Évaluation de l'impact du remplacement de la fléole des
prés par la fétuque élevée, en espèce pure ou en
mélange avec la luzerne, dans les rations des vaches en
lactation
Mémoire
Anne-Marie Richard
Maîtrise en sciences animales - avec mémoire
Maître ès sciences (M. Sc.)
Résumé
L’objectif du projet présenté dans ce mémoire de maitrise est d’évaluer l’effet du remplacement de la fléole des prés par la fétuque élevée, offertes en espèce pure ou en mélange avec la luzerne, et de vérifier l’impact de la méthode de conservation de la fétuque élevée, soit en ensilage préfané (35 % de matière sèche, MS) ou demi-sec (55 % de MS), sur les performances des vaches en lactation. Pour ce faire, cinq traitements ont été testés : 1) fléole des prés (70 % de la ration) en ensilage préfané ; 2) fléole des prés (42 % de la ration) + luzerne (28 % de la ration) en ensilages préfanés ; 3) fétuque élevée (70 % de la ration) en ensilage préfané ; 4) fétuque élevée (42 % de la ration) + luzerne (28 % de la ration) en ensilages préfanés ; 5) fétuque élevée (70 % de la ration) en ensilage demi-sec. Quinze vaches Holstein (128 33 JEL) ont été distribuées dans un triple carré latin 5 × 5. Afin de comparer les traitements, les contrastes a priori suivants ont été réalisés : 1) fléole des prés-ensilage préfané vs fétuque élevée-ensilage préfané [(FP et FP + L) vs (FE et FE + L)]; 2) espèces pures-ensilage préfané vs mélanges-ensilage préfané [(FP et FE) vs (FP + L et FE + L)]; 3) interaction entre l’espèce de graminée fourragère et le mélange, pour les traitements à base d’ensilage préfané; et 4) ensilage préfané de fétuque élevée vs ensilage demi-sec de fétuque élevée [FE vs. FE (DS)]. Les contrastes ont été considérés comme significativement différents à des valeurs de P 0,05 et une tendance a été considérée quand 0,05 P 0,10. Les résultats ont démontré que l’espèce de graminée n’a pas influencé la consommation volontaire de matière sèche, la production laitière, et la teneur en gras du lait. La prise alimentaire et la production laitière ont été supérieures avec les mélanges fourragers, mais la teneur en gras du lait n’a pas été avantagée par ces derniers. La teneur en protéine du lait n’a pas été affectée par l’espèce de graminée pour les traitements à base de mélange fourrager, mais a été supérieure avec la fléole des prés pour les traitements de graminées seules. Les vaches ont consommé davantage de fétuque élevée sous forme d’ensilage préfané comparativement à l’ensilage demi-sec, mais la production laitière et la teneur en gras du lait n’ont pas été affectées par le mode de conservation. En somme, les résultats démontrent que la fétuque élevée est une bonne alternative à la fléole des prés pour les vaches en lactation et confirment les effets bénéfiques des mélanges fourragers dans leur ration.
Table des Matières
Résumé
II
Table des Matières
III
Liste des tableaux
V
Liste des figures
VI
Liste des abréviations
VII
Remerciements
IX
Avant-propos
XII
Introduction
1
1.
Influence des fourrages sur la prise alimentaire des vaches
4
1.1. Impact de l’espèce fourragère 5
1.1.1. Appétence du fourrage 5
1.1.2. Quantité de fibres 5
1.1.3. Teneur en protéines 7
1.1.4. Les glucides non-structuraux 8
1.1.5. Influence des caractéristiques des fourrages et des nutriments contenus dans la diète
sur les signaux de satiété 10
1.1.6. Impact de mélanger des graminées avec des légumineuses sur la prise alimentaire 16 1.2. Impact du mode de conservation sur la prise alimentaire 18
1.2.1. Longueur de la fibre 18
1.2.2. Propriétés physico-chimiques 19
2. Influence des fourrages sur les paramètres ruminaux
22
2.1. pH ruminal 22
2.2. Azote 23
2.3. Acides gras volatils 24
