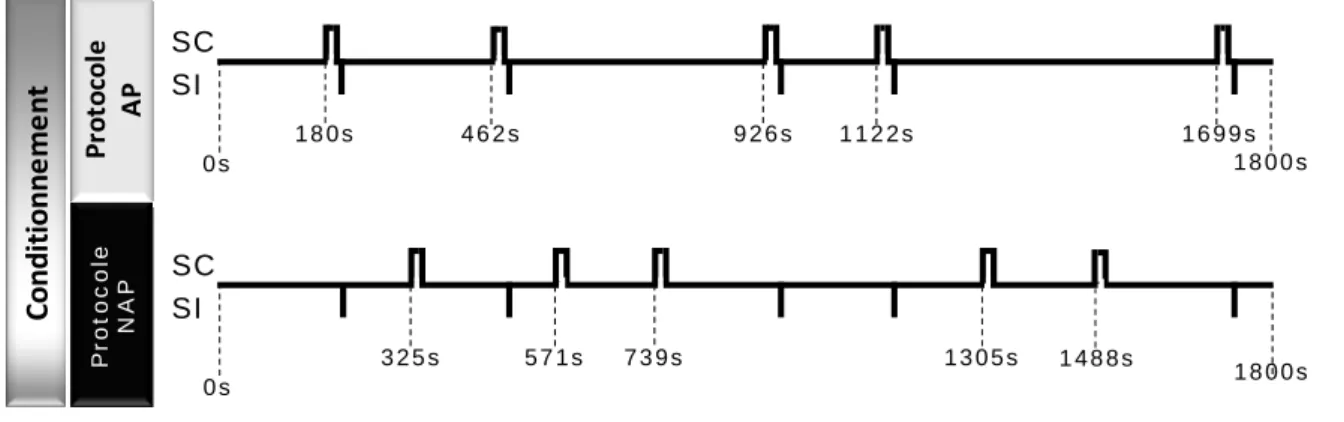
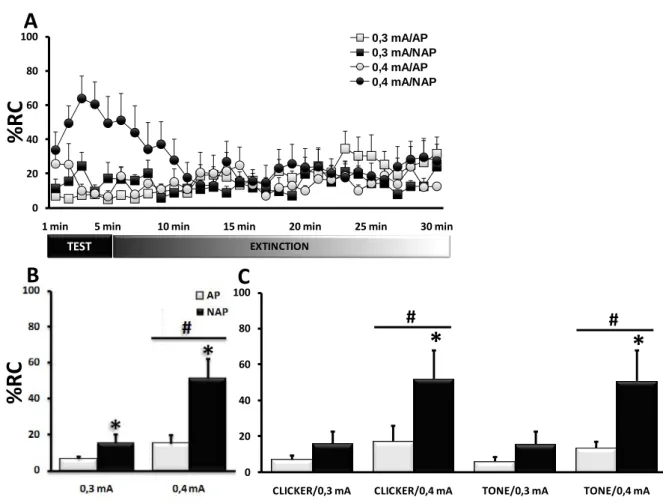
Expérience 2 : Possibilité d’un conditionnement
au contexte par un protocole non apparié.
40
Expérience 3 : Sélection d’un intervalle de Trace
46
DISCUSSION
I- Intensité du SI et Réponse Conditionnée.
55
III- Conditionnement avec intervalle de Trace variable.
56
II- Nature du SC et Réponse Conditionnée.
57
CHAPITRE II :
Composante contextuelle du conditionnement de Trace
«Implication de l’hippocampe et de ses sous-régions»
INTRODUCTION
I- L’hippocampe.
63
II- Approche expérimentale.
74
Expérience 1: Lésion totale de l’hippocampe.
75
Expérience 2: Inactivation des sous-régions hippocampiques.
84
DISCUSSION
I- Aspects méthodologiques.
92
II- L’inactivation de l’hippocampe ventral
reproduit les effets de la lésion totale.
93
III- Contribution de l’hippocampe dorsal au
conditionnement de peur associé au contexte et de Trace.
97
IV- Relation entre conditionnement de trace et
conditionnement contextuel.
98
V- L’hippocampe comme support de la « Trace ».
99
CONCLUSION
102
CHAPITRE III :
Conditionnement de Trace et Trace persistante
«Rôle du cortex entorhinal et du cortex préfrontal médian»
INTRODUCTION
I- Le Cortex Préfrontal.
103
II- Le cortex entorhinal.
112
III- Approche expérimentale.
116
Expérience 2: Lésion du cortex entorhinal.
124
DISCUSSION
I- Effets de la lésion du CPFm.
131
II- Effets d’une lésion du CE.
134
CONCLUSION
138
CHAPITRE IV :
Conditionnement de Trace et Trace persistante
«Implication du système cholinergique du cortex entorhinal
lors du conditionnement de Trace»
INTRODUCTION
I- Les neurones à activité persistante.
139
II- Système cholinergique du cortex entorhinal.
142
III- Approche expérimentale.
147
Expérience 1 : Déafférentation cholinergique
du cortex entorhinal.
149
Expérience 2: Blocage des récepteurs muscariniques M1
du CE par la pirenzépine durant le conditionnement.
157
Expérience 3: Blocage des récepteurs muscariniques M1 du CE
par la pirenzépine après le conditionnement.
164
DISCUSSION
I- Absence d’effet de la déafférentation cholinergique.
169
II- Effets sélectifs du blocage des récepteurs M1.
171
III- Rôle de l’acétylcholine dans le conditionnement de Trace. 174
DISCUSSION GENERALE
I- Rappel des principaux résultats.
177
II- Nature et spécificité du conditionnement de Trace.
179
III- Structures et circuits impliqués.
184
IV- Nouveaux mécanismes proposés.
189
CONCLUSION ET PERSPECTIVES
REFERENCES BIBLIOGRAPHIQUES
INTRODUCTION
GENERALE
INTRODUCTION GENERALE
I- Cadre général du travail de thèse.
La totalité des travaux présentés dans ce travail de thèse concerne l’étude des mécanismes impliqués dans la mise en place d’une forme particulière d’apprentissage associatif : le conditionnement de Trace.
Les apprentissages associatifs reposent sur la capacité des animaux, des invertébrés jusqu’aux mammifères supérieurs, à établir des associations entre divers éléments de leur environnement et la survenue d’événement signifiants. Un exemple simple serait la capacité qu’ont les abeilles à associer la présentation d’une odeur à l’obtention d’une solution sucrée appliquée sur leur proboscis. L’association faite par l’abeille entre ces deux éléments induira alors une modification comportementale et la simple présentation de l’odeur sera par la suite capable d’induire à elle seule une extension du proboscis (Bitterman et coll., 1983; Vergoz et coll., 2007).
Afin de faciliter la compréhension et la généralisation de ces phénomènes, l’étude des apprentissages associatifs a donné naissance à un champ lexical qui lui est propre. Ainsi, si on reprend l’exemple précédemment cité, l’odeur présentée à l’abeille serait définie comme étant le "stimulus conditionnel" (SC), c'est-à-dire un stimulus neutre préalablement à l’apprentissage, alors que la solution sucrée ferait quant à elle office de "stimulus inconditionnel" (SI), c'est-à-dire un stimulus ayant une pertinence biologique pour l’animal. L’extension du proboscis après présentation de l’odeur à la suite du conditionnement constitue la "réponse conditionnée" (RC) due à l’association faite entre le SC et le SI.
