Influence des interactions avec le Soi sur le devenir des cellules T CD4 naïves en phase effectrice
Texte intégral
Figure





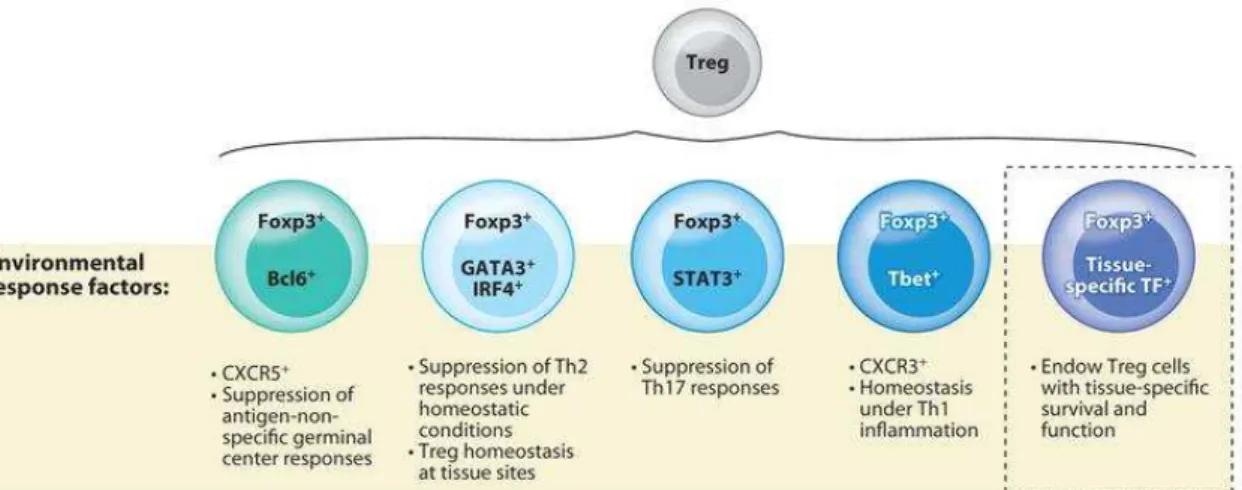
Outline
Documents relatifs
To test the hypothesis that CCR5 expression could determine the natural course of the disease in HIV-1–infected children, we com- pared their DR 2 CD4 + T cell surface CCR5 density
Pathogenic interplays between Hepatitis C virus, myeloid dendritic cell functions and virus-specific CD8+ T cell responses in chronically infected patients..
Mon travail de thèse a consisté à essayer de mieux comprendre les mécanismes d’action employés par ces lymphocytes T CD4 + CD25 + régulateurs pour contrôler la réponse
agonist complexes partially restored the homeostatic proliferation of regulatory CD4 + T cells injected into lymphopenic mice lacking expression of MHC class II molecules, whereas
agonist complexes partially restored the homeostatic proliferation of regulatory CD4 + T cells injected into lymphopenic mice lacking expression of MHC class II molecules, whereas
(B) The percentages of Foxp3 + T cells was assessed following 5 days of activation in nutrient-replete and glutamine-deprived conditions, in the presence or absence of
In conclusion, we showed that cDC1 and their cell- intrinsic response to STAT1 signalling are critical for adequate activation of the effector functions of CD8 + and CD4 + T cells
Our aim was to better characterize the effect of OCLs on T cells and in particular to com- pare both OCL subsets (DC-OCLs and MN-OCLs) in terms of antigen presentation and T