3. Influence des fourrages sur la production laitière et ses
composants
25
3.1. Quantité de lait produit 25
3.2. Gras du lait 26
3.2.1. Acides gras du lait 28
3.3. Protéine 30
3.4. Urée 31
5. Performance of lactating dairy cows fed tall fescue or timothy as
sole forage of the diet or in association with alfalfa
35
5.1. Interpretative summary 36
5.2. Résumé 36
5.3. Abstract 37
5.4. Introduction 38
5.5. Materials and methods 39
5.5.1. Forage Production 39
5.5.2 Cows, Experimental Design and Diet 39
5.5.3. Measurements and Sampling 40
5.5.4. Chemical Analysis 41
5.5.5. Statistical Analysis 42
5.6. Results and discussion 44
5.6.1. Intake 44
5.6.2. Rumen Fermentation 45
5.6.3. Milk yield, and milk composition 46
5.7. Conclusion 48
5.8. Acknowledgments 48
5.9. References 50
Conclusion générale
66
Liste des tableaux
Tableau 1.1. Concentrations en N, aNDF, ADF, IVTD, GNS et sucres solubles de la fléole
des prés et de la fétuque élevée, en première coupe fauchées le matin……….9
Tableau 1.2. Contenu en protéines brutes (PB), en azote non protéique (ANP) et en protéine vraie de différents fourrages selon leur entreposage………...21
Tableau 1.3. Analyse des fourrages au 31 décembre 2015 de Valacta………...22
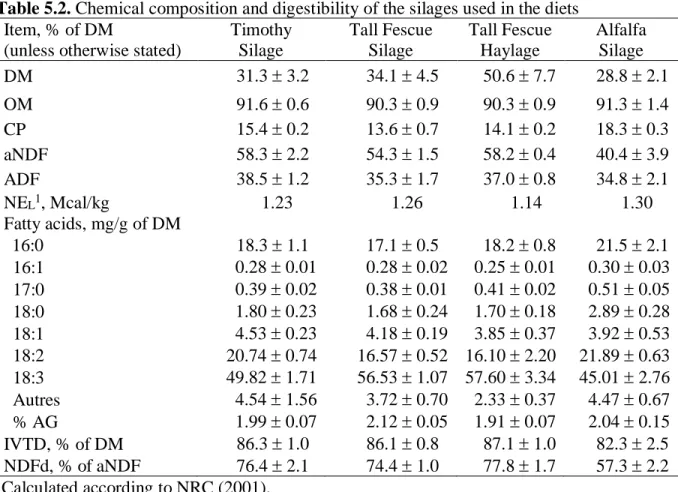
Tableau 5.1. Ingredients of the experimental total mixed rations………...56
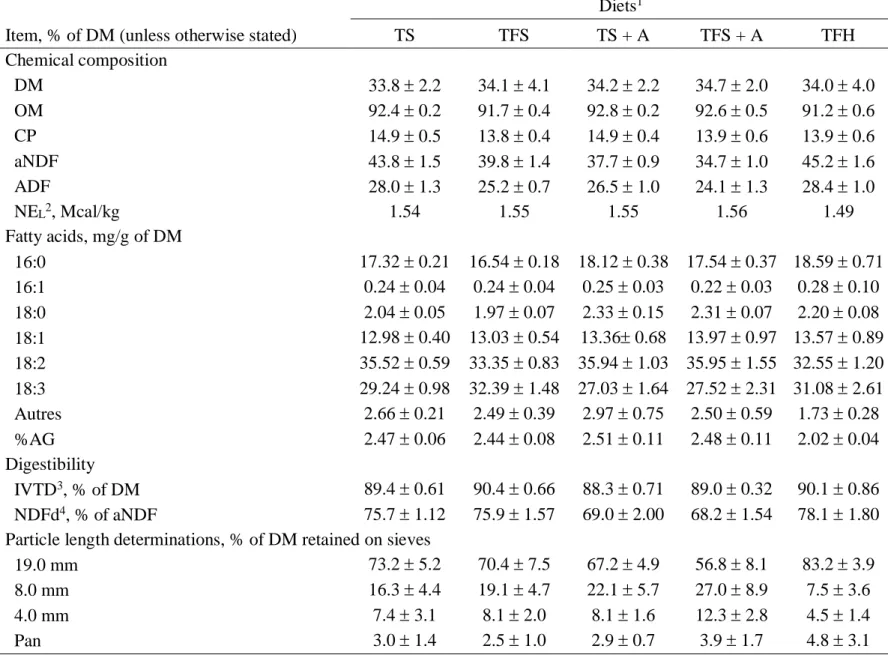
Tableau 5.2. Chemical composition of the forages……….57
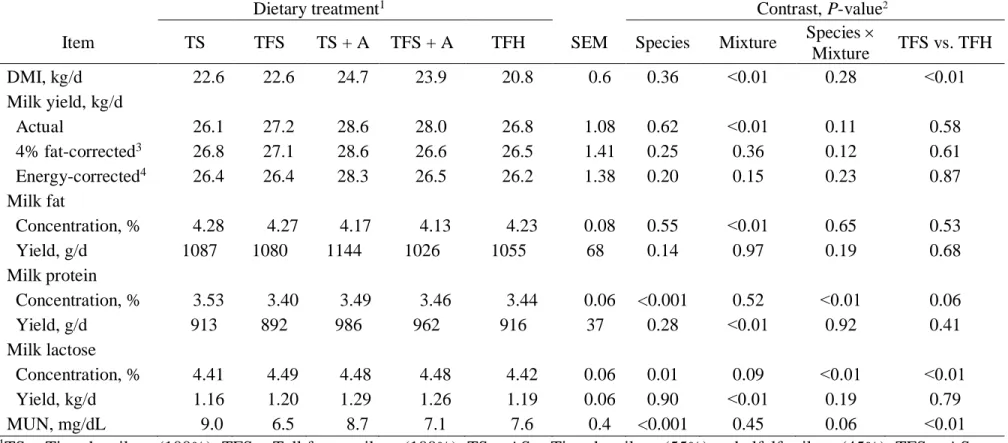
Tableau 5.3. Chemical composition, digestibility and particle length of the dietary treatments ………...58
Tableau 5.4. Effect of experimental diets on dry matter intake, milk yield and composition ………60
Tableau 5.5. Effect of experimental diets on ruminal pH, NH3-N and VFA content ....61
Liste des figures
Figure 1.1 Le modèle expliquant le contrôle de la prise alimentaire par la théorie de l’oxydation hépatique……….15
Liste des abréviations
C : Degrés Celcius
ADF : Fibres insolubles au détergent acide AG : Acides gras
AGMI : Acides gras mono-insaturés AGV: Acides gras volatils
aNDF : Fibres insolubles dans le détergent neutre dosées avec de l’alpha-amylase ANP : Azote non-protéique
BW : Body weight
CAT : Cycle de l’acide tricarboxylique CCK : Cholécystokinine
CVMS : Consommation volontaire de matière sèche DM : Dry matter
DIM : Day in milk DMI : Dry matter intake ECM : Energy corrected milk
eNDF : Fibres insolubles au détergent neutre efficaces ENL : Énergie nette de lactation
ET : Écart-type
FCM : Fat corrected milk
GLP-1 : Glucagon-like peptide 1 GNF : Glucides non-fibreux GNS : Glucides non-structuraux JEL : Jours en lactation
MG : Matière grasse MS : Matière sèche
MUN : Azote uréique dans le lait NAN : Azote non-ammoniacal
NDF : Fibres insolubles au détergent neutre
PB : Protéines brutes
peNDF : Fibres insolubles dans le détergent neutre physiquement efficaces RR : Réticulo-rumen
RTM : Ration totale mélangée SD : Standard deviation VFA : Volatile fatty acids
Remerciements
Tout d’abord, je tiens à remercier les partenaires financiers de ce travail qui, sans eux, n’aurait pas pu être possible. Ce projet de recherche a été financé par l’action concertée FRQNT-Novalait-MAPAQ. Le Centre de recherche en sciences animales de Deschambault a aussi contribué à la réalisation du projet et j’aimerais souligner le travail minutieux de ses employés grâce à lequel j’ai obtenu des données impeccables. Ces gens m’ont fait évoluer en tant qu’agronome, mais surtout en tant que personne. Mon séjour à leur côté est à jamais gravé dans ma mémoire et dans mon cœur. Annie, André, Jonathan, Michaël, Michel, Sébastien et tous ceux que j’ai côtoyés pendant ces quelques mois, mille mercis !