Selon les théories actuelles du conditionnement classique, il apparaît que la présentation du SC suite à un conditionnement peut évoquer une représentation du SI et que c’est alors cette représentation du SI qui provoquerait l’expression de la RC. En d’autres termes, présenter l’odeur à l’abeille alors que l’abeille a déjà expérimenté que cette odeur précédait l’obtention d’une solution sucrée va provoquer chez l’abeille un rappel de cette solution sucrée, et ainsi entraîner la réponse d’extension du proboscis.
1) Un peu d’histoire.
Les bases théoriques du conditionnement classique ont été posées par Ivan P. Pavlov (1849-1936). Physiologiste de formation, il est avec le psychologue américain John B. Watson (1878-1958) un des fondateurs et piliers du mouvement behavioriste selon lequel la psychologie repose sur l'étude du couple stimulus-réponse et l'adaptation à une situation déterminée. Le behaviorisme, pour lequel les apprentissages font référence à un changement dans le comportement de l’organisme, s’oppose alors au mouvement cognitiviste (Edward C. Tolman 1886-1959) pour lequel les apprentissages reposent sur l’acquisition d’un savoir.
Au cours de ses travaux menés sur le chien et portant initialement sur la physiologie des glandes gastrique et salivaire, Pavlov étudiait la mise en place du réflexe salivaire lors de la présentation d’aliments. Or, au fur et à mesure de ces expériences, Pavlov s’aperçut qu’après un certain nombre de présentations du stimulus alimentaire, ce dernier n’était plus le seul à provoquer chez l’animal une salivation, mais que divers autres stimuli, tels que le bruit induit par l’arrivée de l’expérimentateur apportant la nourriture, étaient eux aussi capables d’induire ce phénomène. Pavlov en conclut qu’un réflexe appris avait été mis en place par l’appariement d’un stimulus neutre et de la nourriture dans la bouche de l’animal. Pavlov, conscient que ce phénomène constituait une forme simple d’apprentissage et que l’apprentissage représentait un mystère scientifique, décida de concentrer ses efforts sur l’étude de ce phénomène. Il réalisa alors rapidement qu’une approche purement psychologique et descriptive se révélerait sujette à de nombreuses controverses. De plus, ce type d’approche était totalement contraire à sa formation de physiologiste. Il décida alors de mener son étude de façon plus expérimentale puisque, selon lui, le cerveau pouvait être considéré et étudié comme n’importe quel autre organe. Ainsi l’apprentissage devrait pouvoir être traité comme n’importe quel autre mécanisme biologique à l’aide de constatations objectives et de protocoles expérimentaux contrôlés.
A partir de ce constat, Pavlov et ses collaborateurs conçurent un grand nombre d’expériences simples afin d’étudier le phénomène d’apprentissage. Ses travaux, toujours menés chez le chien, furent publiés sous le titre "Conditioned reflexes : an investigation of the
physiological activity of the cerebral cortex" (Pavlov, 1927). Ils constituent encore
conditionnement de Trace qui est le sujet de ce manuscrit a été défini par Pavlov dans le cadre de ces études.
2) Les règles gouvernant les associations Pavloviennes.
Dès 1927, un certain nombre de lois sous-tendant la mise en place et la force d’un apprentissage associatif furent énoncées par I.P. Pavlov. Pavlov en énonça trois comme prépondérantes : la contiguïté, la fréquence et l’intensité. Ces trois lois stipulent que l’association entre un SC et un SI va pouvoir être réalisée à partir du moment où ces deux stimuli vont être présenté de façon contiguë (loi de contiguïté) (Gormezano et Kehoe, 1981), qu’ils vont être associés de manière répétée (loi de fréquence) et enfin que leur intensité respective va déterminer la force de leur association (loi d’intensité).
Bien que ces lois soient restées d’actualité durant une trentaine d’années après la mort de Pavlov, R. Rescorla les révisa dans les années 60. Deux des avancées majeures apportées par Rescorla dans ce cadre furent l’ajout de la loi de contingence ainsi que la prise en compte de la compétition entre divers SC dans le modèle formel d’apprentissage qu’il établit (Rescorla et Wagner, 1972).
La loi de contingence stipule que la probabilité de survenue du SI doit être plus forte en présence du SC qu’en son absence. En d’autres termes, il faut que le SC soit "prédictif" de l’occurrence du SI. Ainsi, de manière pratique, il faut non seulement que l’appariement SC/SI soit présenté de façon répétée (on retrouve ici la loi de fréquence), mais aussi que le SI ne soit pas présenté indépendamment du SC. Pour Rescorla, c’est sur le facteur de contingence plus que sur celui de fréquence que repose la capacité du SC à correctement induire un conditionnement (Rescorla, 1968; Miller et coll., 1995).
II- Conditionnement de Trace : Définition et aspects cognitifs.
1) Définition.
Le conditionnement de Trace est une forme particulière de conditionnement Pavlovien lors duquel la présentation du SC et du SI sont séparés par un intervalle de temps "vide"
généralement nommé intervalle de Trace. On oppose généralement le conditionnement de Trace au conditionnement de Délai durant lequel les stimuli sont présentés de manière contiguë. Très tôt, il a été reconnu que la capacité des animaux à établir une association entre ces deux stimuli malgré la présence de cet intervalle de Trace allait à l’encontre de la loi de contiguïté (Guthrie, 1930 ; Razran, 1930). Il était pourtant largement admis que l’existence de la contiguïté entre stimuli est nécessaire à l’établissement de cette association. Ainsi, un des enjeux majeurs quant à la compréhension du conditionnement de Trace consiste à appréhender les mécanismes cognitifs et neurobiologiques qui permettraient de compenser cette violation de contiguïté.
2) Les différentes formes de conditionnement de Trace.
Depuis la première description du conditionnement de Trace par Pavlov, les caractéristiques de cet apprentissage associatif ont été décrites dans de nombreux autres paradigmes expérimentaux parmi lesquels : le conditionnement du clignement palpébral chez l’homme (Hansche et Grant, 1960), la réponse masticatoire conditionnée (Seager et coll., 1999), le conditionnement de la membrane nictitante chez le lapin (Gormezano et coll., 1983) le conditionnement de peur (Kamin, 1965) ou encore l’aversion gustative ou olfactive conditionnée (Garcia et coll., 1966; Ferry et coll., 2006). Ces trois derniers paradigmes expérimentaux sont particulièrement intéressants, tout d’abord parce qu’ils sont à l’origine de la majorité de la littérature actuelle portant sur le conditionnement de Trace, mais aussi et surtout parce qu’il existe une différence fondamentale entre eux en ce qui concerne la durée de l’intervalle de Trace qu’ils autorisent.
a) Conditionnement de la membrane nictitante.
Le conditionnement de la membrane nictitante repose sur l’association d’un stimulus élémentaire (SC) tel qu’un son ou une lumière avec la présentation d’un souffle d’air ou d’un léger choc électrique (SI) au niveau de l’œil de l’animal (la plupart du temps le lapin). Cette association se traduit par la mise en place d’une réponse conditionnée qui consiste en un clignement de la membrane nictitante (une troisième paupière translucide que possèdent certains animaux et qui permet de protéger et d’humidifier l’œil tout en conservant une certaine vision). Il faut noter que, dans ce protocole, la RC quantifiée est identique à la
réponse comportementale manifestée en présence du SI ou réponse inconditionnée (RI). De plus, il a été largement démontré que lors d’un conditionnement de Trace de la membrane nictitante, le pic de RC après présentation du SC se produit peu avant le moment prévu de l’arrivée du choc (Gormezano et coll., 1983). Ceci met en évidence que la composante temporelle du conditionnement de Trace fait partie intégrante de l’association acquise (Marchand et coll., 2004). Ce phénomène représente une forte contrainte pour les modèles d’apprentissage qui se sont appliqués à rendre compte de ce phénomène, comme nous le verrons par la suite.
La mise en place d’un tel apprentissage nécessite généralement un grand nombre d’appariement SC/SI, de l’ordre de 400 (Tableau O-1) que le conditionnement soit effectué en protocole de Délai ou de Trace. De plus, les intervalles de Trace que peut supporter ce conditionnement sont courts, de l’ordre de quelques centaines de millisecondes (Tableau O-1).
Références Protocole Durée du
SC en ms Durée de l'intervalle de Trace en ms Durée du SI en ms nombre total d'appariements Trace courte 100 300 150 Trace longue 100 500 150
Nowak et Gormezano (1990) Délai 400 0 100 480
Délai 350 0 100
Trace 250 500 100
Thompson et coll. (1996) ;
Griffin et coll. (2004) Trace 100 500 100 400
Chachich et Powell (1998) Délai 1000 0 250 450 maximum
Délai 400 0 150 Trace 100 500 150 Délai 250 0 100 Trace 250 250 100 Délai 650 0 50 Trace 500 500 50 Délai 250 0 100 Trace courte 250 250 100 Trace longue 250 500 100 Délai 1000 0 100 Trace 500 500 100 Délai 590 0 10 Trace 280 300 10 Kim et coll. (1995) 90 McLaughlin et coll. (2002) 960 Tseng et coll. (2004) 300 Green et Arenos (2007) 540 Oswald et coll. (2006) 450 Kronforst-Collins et Disterhoft (1998) ; McEchron et Disterhoft (1999) 800 Weiss et coll. (1999) 750
Moyer et coll. (1990) entre 400 et 800
Tableau O-1 : Paramètres courants du conditionnement de la membrane nictitante. Dans le protocole de Délai, SC et SI se terminent au même instant (coterminent). Ces intervalles de
b) Conditionnement de peur.
L’intégralité de nos travaux se situe dans le cadre du conditionnement de peur. Dans le conditionnement de peur, le SC va la plupart du temps être un son et le SI un choc électrique appliqué au niveau des pattes d’un animal explorant un environnement. En raison de l’appariement systématique entre ces deux stimuli, une association va se mettre en place. Cette association se traduira par la suite par l’apparition d’une RC de peur lors des présentations ultérieures du son. La peur conditionnée se manifeste par diverses réactions comportementales et viscérales (Fanselow, 1984; LeDoux, 1998; Antoniadis et McDonald, 1999), mais la mesure la plus utilisée chez le rongeur est le comportement d’immobilité ou
freezing. Cette RC particulière se caractérise par une immobilisation totale de l’animal à
l’exception des mouvements respiratoires (Blanchard et Blanchard, 1969).
A l’inverse du conditionnement de la membrane nictitante, la spécificité temporelle de la RC de peur suite à un conditionnement de Trace est peu documentée. Il semblerait tout de même qu’il existe une sélectivité de la RC pour des intervalles de Trace inférieurs à 12s (Burman et Gewirtz, 2004). Cependant cette sélectivité serait moindre lors d’utilisation d’intervalles plus longs (Marchand et Kamper, 2000).
Cette forme d’apprentissage présente de multiples intérêts dans le cadre de l’étude du conditionnement de Trace. Premièrement, ce type de conditionnement nous permettra d’évaluer la relation entre conditionnement de Trace et conditionnement contextuel, comme nous le verrons plus loin. Deuxièmement, le conditionnement à un stimulus élémentaire reste possible avec des intervalles de Trace allant jusqu’à une minute (Kamin, 1965; McKinzie et Spear, 1995). En particulier, des intervalles de Trace de 20 à 30 s sont couramment employés (Tableau O-2), ce qui semble approprié dans le cadre de l’étude d’un phénomène concernant des stimuli séparés dans le temps. Enfin ce conditionnement peut être obtenu en peu d’essais, fréquemment moins d’une dizaine, qui peuvent être effectués au cours d’une même séance d’acquisition.
c) Aversion gustative conditionnée.
Ce type de conditionnement consiste à associer un aliment ou une boisson peu familière (SC) avec un malaise gastrique induit par l’expérimentateur (SI). L’association entre ces deux stimuli se traduira par la suite par une réponse d’évitement de l’aliment conditionné (RC) (Garcia et coll., 1966). Cette association reste possible même si l’intervalle entre le SC
et le SI est de plusieurs heures. La possibilité d’obtenir un conditionnement malgré des intervalles de Trace aussi longs semble inexplicable par toutes les formulations théoriques basées sur la contiguïté (Rozin et Kalat, 1971), même si un intervalle de Trace de quelques centaines de millisecondes est déjà en lui-même une violation de la contiguïté (Gormezano et Kehoe, 1981).
Références Protocole Durée du SC en s Durée de l'intervalle de Trace en s Durée du SI en s nombre total d'appariements Délai 15 0 0,5 10 Trace 15 30 0,5 10 Délai 15 0 0,5 10 Trace 15 30 0,5 10 Délai 5 0 0,5 Trace 5 2 0,5 Délai 30 0 1 2 Trace 30 2,5 1 4 Délai 16 0 2 Trace 16 18 2 Délai 10 0 0,7 Trace 10 20 0,7 Délai 15 0 0,5 8 Trace 15 30 0,5 8 Délai 20 0 2 5 Trace 20 20 (maximum) 2 5 Gilmartin et McEchron (2005) Trace 5 20 0,8 24 Délai 30 0 2 1 Trace 30 45 (maximum) 2 1 Délai 20 0 1 5 Trace 20 60 (maximum) 1 5
Blum et coll. (2006) Trace 10 20 0,7 7
Long Délai 20 0 1 10
Trace 10 10 1 10
Yoon et Otto (2007) Trace 20 30 2 10
Quin et coll. (2008) Trace 16 28 2 10
Wanisch et coll. (2005)
Hunt et Richardson (2007)
Runyan et coll. (2004)a ;
(2004)b 8 Weitemier et coll. (2004) Chowdhury et coll. (2005) ; Wiltgen et coll. (2005) Misane et coll. (2005) 10 Huerta et coll. (2000) Kinney et coll. (2002) Han et coll. (2003) 6 McEchron et coll. (1998) Baeg et coll. (2001)
3) Aspects cognitifs du conditionnement de Trace.
Si l’étude des mécanismes permettant la mise en place du conditionnement de Trace présente un intérêt en soi, les recherches actuelles menées sur les mécanismes et les spécificités de ce conditionnement suggèrent qu’il pourrait constituer un modèle d’étude d’un grand intérêt dans le cadre de la compréhension de processus neurobiologiques majeurs tels que ceux impliqués dans l’attention, la conscience ou encore le vieillissement cérébral.
a) Trace, attention, conscience.
Une des particularités du conditionnement de Trace par rapport aux conditionnements effectués en protocole de Délai est qu’il semble nécessiter une capacité cognitive d’ordre supérieur, la conscience. Il a effectivement été montré chez l’homme, à la suite d’un conditionnement du réflexe de clignement palpébral, que le niveau de RC manifesté en présence du SC était étroitement lié au degré de conscience qu’auraient les sujets du lien d’appariement entre SC et SI. La conscience de cette relation n’influe sur les performances que dans le cas d’un conditionnement de Trace et mais pas en protocole de Délai (Clark et Squire, 1998). Ces résultats ont par la suite été reproduits à plusieurs reprises (Clark et Squire, 1999; Manns et coll., 2000b, 2000a, 2002).