J’adresse également mes remerciements à Édith Charbonneau, ma directrice de recherche. Ta générosité à partager tes connaissances, ton écoute, ta disponibilité, ton ouverture d’esprit, ton amour pour la profession et tes commentaires on ne peut plus pertinents m’ont permis de grandir et d’acquérir une multitude de connaissances. Je te suis infiniment reconnaissante.
Aussi, je tiens à remercier Rachel Gervais, ma co-directrice de recherche. En fait, à mes yeux, Rachel, tu es excessivement significative et le rôle que tu as joué dans ma vie a été des plus déterminants. Mon séjour au sein de ton équipe de recherche au premier cycle a été l’élément déclencheur, l’étincelle qui a fait naitre en moi le désir de poursuivre mes études au deuxième cycle. La professionnelle que tu es, l’alliée que tout le monde voudrait avoir à ses côtés, la maman dévouée, la directrice de recherche à l’écoute et toujours prête à aider, malgré les mille et une choses qui attendent sur le coin de ton bureau, bref, la femme que tu es est celle que j’aurais rêvé d’être. Encore aujourd’hui, j’ai le réflexe d’aller vers toi pour des conseils, du support ou de l’écoute. Ma vie est rocambolesque (et le mot est faible !) par moment et ta générosité, ton ouverture d’esprit et ton non-jugement me font oublier ta notoriété. Les mots me manquent Rachel pour témoigner de l’impact que tu as eu dans mon cheminement. Le seul regret que je puisse avoir est celui de ne pas t’avoir rencontrée plus tôt dans ma vie. Encore une fois, merci Rachel.
Deux personnes ont également été bien significatives dans mon parcours et je parle de Gaëtan Tremblay et Gilles Bélanger. Vos commentaires toujours très pertinents, votre énorme bagage de connaissances et votre professionnalisme ont bonifié tous les documents que je vous ai transmis et m’ont fait évoluée en tant qu’agronome. Mille mercis pour votre engagement !
De façon plus générale, je tiens à adresser mes remerciements à toutes les personnes du département de sciences animales qui ont partagé mon quotidien pendant ces deux années. Je pense à tous les professeurs, chargés de cours, professionnels de recherche ou techniciens de laboratoire. Doris P., Dany C.-M., Jean B., Robert C., Marie-Eve B., Yolaine L. et Nancy B., vous êtes de ceux qui ont ensoleillé mes journées, vous avez partagé vos connaissances si généreusement, vous m’avez soutenue pendant mes moments plus difficiles, vous êtes des personnes d’exception.
Je tiens à remercier également tous les étudiants qui ont travaillé sur mon projet : Catherine et Raphaël au CRSAD, tout comme Carmen, Andréa, Heïdi et Any-Pier au laboratoire, je vous suis très reconnaissante.
De plus, quelques personnes ont une place toute spéciale dans mon cœur. Mon papa a rejoint les nuages quelques jours seulement après le début de ma phase animale. Ma vie s’est alors arrêtée pendant quelque temps, le temps de retomber sur mes pieds et de retrouver mon équilibre. J’ai eu besoin de mettre ma vie sur pause un moment, mais le projet, de son coté, se poursuivait. Les animaux avaient besoin d’être nourris et produisaient toujours du lait ! Des collègues et amis se sont alors offerts pour me remplacer et ce, si spontanément et généreusement que j’en suis encore émue aujourd’hui. Maxime, non seulement tu as été là pendant mon absence, mais tu m’as accompagnée à toutes mes collectes de liquide ruminal parce que j’avais besoin d’aide. Tu es un homme fort au cœur tendre, mille mercis d’avoir été là pour moi. Véronique, ma confidente si attentionnée et intelligente ! Une dizaine d’années nous séparent, mais tu as cette maturité qui me les faisaient oublier à tout coup ! Mille mercis d’avoir été là pour moi ! Marguerite, toi mon mystère sur deux pattes ! Une si petite femme et si grande à la fois. Tu es assurément la
personne qui m’aura le plus fait réfléchir durant mes années à l’université. De la collègue de travail dans le laboratoire au baccalauréat jusqu’à ma « coloc », presque partenaire de vie durant nos maitrises, tu m’as fait grandir en tant que femme, tu as une place spéciale dans mon cœur. Mille mercis d’avoir été là pour moi ! Stéphanie, si tu savais combien je me voyais en toi une douzaine d’années en arrière ! Tu me ressembles, mais avec tellement plus d’intelligence et de prévoyance. À tous les jours durant ces deux années, tu as été ma confidente, « my partner in crime » avec nos cafés ou diners improvisés, nos conversations que nous croyions faire à voix basse, alors que tous nos collègues nous détestaient à force de se faire déranger. Je ne me suis jamais sentie coupable tu sais, le destin avait décidé de nous placer côte à côte dans ce local ! Peggy-Sue, mille mercis d’avoir été là pour moi !