Dans une étude de 1999, Robert Clark et Larry Squire ont mis en évidence que l’insertion d’un stimulus distracteur ponctuel, présenté de façon régulière durant tout le conditionnement, induisait chez l’homme une diminution de la conscience de la relation entre SC et SI, que le conditionnement soit effectué en Délai ou en Trace. Cependant la présence du distracteur n’induisait un déficit de RC que chez des sujets conditionnés en Trace. De plus, ce déficit était similaire à celui observé chez les sujets conditionnés en Trace sans stimulus distracteur, quand ces sujets n’avaient pas pris conscience de la relation entre SC et SI (Clark et Squire, 1999). Il a été montré par la suite chez l’homme, dans le cadre d’un conditionnement de peur, que le conditionnement de Trace, mais pas de Délai, représente une charge attentionnelle (Carter et coll., 2003).
Les relations entre conscience et attention identifiées ainsi dans le conditionnement de Trace sont d’une importance capitale en ce qui concerne la généralisation de ces concepts à l’animal (Griffin et Speck, 2004). Il s’agit d’un point théorique difficile, mais le fait que, chez la souris, la présentation d’un stimulus distracteur durant un conditionnement de peur interfère
là aussi sélectivement avec le conditionnement de Trace (Han et coll., 2003) suggère qu’un processus analogue à la conscience pourrait également être requis pour l’établissement d’un conditionnement de Trace chez le rongeur.
Ces résultats pourraient, selon les auteurs, rendre compte de l’implication dans le conditionnement de Trace de structures cérébrales telles que l’hippocampe ou encore le cortex cingulaire antérieur, en raison de leur rôle supposé dans les processus de mémoire déclarative et attentionnels (Clark et Squire, 1998; Han et coll., 2003).
b) Vieillissement et Pathologies.
Le vieillissement cérébral s’accompagne un certain nombre de déficits mnésiques, spécialement en ce qui concerne les tâches impliquant le lobe temporal médian (Rapp et Amaral, 1991; Knuttinen et coll., 2001; Rosenzweig et Barnes, 2003). Une partie de ces déficits semblent imputable à des troubles de l’attention chez les sujets âgés (Burke et coll., 1987; Amenedo et Diaz, 1998; Kok, 2000). Par conséquent, il a été suggéré que l’âge pouvait aussi induire des déficits de conditionnement de Trace. Cela semble effectivement être le cas puisqu’il a été montré que des sujets âgés présentent un déficit sélectif dans leur capacité à acquérir un conditionnement de Trace du réflexe de clignement palpébral (Weiss et Thompson, 1992; Knuttinen et coll., 2001) alors qu’ils arrivent tout de même à acquérir ce conditionnement en protocole de Délai, bien que la vitesse d’acquisition soit plus lente que celle de sujets plus jeunes (Woodruff-Pak et Thompson, 1988; Weiss et Thompson, 1992; Solomon et Groccia-Ellison, 1996; Knuttinen et coll., 2001). Plus récemment, des résultats comparables ont été obtenus avec un conditionnement de peur effectué en Trace chez le rat (McEchron et coll., 2004; Moyer et Brown, 2006). Le fait que les animaux âgés ne soient déficitaires que dans le cas du conditionnement de Trace confirme que ce déficit est bien d’ordre mnésique et non pas simplement lié à l’expression de la RC.
L’étude des mécanismes sous-tendant le conditionnement de Trace semble donc très intéressante dans le cadre de la compréhension des perturbations neurobiologiques induites par l’âge et dans la recherche de traitements visant à les compenser.
III- Aspects théoriques du conditionnement de Trace.
Le conditionnement de Trace doit son nom à la principale théorie visant à rendre compte de la possibilité des animaux à établir une association Pavlovienne malgré la présence d’un intervalle de temps. Il a en effet très rapidement été suggéré que si le SC n’était pas présent au moment de l’arrivé du SI, il devait néanmoins avoir laissé une empreinte ou "Trace" de son passage. Ce serait alors cette Trace du SC et non le SC lui-même qui serait associé au SI.
Il est intéressant de noter que cette hypothèse peut être mise en parallèle avec la théorie du philosophe anglais David Hartley (1705-1757) pourtant antérieur à Pavlov. Sa théorie des vibrations, bien qu’incorrecte au niveau physiologique, postulait effectivement déjà à l’époque l’existence « d’une vibration physiologique rendant compte d’une persistance des sensations après arrêt de l’événement stimulant » (Gormezano et Kehoe, 1981).
Ainsi le problème majeur posé par le conditionnement de Trace consisterait à comprendre quelle pourrait être la nature de la " Trace " laissée par le SC. Il existe à l’heure actuelle deux théories majeures: la médiation contextuelle et la Trace persistante.
1) La Médiation contextuelle.
Lors de tout conditionnement, il existe un ensemble d’éléments présents en permanence dans l’environnement du sujet qui eux aussi sont capables d’entrer en association avec le SI. L’ensemble de ces éléments est regroupé sous le terme générique de "contexte". Les théories actuelles du conditionnement contextuel supposent que, au contraire d’une association avec un stimulus élémentaire, l’association entre le contexte et le SI nécessite l’acquisition préalable par l’animal d’une représentation unifiée/configurale de cet ensemble d’éléments (Rudy et O'Reilly, 1999).
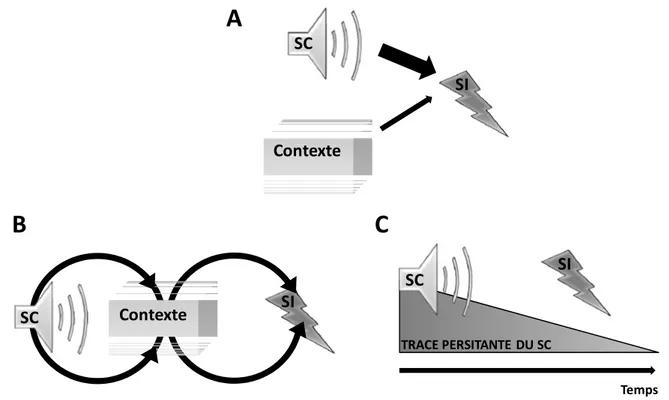
L’hypothèse de la médiation contextuelle repose sur cette capacité de l’animal à associer le contexte et le SI, car elle postule que c’est cette association qui permettrait à l’animal d’établir une association entre le SC présenté en Trace et le SI (figure O- 1B). L’hypothèse de la médiation contextuelle regroupe en fait trois hypothèses distinctes mais ayant toutes trois comme point commun de donner un rôle central au contexte de conditionnement lors du conditionnement de Trace.
Selon la première hypothèse, le conditionnement de Trace serait en fait la résultante d’une association de second ordre entre le SC et le contexte (Quinn et coll., 2002). Le conditionnement de second ordre est généralement mis en évidence à l’aide d’un protocole particulier durant lequel une première phase consiste à présenter un premier SC (SC1) apparié à un SI. Dans une seconde phase, on présente un deuxième SC (SC2) de manière appariée au SC1. Après cette seconde phase le SC2 pourra évoquer une RC comme s’il avait été directement associé au SI (Yin et coll., 1994). Dans le cas du conditionnement de Trace le contexte jouerait le rôle de SC1 et le stimulus présenté en Trace celui de SC2. En outre les deux phases d’association seraient effectuées simultanément (Kaplan et Hearst, 1982; Chang et coll., 2004).
La deuxième hypothèse repose sur la nature même du conditionnement contextuel et postule que le conditionnement de Trace résulterait d’une intégration du SC à la représentation configurale du contexte. Le SC serait en fait considéré comme un élément à part entière du contexte et associé au SI en tant que tel (Quinn et coll., 2002; Green et Arenos, 2007).
La troisième hypothèse reste relativement vague et postule que c’est en fait l’intégralité de l’épisode de conditionnement qui serait rappelée par la présentation du SC lors du test.
Le point commun entre ces trois hypothèses est qu’elles nécessitent toutes les trois que l’animal soit capable d’établir un conditionnement au contexte pour pouvoir établir un conditionnement de Trace.
Ainsi, il apparaît nécessaire de tenter d’élucider les relations entre le conditionnement de Trace et le conditionnement contextuel en tentant pour cela de mettre en évidence d’éventuelles dissociations entre ces deux types de conditionnement (Marchand, 2004).
2) La Trace persistante.
Une seconde classe d’hypothèses suppose que le SC serait capable à lui seul d’induire une activité persistante qui constituerait le stimulus effectif entrant en association avec le SI. La Trace du SC serait alors une sorte de "représentation" du SC au niveau neuronal, et c’est cette "représentation" qui serait associée au SI (Figure O-1B). L’hypothèse de la trace persistante rapprocherait ainsi le conditionnement de Trace du conditionnement de Délai en
rétablissant au niveau neuronal la contiguïté absente entre les stimuli. Une telle hypothèse ne nécessite pas l’intervention d’indices autres que le SC lui-même.
La Trace du SC est définie comme le maintien, déclenché par la présentation du SC, d’une information qui est propre à celui-ci. La question reste de savoir par quel processus ce maintien de la Trace neuronale durant plusieurs dizaines de secondes serait rendu possible. Il est possible d’envisager qu’un des processus requis serait la mémoire de travail.
La mémoire de travail peut être définie comme étant un système ayant une capacité de stockage limitée et temporaire, et une capacité de manipuler les informations lors de tâches cognitives (Hasselmo et Stern, 2006). Ce type de mémoire est actif et efficace uniquement durant une courte période de temps, généralement de l’ordre de plusieurs dizaines de secondes (Goldman-Rakic, 1995). Un exemple couramment utilisé d’utilisation humaine de ce type de mémoire est la capacité de retenir un nouveau numéro de téléphone qui, une fois composé, aura de forte chance d’être oublié. Chez l’animal, ce type de mémoire est généralement évalué dans des paradigmes de reconnaissance différée (appariement ou non appariement retardé) dans lesquels l’animal doit retenir durant quelques secondes la nature d’un item (objet, odeur, emplacement) qui vient de lui être présenté, afin d’effectuer un choix au terme d’un délai imposé.
Compte tenu de leurs définitions respectives, il semble possible que ce soit par le biais de la mémoire de travail que la Trace du SC serait maintenue durant l’intervalle de Trace.
Pourtant, à notre connaissance, aucun auteur n’a encore démontré de façon probante cette possibilité, même si elle a déjà été suggérée (Kesner, 2005). Ceci est probablement en grande partie dû au fait que la mémoire de Travail est mise en jeu dans la réalisation de tâches préalablement entraînées et que c’est la connaissance préalable de cette tâche qui induit le maintien de l’information pertinente. Dans le conditionnement de Trace, à l’inverse, l’information à maintenir est relativement nouvelle et ne devient pertinente qu’au cours de l’association SC/SI.
Cependant, nous pouvons poser l’hypothèse que la mémoire de Travail et le conditionnement de Trace sont étroitement liés. S’il existe une telle relation entre ces deux processus, alors il devrait être possible d’observer un certain nombre de similitudes entre les systèmes neurobiologiques qui les sous-tendent. Nous verrons que le travail présenté dans ce mémoire met en évidence certaines de ces similitudes.
SC SI Contexte SC Contexte SI SC SI TRACE PERSITANTE DU SC Temps
A
B
C
3) Modèles Formels.Il existe plusieurs modèles formels d’apprentissage pouvant être utilisés pour expliquer le conditionnement de Trace. Nous ne ferons pas ici une liste exhaustive de ces modèles mais nous parlerons de trois modèles présentant tous un intérêt particulier vis-à-vis d’au moins une composante du conditionnement de Trace. Bien que ces modèles ne soient pas discutés par la suite, ils sont en partie responsables du choix de certains de nos paramètres expérimentaux ainsi que du choix des moments de réponse observée.
a) Le modèle de Rescorla-Wagner.
Le modèle de Rescorla et Wagner (1972) est un modèle formel d’apprentissage basé sur les lois de contiguïté et de contingence que nous avons explicitées précédemment, mais
Figure O-1: Représentation schématique du mode d’association du SC et du SI. A : Lors d’un conditionnement de délai, le SC et le SI sont contigus. L’association s’effectue directement, il peut aussi y avoir une faible association du contexte avec le SI. B,C : lors d’un conditionnement de Trace. B : Selon l’hypothèse de la médiation contextuelle, le contexte va servir de support au SI et le niveau de conditionnement du SC sera alors dépendant de l’association préalable du contexte et du SI. C : Selon l’hypothèse de la Trace persistante, la présentation du SC va déclencher une activité neurobiologique persistant dans le temps et c’est cette Trace persistante qui sera le support effectif de l’association avec le SI.
plusieurs SC selon leur histoire. Il permet ainsi d’expliquer les phénomènes de compétition tels que le masquage ou le blocking, mais aussi des phénomènes tels que l’extinction et l’inhibition conditionnée. Cette théorie suppose que l’expression de la RC est directement fonction d’une "force associative" acquise par un SC lors de son appariement avec un SI. L’incrément de force associative lors d’un essai obéit à l’équation suivante :
ΔVX n+1 = αXβ (λ –Vntotal)
Cette équation décrit le changement ΔVX n+1 de la force associative (V) d’un CS X
après l’appariement avec un SI lors de l’essai n+1; αX est le potentiel d’association du CS X
(ou saillance, liée en grande partie à son intensité) ; β est le potentiel d’association du SI (lié en grande partie à son intensité) ; λ est la force associative maximale que le SIpeut assumer dans une situation donnée ; Vntotal est la somme des forces associatives de l’ensemble des CS
présents lors de l’essai n+1.
Cette équation prédit qu’au fil des appariements, la force associative totale Vntotal tend
vers une asymptote λ déterminée par les caractéristiques du SI. Lorsque cette force associative totale est déjà importante, un SC donné ne pourra gagner que peu de force associative lors d’un essai. L’existence d’une force associative totale limitée qui doit se partager entre les divers SC en présence est à la base des processus de compétition (Miller et coll., 1995).
Ce modèle met en évidence plusieurs points essentiels que nous aurons à prendre en compte par la suite. Un SI donné ne peut pas s’associer à l’infini mais ne s’associera préférentiellement qu’avec les éléments les plus saillants et les plus prédictifs de son occurrence.