Finalement, je ne serais pas en train d’écrire ces lignes aujourd’hui si je n’avais pas reçu le support et l’amour de mes proches et de ma famille. Tout d’abord, mes enfants, Pénélope, Olivia et Charles, vous êtes ma source d’inspiration qui me pousse à devenir une meilleure version de moi-même un peu plus à tous les jours. Vous êtes mes amours ! Maman, François, Marie-Josée, Dominique, Éric et Caroline, après tant d’années de mauvais choix et de déceptions, j’espère que vous aurez enfin un sentiment de fierté après avoir lu ces pages. Je les ai écrites à cette fin, pour panser un peu les blessures du passé. Je suis convaincue que papa est témoin de tout ça là-haut et c’est aussi à lui que je dédie ce travail. Je vous aime !
Avant-propos
Ce mémoire contient un chapitre rédigé sous forme d'article scientifique. Je suis l'auteure principale de cet article et les coauteurs sont les chercheurs impliqués dans les travaux de recherche. L’article sera soumis pour publication dans la revue « Journal of Dairy Science » sous la référence suivante: « Performance of lactating dairy cows fed tall fescue or timothy as sole forage of the diet or in association with alfalfa », A.-M. Richard, R. Gervais, G. F. Tremblay, G. Bélanger et É. Charbonneau.
Introduction
Les espèces fourragères sont sélectionnées en fonction des caractéristiques climatiques des régions de culture. Au Québec, le climat est de type continental humide avec des étés tempérés (Gosselin et Bélanger, 2010). Ainsi, la fléole des prés est une plante fourragère très représentée dans les champs du Québec; c’est une espèce qui offre une excellente valeur nutritive, de très bons rendements, une survie hivernale supérieure et une grande palatabilité (Bélanger et al., 2006). Par contre, ces caractéristiques sont optimales dans un climat tempéré avec des températures estivales relativement basses, c’est-à-dire entre 17C et 21C le jour (Bertrand et al., 2008). Depuis quelques années, le climat subit des modifications qui sont déjà perceptibles dans le sud-ouest du Québec. Les conséquences des changements climatiques prévus sont une hausse des températures estivales et hivernales, une plus grande accumulation des degrés jours (Qian et al., 2013) et une modification de distribution des précipitations (Qian et al., 2010). Selon Bertrand et al., (2008), une hausse des températures de l’air de l’ordre de 2 ou 3C diminuera de façon substantielle les rendements de la fléole des prés et sa valeur nutritive. Ces conséquences négatives risquent d’être exacerbées en considérant l’évapotranspiration supplémentaire causée par des températures plus élevées et des périodes de sécheresse plus fréquentes (Bertrand et al., 2008). De ce fait, la fléole des prés risque de perdre son avantage actuel sur les autres graminées fourragères et de souffrir davantage de ces modifications climatiques dans les prochaines années.
Afin de faire face aux changements climatiques, les producteurs de fourrages doivent s’adapter. Une solution qui est envisagée est de remplacer la fléole des prés par une autre graminée fourragère mieux appropriée pour croître dans un climat plus sec et plus chaud et qui offre la même valeur nutritive que la fléole des prés. Plusieurs graminées sont couramment utilisées en Europe et aux États-Unis, dont la fétuque élevée. Cette espèce fourragère a une très grande tolérance à la sécheresse et à la chaleur et sa valeur nutritive est appréciable (Peratoner et al., 2011). Par contre, elle est considérée par les producteurs laitiers québécois comme étant moins sapide que plusieurs autres graminées. La résistance
accrue du feuillage à la rupture pendant la mastication, certains constituants volatils et traits morphologiques de la fétuque élevée affectent négativement sa palatabilité (Scehovic et al., 1985). Aussi, selon ce même auteur, la concentration importante de composés soufrés dans la fétuque élevé pourrait être une des raisons de l’infériorité relative de l’appétibilité de cette espèce.
Dans la littérature, un essai a été mené afin de valider les effets du mode de conservation de différents cultivars de fétuque élevée (à feuilles molles et à feuilles rugueuses) sur sa palatabilité (Peratoner et al., 2011). Il a été noté qu’elle est minimale au pâturage, intermédiaire sous forme de foin et maximale en ensilage, et des différences subsistent entre les différents types de feuillage, tous modes de conservation confondus. Par contre, les données scientifiques sont rares en ce qui a trait aux différences entre les types d’ensilage de fétuque élevée (plus ou moins humides, fibres plus ou moins longues). Les producteurs laitiers offrent une panoplie d’ensilages à leurs animaux en fonction des outils qu’ils ont à leur disposition pour la récolte et la conservation des fourrages. Ces informations deviennent donc essentielles pour la sélection des espèces à cultiver.
En plus des graminées, les légumineuses sont intégrées dans la diète des vaches en lactation. La luzerne est largement utilisée dans les mélanges fourragers au Québec (Allard et al., 1998). Par exemple, pour l’année 2016, 764 182 kg de semence de fléole des prés et 713 927 kg de semence de luzerne ont été utilisées au Québec (STATCAN, 2017). Grâce à la capacité de fixation symbiotique de l’azote des légumineuses, la quantité de fertilisants azotés pour assurer une production de fourrage optimale est considérablement diminuée (Obaton, 1992). Aussi, grâce à leur teneur élevée en protéines, les légumineuses permettent de réduire le coût des achats de concentrés destinés aux rations des vaches en lactation. Les résultats d’une étude portant sur l’impact de la combinaison de la fléole des prés à la luzerne sur les plans du rendement fourrager, de la valeur nutritive et de la suppression des mauvaises herbes dans les environnements nordiques indiquent qu’il est avantageux de cultiver ces deux espèces en association (Bélanger et al., 2013). Par contre, alors que les rendements de la fléole des prés risquent d’être affectés négativement par les changements climatiques (Bertrand et al., 2008), ceux de la luzerne devraient augmenter (Charbonneau
et al., 2013) ; la composition botanique du fourrage de ces deux espèces cultivées en association risque donc d’être de moins en moins homogène.
La sélection d’espèces fourragères adaptées aux changements climatiques est indispensable afin d’offrir un mélange fourrager de qualité aux animaux en production. Il est important de réaliser cette démarche aux champs, mais aussi de considérer l’alimentation animale. Ainsi, une plante comme la fétuque élevée soulève l’intérêt des producteurs de fourrages à cause de ses caractéristiques intéressantes pour la culture. Il est toutefois essentiel de s’assurer que cette plante sera consommée par les animaux et qu’elle puisse permettre des performances de production optimales. Plusieurs questionnements existent par rapport à l’appétence de la fétuque élevée et sur les impacts de son utilisation en production laitière. Cette étude permettra de vérifier si cette graminée fourragère est une alternative intéressante à la fléole des prés dans les rations des vaches en lactation.
1. Influence des fourrages sur la prise alimentaire des vaches
Un des principaux facteurs déterminant la production laitière est la prise alimentaire. En effet, pour une production laitière optimale, la consommation de matière sèche doit être maximale. Plusieurs définitions peuvent être attribuées à la notion de prise alimentaire. La prise alimentaire est un comportement complexe, périodique, grâce auquel un organisme tire de son environnement les ressources nutritionnelles nécessaires à la survie (Bellisle, 2005). La régulation de la prise alimentaire est un mécanisme complexe qui engage de nombreux signaux parfois redondants permettant d’adapter un organisme aux variations de la disponibilité en nutriments afin de maintenir l’homéostasie énergétique (Luquet, 2008). Il a été démontré que la régulation se fait au niveau du maintien énergétique, donc de façon quantitative, mais également de façon qualitative, de sorte que l’animal oriente son choix de nutriments vers une compensation de carence potentielle. Évidemment, en situation d’élevage, la régulation qualitative est plus ou moins possible, puisque, d’une part, l’animal n’a pas nécessairement la liberté de choisir ses aliments, et de l’autre, les rations sont formulées en fonction des besoins des animaux. Par contre, la nature des aliments offerts influence la quantité que les animaux vont ingérer. Ces notions sont donc imbriquées une dans l’autre, s’influençant mutuellement.
La prise alimentaire est gérée par l’animal qui reçoit des signaux et les intègre de façon à orienter ses comportements. Aussi, les aliments offerts aux animaux influencent la quantité qui sera ingérée (Faverdin et al., 2007). Des préférences de toutes sortes, allant des caractéristiques physiques jusqu’à la composition chimique des aliments incitent ou nuisent à leur consommation. Le contrôle de la prise alimentaire est très complexe et finement régulé, où chacun des signaux est un élément dans l’équation permettant l’homéostasie et l’homéorhésie.
1.1. Impact de l’espèce fourragère
1.1.1. Appétence du fourrage
La fléole des prés est un aliment utilisé depuis des décennies en production laitière grâce, entre autres, à sa sapidité. À l’opposé, la fétuque élevée est souvent écartée puisqu’elle est moins sapide que plusieurs autres graminées fourragères. Comment peut-on expliquer cette différence ? En quoi un fourrage devient plus ou moins appétent qu’un autre ? Une étude de Fisher et al. (1993) a été menée afin de comparer deux ensilages à base de fétuque élevée et de dactyle pelotonné. Les vaches ont consommé significativement davantage de matière sèche (MS) avec la fétuque élevée. Dans cette même étude, des essais de préférences alimentaires ont aussi démontré que les vaches ont préféré la fétuque élevée au dactyle. Plusieurs paramètres peuvent expliquer ces résultats, dont les teneurs différentes des deux espèces en protéines brutes, en fibres insolubles au détergent neutre (NDF), en fibres insolubles au détergent acide (ADF) et en MS. De plus, une étude de Tava et al. (1995) a été menée afin de comparer la palatabilité de plusieurs cultivars de fétuque élevée. Une relation précise entre les préférences alimentaires des ruminants pour certains cultivars de fétuque élevée et leurs profils en composés volatils n’a pas pu être mise en évidence. Par contre, il a été démontré que la morphologie du feuillage et les changements saisonniers de la composition chimique de la matière sèche, particulièrement au niveau de la protéine et du contenu en sucres solubles, affectent négativement la palatabilité de la fétuque élevée (Scehovic et al., 1985). Plusieurs facteurs influencent l’appétence et, par ricochet, les quantités ingérées des fourrages. Par exemple, les teneurs en fibres, protéines brutes et sucres solubles peuvent expliquer les préférences alimentaires pour certaines espèces ou types de fourrage. Il est intéressant de s’y pencher afin de moduler la régie de culture en conséquence.