Cependant, dans le cadre de notre étude, ce modèle pose un problème car il est basé sur la notion d’essai et ne considère que les SC présentés de façon contiguë avec le SI. Dans sa forme de base, le modèle de Rescorla-Wagner ne tient pas compte de l’agencement temporel des stimuli et n’est donc pas directement applicable au conditionnement de Trace.
b) Le modèle SOP
Un autre type de modèle peut prendre en compte la configuration temporelle des stimuli, le SOP (Sometimes Opponent Process) de Wagner (Wagner, 1981; Wagner et Brandon, 1989). Ce modèle est conçu comme une variation "en temps réel" du modèle de
Rescorla-Wagner. Le principe central de ce modèle est qu’il stipule que la représentation cérébrale d’un stimulus consiste en un large ensemble d’éléments théoriques appelés nœuds. Chaque élément d’un nœud peut assumer trois états dynamiques distincts : les états actifs A1 et A2 et l’état inactif I. La présentation d’un stimulus va faire passer le nœud de l’état inactif I à l’état actif A1. Le nœud ne va pas rester dans cet état mais passer progressivement à l’état A2 et enfin si le stimulus n’est plus présent, passer progressivement à l’état I. Dans le cadre du conditionnement Pavlovien, il y a donc deux nœuds essentiels, le premier codant pour la représentation du SC et le second pour la représentation du SI. Le SOP suggère que 2 nœuds vont être associés lorsqu’ils sont dans un état actif identique. Ainsi pour qu’un conditionnement associatif ait lieu, il faut que le nœud codant pour le SC soit encore dans l’état A1 au moment de l’arrivée du SI. Lors de la phase de test, cette association se traduira par le fait que la présentation du SC et le passage du nœud SC en état A1 induira un passage du nœud SI de l’état I à l’état A2. C’est ce passage en état A2 du nœud SI qui induit l’expression de la RC.
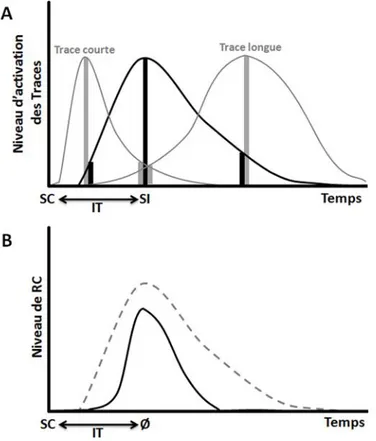
Le conditionnement de Trace serait donc explicable si l’on suppose que le passage de l’état A1 à l’état A2 se fait très lentement (plusieurs centaines de millisecondes pour le conditionnement de la membrane nictitante mais plusieurs dizaines de secondes pour le conditionnement de peur) (Figure O-2).
Cependant, si le SOP permet d’envisager la possibilité d’établir un conditionnement de Trace, il ne permet pas de comprendre la spécificité temporelle des réponses manifestées lors de certains conditionnements de Trace tel que le conditionnement de la membrane nictitante. En effet, la cinétique de passage des nœuds entre ces différents états est fixée dès le départ par le modèle. Elle contraint à la fois les intervalles interstimuli pour lesquels le conditionnement de Trace pourra être obtenu et les cinétique des réponses conditionnées. Ceci a conduit Wagner et Brandon (1989) à envisager l’existence de deux systèmes de type SOP en parallèle. Le premier avec une cinétique rapide rendrait compte des RC défensives ou d’évitement comme le clignement de la paupière. Le second beaucoup plus lent rendrait compte des RC affectives/émotionnelles comme dans le cas du conditionnement de peur.
SI A1 A2 I SC A1 A2 I SI SC A2 A1
A
B
IT P ropor ti on to ta le d ’él ém en ts SC P ropor ti on to ta le d ’él ém en ts SI P ropor ti on to ta le d ’élém en ts SC et SI A1 A2 A1SC A1SI c) Le modèle spectral.Un certain nombre de modèles, dits modèles spectraux, visent à mieux rendre compte de la spécificité temporelle de la réponse conditionnée. Le modèle spectral (Church, 1999) postule que tout événement sensoriel va induire de nombreux patrons d’activité parallèles ou Traces au niveau du système nerveux central. Chaque Trace a une cinétique et une durée de vie qui lui est propre. Ainsi, le temps passé depuis la présentation du SC serait représenté par le patron des Traces encore actives à ce moment là et c’est donc l’ensemble de ces Traces qui serait associé au SI (Buonomano et Mauk, 1994). Ce modèle peut supposer de plus que les Traces qui sont actives sans présentation du SI deviennent inhibitrices (Marchand et coll., 2004). Ainsi, toute les Traces s’éteignant avant la présentation du SI et toutes celles persistant après la présentation de ce dernier deviendraient inhibitrices comme l’indique la figure O-3.
Figure O- 2 : Modèle SOP. A : Représentation de deux nœuds dans le modèle d’apprentissage SOP, l’un étant activé par le SC et l’autre par le SI lors d’un conditionnement Pavlovien. Les cercles interconnectés représentent les trois états possibles de chaque élément des nœuds. Les flèches indiquent le sens de passage d’un état à un autre. Les nœuds sont reliés entre eux pour indiquer la possibilité d’une association excitatrice (flèche) ou d’une association inhibitrice (ligne en T). B : Représentation d’une simulation décrivant la proportion d’éléments du nœud SC (graphe du haut) et d’éléments du nœud SI (graphe du milieu) en état A1 ou A2 à un instant donné. Le graphe du bas met en évidence le fait qu’un certain nombre d’éléments des nœuds SC et SI peuvent se retrouver dans l’état A1 simultanément, même en présence d’un intervalle de Trace. Abréviation : A1 et 2, état Actif ; I, état Inactif ; IT, intervalle de Trace.
La cinétique de la RC évoquée refléterait alors une combinaison du profil des Traces qui ont été associées au SI. Le maximum de RC évoqué correspondrait alors approximativement au moment de survenue du SI. Ce modèle, qui existe sous plusieurs formes (Desmond et Moore, 1991; Grossberg et Merrill, 1992; Young et coll., 1997; Buhusi et Schmajuk, 1999) présente donc l’intérêt d’expliquer la sélectivité temporelle des RC observées dans le cadre de certains conditionnements de Trace. Il faut cependant préciser que cette spécificité temporelle est surtout observable pour des intervalles de Trace courts dans un conditionnement de la membrane nictitante (Grossberg et Schmajuk, 1989). Il n’est donc pas certain que ce modèle reste valable pour les intervalles de temps classiquement utilisés en conditionnement de peur (Buhusi et Schmajuk, 1999; Marchand et Kamper, 2000; Marchand et coll., 2004).
Figure O- 3 : Modèle Spectral. A : Niveau d’activation de plusieurs Traces déclenchées par le même SC en fonction du temps. Seule la Trace ayant un niveau maximum au moment de l’apparition du SI (courbe noire) va être associée au SI de façon activatrice, les autres Traces (en gris) vont être associées au SI de façon inhibitrice. Les barres noires représentent le niveau d’activation et les barres grises le niveau d’inhibition à un instant t. B : Cinétique du niveau de réponse conditionnée (RC) lors du Test (courbe noire pleine). La courbe grise représente la cinétique hypothétique de réponse sans les associations inhibitrices. On peut voir que c’est le cumul des associations activatrices et inhibitrices qui permet la mise en place d’une RC centrée autour du moment prédit de l’arrivée du SI (Ø). Abréviations : IT, intervalle de Trace ; SI, stimulus inconditionnel ; SC, stimulus conditionnel.
IV- Aspects neurobiologiques.
Nous présenterons ici principalement les circuits neurobiologiques sous-tendant les conditionnements de la membrane nictitante et de peur lorsqu’ils sont effectués en protocole de Délai, car la connaissance préalable de ces circuits est indispensable dans le cadre de l’étude du conditionnement de Trace. Nous mentionnerons ici brièvement les principales structures spécifiquement impliquées dans les versions "Trace" de ces paradigmes puisque les mécanismes et les fonctions de ces structures seront par la suite plus précisément décrits dans le corps de la thèse.
1) Conditionnement de la membrane nictitante en Délai.
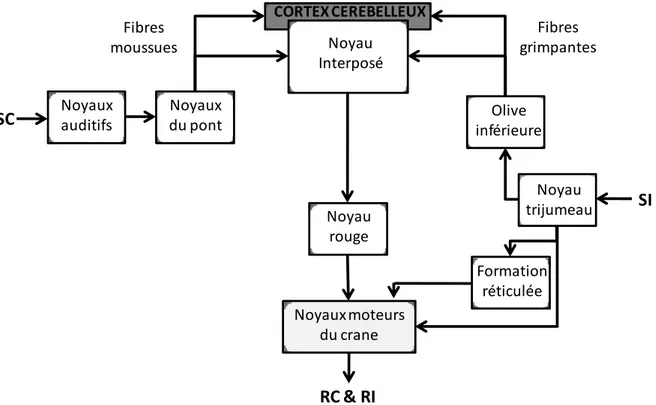
Il est de nos jours admis que, lors d’un conditionnement de la membrane nictitante effectué en Délai, le cervelet et les circuits qui lui sont associés sont essentiels (figure O-4) (Thompson et Krupa, 1994; Woodruff-Pak et Disterhoft, 2008).
Ce circuit de base comprend deux voies parallèles responsables respectivement du traitement du SC et du traitement du SI et concerne essentiellement des structures du tronc cérébral et cérébelleuses.
Le traitement d’un SC de nature auditive serait effectué par un circuit partant des noyaux auditifs et relayé au noyau interposé et au cortex cérébelleux via les fibres moussues en provenance des noyaux du pont. Il a effectivement été montré que des lésions de ce noyau induisaient un déficit de conditionnement à un stimulus auditif mais pas lumineux (Steinmetz et coll., 1987). De plus, des stimulations électriques de cette régions peuvent faire office de SC et se révèlent même plus efficaces qu’un stimulus sonore (Steinmetz et coll., 1986; Tracy et coll., 1998). En outre, une lésion du noyau interposé empêche le conditionnement (Steinmetz et coll., 1986).
La voie du traitement du SI serait en fait l’association de deux voies, l’une responsable de l’émission de la réponse inconditionnée (RI) et l’autre responsable de l’association du SI avec le SC et de la mise en place de la RC. Il est ici important de rappeler que la RI et la RC sont de nature identique dans le cadre du conditionnement de la membrane nictitante.
Ces deux voies parallèles ont pour origine les noyaux trijumeaux qui reçoivent les informations sensorielles (tactiles et nociceptives). Lors de la mise en place de la RI, les
informations vont être rapidement transmises aux noyaux moteurs du crâne par une voie directe, mais aussi indirectement via la formation réticulée. Lors de l’association SC/SI, les informations vont être transmises au noyau interposé et au cortex cérébelleux via les fibres grimpantes en provenance de l’olive inférieure. Il a effectivement été montré qu’une lésion de l’olive inférieure effectuée juste avant l’acquisition empêche la mise en place du conditionnement (Mintz et coll., 1994). De plus, les neurones de cette aire ne répondent pas au SC mais uniquement à la présentation du souffle d’air (SI) (Sears et Steinmetz, 1991). L’olive inférieure est donc capitale dans le traitement du SI lors de l’apprentissage.
La quasi indépendance des voies de traitement du SI responsables de l’expression de la RI et de l’association avec le SC a pu être mise en évidence par le fait qu’un certain nombre de lésions du cervelet telle que celles des noyaux interposés abolissent totalement l’ensemble des composantes de la RC sans affecter la RI, indépendamment de l’intensité du SI (Steinmetz et coll., 1992; Ivkovich et coll., 1993). De plus, de façon encore plus surprenante, une inactivation réversible de la formation réticulée par injection de muscimol juste avant la phase d’acquisition abolit totalement la RI manifestée en présence du souffle d’air mais n’induit pas de déficit de performance lors de la phase de test ultérieure (Zhang et Lavond, 1991). Ainsi, l’association entre SC et SI peut être acquise alors même que la RI n’est pas exprimée.
Enfin il est supposé que c’est l’activation conjointe des voies traitant le SI et le SC et leur convergence au niveau du cortex cérébelleux et du noyau interposé qui permet l’association entre les deux stimuli (Christian et Thompson, 2003).
SI Noyaux
auditifs
Noyaux
du pont inférieureOlive
Noyau trijumeau Formation réticulée Noyau rouge Noyaux moteurs du crane Noyau Interposé Fibres grimpantes CORTEX CEREBELLEUX SC RC & RI Fibres moussues
2) Conditionnement de Peur en Délai.
L’apparente simplicité du conditionnement de peur est à mettre en parallèle avec la difficulté d’explication des processus opératoires sous-jacents. Pourtant, depuis quelques années, des apports expérimentaux majeurs ont conduits à l’émergence d’un consensus sur ces mécanismes. Il a ainsi été défini deux circuits du conditionnement de peur sous-tendant respectivement le conditionnement à un stimulus élémentaire et le conditionnement au contexte.
a) Conditionnement de peur à un stimulus élémentaire.
C’est principalement un circuit direct thalamo-amygdalien, ou "circuit rapide" en provenance du corps genouillé médian et du noyau intralaminaire postérieur (LeDoux et coll., 1990; Turner et Herkenham, 1991) qui permettrait la formation d’un conditionnement
Figure O- 4 : Schéma simplifié du circuit cérébelleux impliqué dans le conditionnement de la membrane nictitante en protocole de Délai. Deux voies parallèles sous-tendent le traitement du SC et le traitement du SI. Ces deux voies présentent des parties communes notamment au niveau du cortex cérébelleux et du noyau interposé.
élémentaire impliquant un SC sonore simple et unimodal. Les informations relatives au SI transiteraient également par ce circuit court (figure O-5).
Lorsque le SC doit faire l’objet d’une discrimination sur la base de ses propriétés perceptuelles, un second circuit, plus "long" incluant le cortex auditif semble s’avérer nécessaire (Romanski et LeDoux, 1993). Le circuit rapide permettrait la mise en place de la RC avant même que le SC ne soit reconnu, la reconnaissance de ce SC passant par le circuit long.
L’intégration des données de ces deux circuits semble se faire au niveau du noyau latéral de l’amygdale (LA). En effet, les projections thalamo-amygdaliennes et cortico-amygdaliennes convergent sur les même neurones au sein de ce noyau (LeDoux et coll., 1991; Li et coll., 1996). Des lésions ou des inactivations du LA provoquent d’importantes perturbations du conditionnement de peur à un SC élémentaire sonore, ce qui confirme le rôle de ce noyau dans ce type de conditionnement (Muller et coll., 1997; Wilensky et coll., 1999; Amorapanth et coll., 2000; Nader et coll., 2001)). Il est maintenant largement accepté que le LA serait un site clef de la plasticité synaptique nécessaire à l’association SC/SI (Maren, 2001; Ferreira et coll., 2008).