1.1.2. Quantité de fibres
Le matériel végétal est composé d’un ensemble de nutriments, tels que les protéines, les lipides, les glucides non-structuraux, les glucides contenus dans les parois cellulaires (NDF), les vitamines et les minéraux (Berthiaume et al., 2015). La teneur en NDF de l’aliment est un élément majeur qui affecte la consommation volontaire de matière sèche (CVMS) des ruminants (Mertens, 1987). En effet, les aliments fibreux doivent rester plus longtemps dans le rumen et le transit est, de ce
fait, ralenti. Un débit plus lent du bolus alimentaire cause une distension d’un ou de plusieurs segments du tractus digestif, résultant en une baisse de prise alimentaire (Allen, 1996). L’encombrement d’un fourrage est proportionnel à son temps de séjour dans le rumen. Le fourrage doit être digéré par les micro-organismes pour réduire la grosseur des particules et être ensuite évacué dans la suite du tube digestif (Allen et Bradford, 2006; Baumont et al., 2009). Le temps de séjour est donc lié à la teneur en parois cellulaires du fourrage (Baumont et al., 2000). À valeur énergétique égale, les légumineuses sont moins encombrantes que les graminées car leur teneur en parois végétales est plus faible (Baumont et al., 2009). Avec un contenu en NDF plus faible que les graminées, la consommation des légumineuses est par le fait même avantagée. Plusieurs études ont démontré que les performances laitières des vaches sont meilleures lorsqu’elles sont nourries avec de l’ensilage de légumineuses comparativement à de l’ensilage de graminées (Thomas et al., 1985; Hoffman et al., 1998; Dewhurst et al., 2003b). Aussi, Steinshamn (2010) mentionne dans sa revue portant sur l’utilisation des légumineuses dans les rations des vaches laitières que la CVMS et la production laitière sont supérieures avec des rations composées de fourrage de légumineuses.
Bien que la teneur en NDF d’un fourrage soit un bon indicatif de son effet d’encombrement (Mertens, 1987), la longueur des particules et leur fragilité, tout comme la digestibilité des fibres NDF sont d’autres facteurs à considérer (Allen, 1996). Selon Berthiaume et al. (2015), la digestibilité in vitro des fibres NDF (NDFd) est considérée comme étant le meilleur paramètre pour estimer la qualité du fourrage, puisqu’elle est étroitement reliée à l’ingestion et aux performances animales. À teneur en NDF égale, le fourrage ayant une NDFd supérieure aura un temps de séjour moins élevé dans le rumen et favorisera ainsi la CVMS de l’animal (Mertens, 2009). Ainsi, bien que les graminées contiennent généralement plus de parois cellulaires que les légumineuses, leurs fibres sont plus digestibles (NDFd plus élevée) (Buxton et Redfearn, 1997).
Quant à la fétuque élevée et la fléole des prés, ce sont toutes deux des graminées fourragères et leur teneur en NDF est similaire, tout comme leur digestibilité, à stade de développement égal. Par contre, la digestibilité des feuilles de fétuque élevée régresse plus drastiquement et plus rapidement au fur et à mesure qu’elle approche son stade reproductif (Easton et al., 1994), ce qui veut donc dire que la NDFd diminue de façon plus importante que celle de la fléole des prés. C’est possiblement un aspect de sa culture qui rebute également les producteurs à son utilisation, puisque
le respect de sa période de coupe est un élément primordial pour optimiser la qualité et l’appétence de ce fourrage.
1.1.3. Teneur en protéines
La teneur en protéines d’un aliment influence l’ingestion de l’animal. Les travaux de Provenza (1995) à ce sujet indiquent qu’une teneur faible en protéines réduit la fermentation ruminale par les micro-organismes, ce qui ralentit le taux de passage et, par ricochet, la prise alimentaire du ruminant. À l’opposé, des rations trop riches en protéines ou en azote non-protéique conduisent à une production excessive d’ammoniac. Cet excès d’ammoniac traverse les parois du rumen, est libéré dans le sang et peut causer une potentielle toxicité, ce que l’animal tente d’éviter en diminuant sa prise alimentaire (Forbes, 2007). En somme, l’ingestion d’une quantité excessive ou non-suffisante de protéines alimentaires induit une diminution de la CVMS.
En contexte de production laitière, les programmes alimentaires sont formulés en fonction des besoins de la vache, selon son stade de lactation et sa production laitière, entre autres. De façon générale, les animaux reçoivent donc des rations équilibrées au niveau des protéines, ce qui évite les carences ou les excès. Par contre, en situation de choix, lorsque l’animal peut orienter ses apports alimentaires, dans un pâturage contenant un mélange fourrager de graminées et de légumineuses par exemple, il a été démontré que les ruminants consomment des quantités de chacune de ces familles afin d’optimiser l’efficacité de la synthèse de protéines microbiennes (Rutter, 2006). En effet, l’auteur de cette revue explique que les différences inhérentes des graminées et des légumineuses, notamment au niveau du carbone (C) et de l’azote (N), justifient les stratégies de sélection des ruminants.
Dans la littérature, certains travaux démontrent que les vaches laitières orientent leur consommation de fourrage en fonction de leurs besoins en protéines dégradables dans le rumen (Tolkamp et al., 1998) ou en fonction de leurs besoins en protéines selon leur stade de production (Kyriazakis et al., 1999). De façon générale, il est raisonnable de penser que la vache laitière ajuste ses apports alimentaires protéiques afin de rencontrer ses besoins, en évitant les excès qui impliqueraient des dépenses énergétiques supplémentaires pour les métaboliser. Par rapport à la
fléole des prés et la fétuque élevée, leur teneur en protéines respective est tributaire de plusieurs facteurs, comme par exemples les apports fertilisants, le cultivar, le fait qu’elles soient cultivées en espèce pure ou en mélange, et les conditions climatiques pendant leur séchage au champ. Il devient donc très difficile de généraliser et de relier l’appétence ou la prise alimentaire à la teneur théorique en protéines de ces plantes. Les analyses de laboratoire sont nécessaires pour obtenir la composition précise des fourrages afin de produire un programme alimentaire adapté.
1.1.4. Les glucides non-structuraux
Les glucides non-structuraux (GNS) incluent le sucrose, le glucose, le fructose, l’amidon et les fructosanes (graminées) ou le pinitol (légumineuses) (Pelletier et al., 2010). Ils sont une source d’énergie rapidement fermentescible dans le rumen. Ils sont positivement corrélés à la qualité d’un fourrage puisqu’ils améliorent l’utilisation par les micro-organismes des protéines dégradables dans le rumen, ce qui augmente la production de protéines microbiennes (Downing et Gamroth, 2007). De plus, la concentration en GNS des fourrages à base de graminées coupés au stade début épiaison serait positivement corrélée avec leur teneur en protéines brutes et négativement corrélée avec leurs teneurs en fibres NDF et ADF (Zhao et al., 2008). Il est connu que les ruminants préfèrent les fourrages qui contiennent davantage de GNS (Fisher et al., 1999) puisque, d’une part, ils reconnaissent et apprécient le gout sucré (Baumont, 1996) et d’une autre, ils préfèrent ingérer des aliments riches en énergie rapidement fermentescibles qui leur procurent une sensation de satiété plus hâtivement (Provenza, 1995). Deux études ont été menées afin de comparer la prise alimentaire de plusieurs cultivars de fétuque élevée. Dans les deux cas, les vaches laitières ont préféré significativement les cultivars ayant une teneur plus élevée en GNS (Tava et al., 1995; Mayland et al., 2000).
La teneur en GNS subit également des variations dans une même journée. En fait, les GNS ont tendance à s’accumuler durant le jour, résultat de la photosynthèse (Fisher et al., 1999; Pelletier et al., 2010). Les ruminants ont d’ailleurs une préférence pour les fourrages coupés en après-midi (Fisher et al., 1999, 2002; Pelletier et al., 2010; Brito et al., 2008). C’est une stratégie qui peut être mise de l’avant pour améliorer la qualité et la palatabilité d’un fourrage.
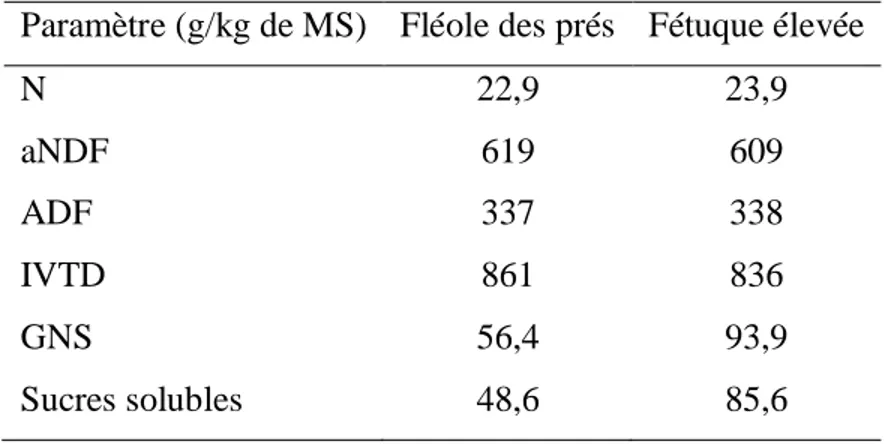
Parallèlement, il est intéressant de comparer la teneur en GNS de la fétuque élevée et de la fléole des prés puisque c’est une caractéristique qui influence la palatabilité d’un aliment. Les concentrations en glucides varient énormément d’une espèce à une autre et la fétuque élevée est généralement avantagée pour cette caractéristique comparativement à la fléole des prés (Mayland et al., 2000; Zhao et al., 2008; Pelletier et al., 2009, 2010). Ces différences de teneurs en GNS sont contradictoires à la faible appétence de la fétuque élevée par rapport à la fléole des prés. Le tableau 1.1 résume les différences entre les valeurs nutritives de la fléole des prés et de la fétuque élevée. La différence dans la palatabilité des deux graminées pourrait être liée à la morphologie du feuillage et à la résistance à la cassure, puisque récoltée au bon stade, la fétuque élevée offre une valeur nutritive semblable à celle de la fléole des prés. Par contre, la fenêtre de coupe pour maintenir ces caractéristiques favorables est considérablement plus petite et le fait que la fétuque élevée soit une graminée non-remontante rend la détection de son stade de développement moins évidente après la première croissance printanière.
Tableau 1.1. Digestibilité in vitro de la matière sèche (in vitro true digestibility, IVTD) et concentrations en azote (N), fibres insolubles au détergent neutre et traitées à l’amylase (aNDF), fibres insolubles au détergent acide (ADF), glucides non structuraux (GNS) et sucres solubles de la fléole des prés et de la fétuque élevée, en première coupe fauchées le matin (Adapté de Pelletier et al., 2010).
Paramètre (g/kg de MS) Fléole des prés Fétuque élevée
N 22,9 23,9 aNDF 619 609 ADF 337 338 IVTD 861 836 GNS 56,4 93,9 Sucres solubles 48,6 85,6
Comme il a été mentionné plus haut, l’ensilage améliore la palatabilité de la fétuque élevée (Peratoner et al., 2011). L’ensilabilité d’un fourrage est corrélé avec sa teneur en GNS. Pendant la fermentation de l’ensilage, ces glucides sont la source principale de substrats pour la fermentation (Downing et Gamroth, 2007). La teneur en GNS améliore ainsi la rapidité de fermentation du
fourrage en ensilage et la chute du pH, ce qui favorise la préservation des nutriments ensilés. La fétuque élevée est donc une plante intéressante à cet effet.