Enfin, c’est le noyau central de l’amygdale (CN) qui assurerait l’expression de la réponse conditionnée, qu’elle soit de nature viscérale, du fait de ses connexions aux régions hypothalamiques, pontine et médullaire impliquées dans la régulation du système nerveux autonome, ou comportementale par l’intermédiaire de la substance grise périaqueducale. Il a effectivement été montré qu’une atteinte du CN interfère avec l’expression des réponses conditionnées (Kapp et coll., 1979; Gentile et coll., 1986; Hitchcock et Davis, 1986; Nader et coll., 2001). Dans un modèle révisé, le CN reste central dans l’expression de la réponse de peur, mais aurait aussi un rôle prépondérant dans les phases d’acquisition et de consolidation de ce type de conditionnement (Pare et coll., 2004; Wilensky et coll., 2006), ce qui a conduit à une remise en cause du modèle classique (LeDoux, 2000) dans lequel le CN n’était pas un site de plasticité.
SC/SI THALAMUS SENSORIEL AMYGDALE LA BLA CN CORTEX AUDITIF RC
b) Conditionnement de peur au contexte.
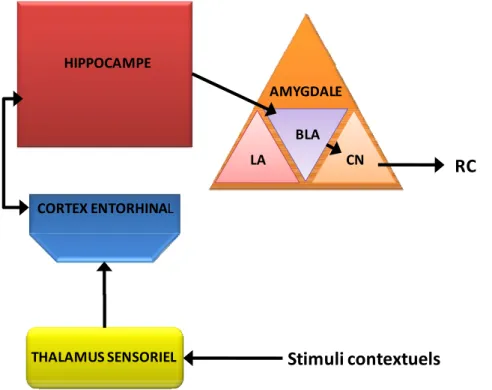
Que le choc soit présenté de manière appariée ou non appariée avec le SC ou encore en l’absence de SC, d’autres stimuli présents dans l’environnement peuvent être associés avec ce SI. Ces éléments, présents de manière continue durant le conditionnement (Rudy et O'Reilly, 1999; Fanselow, 2000) sont généralement regroupés sous le terme générique de "contexte". Si nous venons de voir que la structure centrale du conditionnement élémentaire était l’amygdale et plus précisément le complexe LA/CN, il n’en est pas de même en ce qui concerne le conditionnement contextuel (Selden et coll., 1991; Phillips et LeDoux, 1992).
En effet, de nombreuses études du conditionnement contextuel ont mis en évidence un rôle prépondérant de l’hippocampe et ont abouti à l’idée générale que l’intégrité de cette structure était nécessaire lors de ce conditionnement (Holland et Bouton, 1999; Rudy et coll., 2004). Les hypothèses actuelles supposent que cette structure serait nécessaire à la formation d’une représentation unifiée du contexte (Sutherland et Rudy, 1989; Rudy et coll., 2004). Cette hypothèse est particulièrement étayée par les données observées dans les études de déficit de choc immédiat. Le protocole de déficit de choc immédiat consiste à soumettre l’animal à un seul choc électrique immédiatement après l’avoir introduit dans le contexte de conditionnement et de le retirer immédiatement après. Lors de la phase de test, on s’aperçoit
Figure O-5: Circuit responsable de la peur conditionnée à un stimulus élémentaire. Le Thalamus sensoriel (circuit court) transmet à l’amygdale à la fois les informations concernant le SC et le SI. Les traitements plus complexes du SC sont effectués par le circuit long qui inclut le cortex auditif. Abréviations : BLA, noyau basolatéral ; CN, noyau central ; LA, noyau latéral ; RC, réponse conditionnée ; SC, stimulus conditionnel ; SI, stimulus inconditionné. Adapté de Wilensky et coll. 2006.
généralement que l’animal ne manifeste que très peu de peur associée au contexte. En revanche si l’animal a pu préalablement explorer le contexte pendant quelques minutes on s’aperçoit qu’à la suite du même conditionnement il manifeste beaucoup plus de RC au contexte (Kiernan et Westbrook, 1993; Wiltgen et coll., 2001). Il est supposé que la pré-exposition permet à l’animal d’établir une représentation configurale du contexte et que cette représentation est nécessaire à la mise en place de l’association (Rudy et O'Reilly, 1999, 2001; Rudy et coll., 2002). Il a en parallèle été montré qu’une inactivation de l’hippocampe durant la phase de pré-exposition perturbait par la suite l’établissement du conditionnement, confirmant ainsi le rôle de l’hippocampe dans la mise en place de la représentation du contexte (Matus-Amat et coll., 2004; Stote et Fanselow, 2004).
Les projections de l’hippocampe vers le complexe amygdalien et spécialement le BLA semblent capitale dans la mise en place du conditionnement de peur associée au contexte (LeDoux, 2000). Il a de plus récemment été montré que des inactivations du BLA préalablement au conditionnement induisait un déficit spécifique de la réponse de peur associée au contexte, confirmant l’implication sélective de ce noyau lors de ce type d’apprentissage (Nader et coll., 2001) (figure O-6). A l’inverse, l’inactivation du LA conduisait à une augmentation de la réponse au contexte en parallèle d’une diminution de la réponse au SC lors d’un conditionnement à un son (Calandreau et coll., 2005).
CORTEX ENTORHINAL HIPPOCAMPE THALAMUS SENSORIEL AMYGDALE LA BLA CN RC Stimuli contextuels
3) Mécanismes spécifiques des conditionnements de Trace.
Lors de l’utilisation d’un protocole de Trace, les circuits simples de conditionnement élémentaires ne seraient plus suffisants. Ainsi, aux voies "tronc cérébral-cervelet" pour le conditionnement de la membrane nictitante et "thalamo-amygdalienne" pour le conditionnement de peur s’ajouterait une étape de traitement du SC supplémentaire réclamant la mise en jeu d’une structure majeure dans les processus mnésiques : l’hippocampe.
En effet, il a été montré que des lésions de cette structure perturbaient fortement la mise en place d’un conditionnement de Trace sans perturber le conditionnement de Délai, aussi bien dans le cas du conditionnement de la membrane nictitante (Solomon et coll., 1986; Moyer et coll., 1990) que dans celui du conditionnement de peur (McEchron et coll., 1998; Burman et coll., 2006; Rogers et coll., 2006).
Il semblerait cependant que l’hippocampe soit impliqué de façon différente dans les deux types de tâches puisque dans le cadre du conditionnement de la membrane nictitante les substrats neurobiologiques sous-tendant l’acquisition des conditionnements de Trace et de Délai semblent pouvoir faire l’objet d’une double dissociation. L’hippocampe contribuerait spécifiquement au conditionnement de Trace, alors que le cortex cérébelleux ne serait indispensable que pour un conditionnement de Délai (Woodruff-Pak et coll., 1985; Gerwig et coll., 2008; Woodruff-Pak et Disterhoft, 2008). Ces auteurs émettent l’hypothèse que, dans le cadre d’un conditionnement de Trace mais non de Délai, le traitement effectué par l’hippocampe pourrait se substituer en partie à celui du cervelet pour fournir l’information concernant le SC aux noyaux du pont. Ce serait donc uniquement la voie du traitement du SC qui serait modifiée, alors que les mécanismes sous-tendant l’association et le traitement du SI resteraient les mêmes.
A l’inverse, dans le cadre du conditionnement de peur, il n’a jamais été possible, à notre connaissance, d’obtenir une telle dissociation entre Trace et Délai puisque toutes les manipulations affectant le conditionnement de Délai abolissent également le conditionnement de Trace. Ceci pourrait être dû en grande partie à la simplicité du circuit nécessaire au conditionnement de Délai et laisse supposer que l’intégralité du circuit impliqué dans ce type de conditionnement serait aussi nécessaire au conditionnement de Trace.