1.1.5. Influence des caractéristiques des fourrages et des nutriments contenus dans la diète sur les signaux de satiété
Les propriétés des aliments offerts aux animaux influencent la quantité ingérée. Chez les ruminants, les caractéristiques physiques de l’aliment comme la taille des particules, la résistance à la cassure et la teneur en MS, le goût et l’odeur influencent la réponse des sens (Baumont, 1996). Par exemple, la morphologie du feuillage de certains cultivars de fétuque élevée affecte négativement sa palatabilité et la prise alimentaire de ce fourrage (Scehovic et al., 1985). Le contrôle de la prise alimentaire est très complexe et finement régulé afin de permettre l’homéostasie et l’homéorhésie de la vache laitière. Des évidences suggèrent d’ailleurs que les ruminants orientent leur choix d’aliments et la quantité qu’ils doivent ingérer en fonction de leurs besoins. En effet, on a longtemps cru que la présence du rumen empêchait les ruminants d’apprendre à associer les propriétés sensorielles d’un aliment avec ses conséquences au niveau du métabolisme, qui est une condition préalable nécessaire à une sélection alimentaire efficace. Par contre, plusieurs évidences démontrent que les ruminants sont en mesure de gérer leurs apports nutritionnels en effectuant des choix appropriés (Forbes, 1995, 2007; Forbes et Provenza, 2000). Pour la majorité des herbivores, il est généralement reconnu que les animaux diminuent leur CVMS avec des aliments de qualité et riches en énergie et en nutriments, puisque leurs besoins sont comblés avec moins d’aliments. À l’inverse, lorsque la qualité des aliments diminue, les animaux en ingèrent davantage (Barboza et al., 2008; Meyer et al., 2010). Dans le cas des vaches laitières en début lactation, les besoins sont tellement élevés qu’il devient impossible de les combler par des aliments. Les apports alimentaires sont alors limités par la capacité physique de l’animal, c’est-à-dire l’encombrement maximal du rumen. Cette théorie nécessite donc d’être ajustée pour ces animaux en contexte de production, poussés aux limites supérieures de leur capacité physiologique.
1.1.5.1. Distension du réticulo-rumen
Comme il a été expliqué plus haut, les ruminants ingèrent des aliments fibreux en très grande quantité pour satisfaire leurs besoins énergétiques. Ces aliments doivent passer à travers l’œsophage de l’animal pour arriver dans le réticulo-rumen (RR), lieu de fermentation microbienne. Dans la couche musculaire du RR, de nombreux récepteurs sont présents, et certains sont stimulés par l’étirement des fibres musculaires des parois ruminales (Harding et Leek, 1972). Une fois excités, les récepteurs envoient des stimuli au système nerveux central qui intègre les informations pour ensuite envoyer des signaux de satiété (Forbes, 1996). Plusieurs mécanismes contrôlent la CVMS et certains dominent sur d’autres, en fonction du statut physiologique, de l’environnement ou simplement de l’individu. Allen (2014) suggère d’ailleurs que la distension ruminale domine le contrôle de la prise alimentaire lorsque les ruminants consomment des diètes faibles en énergie ou lorsque leurs besoins en énergie sont élevés. Il semble toutefois que des changements surviennent en situation de balance énergétique positive, alors que les senseurs d’énergie dans les tissus prennent le relais. Lorsque la CVMS est limitée par la distension du RR, la limitation est affectée par le taux de passage du digesta vers l’abomasum. Ce taux lui-même est influencé par de nombreux facteurs, tels le contenu en NDF, la grosseur et la densité des fibres du digesta, la motilité du RR, les caractéristiques fonctionnelles de l’orifice réticulo-omassal, la vitesse de vidange de l’abomasum (Allen, 2000a), la masse et le volume du digesta (Allen, 1996), tout comme le contenu en eau du digesta et sa capacité de rétention d’eau (Allen, 2014).
En plus, la CVMS serait affectée positivement lorsque la digestibilité de la fibre augmente (Allen, 1996). Selon les explications en lien avec les différences de prise alimentaire entre les rations à base de graminées et de légumineuses, un fourrage plus digeste imposerait une période de fermentation plus courte dans le rumen et le taux de passage serait accéléré. Les fibres des légumineuses ont une moins bonne digestibilité totale que les fibres des graminées à cause de leur plus grande teneur en lignine (Johansen et al., 2017b). Par contre, les fibres digestibles des légumineuses se digèrent plus rapidement que celles des graminées, permettant aux micro-organismes de couper les fibres en particules plus petites plus hâtivement pour ainsi leur permettre de s’échapper du rumen (Dewhurst et al., 2003b) et la distension du RR s’en trouve réduit. En somme, bien que la notion de digestibilité totale des fibres soit importante, la vitesse à laquelle
elle a lieu a autant d’importance pour expliquer la cinétique du digesta et son influence sur la distension du RR.
Une fois que l’encombrement du rumen diminue, l’intensité des signaux anorexigènes provenant des récepteurs de la couche musculaire diminue également. Parallèlement, la concentration plasmatique de ghréline augmente. La ghréline est le seul peptide produit par le système digestif connu à ce jour pour stimuler l’appétit en accélérant le taux de passage et la vidange gastrique (Allen et Piantoni, 2013). Sa concentration est élevée avant le repas, en période de jeûne et lorsque l’encombrement du rumen est moins important (Wertz-Lutz et al., 2006). À l’inverse, la consommation d’aliments diminue la concentration plasmatique de ghréline (Wertz-Lutz et al., 2006). Bradford et Allen (2008) ont mis en évidence que les vaches en début de lactation ont une ghrélinémie plus importante, période caractérisée par une balance énergétique négative et une mobilisation lipidique considérable. Plusieurs travaux ont été réalisés pour comprendre les mécanismes dans lesquels la ghréline est impliquée, autant au niveau de la prise alimentaire que de la balance énergétique des vaches laitières. Par contre, les informations en lien avec la composition des fourrages et son impact sur la ghrélinémie manquent. Puisque la composition des fourrages est associée à l’encombrement du rumen, les caractéristiques qui influencent le taux de passage et la distension du RR doivent assurément influencer la sécrétion de ce peptide, comme la concentration en NDF et leur digestibilité.
1.1.5.2. Cholécystokinine et glucagon-like peptide-1
Le passage du chyme dans le tractus digestif stimule la sécrétion de nombreux peptides, dont la cholécystokinine (CCK) et le glucagon-like peptide-1 (GLP-1). Ces deux incrétines sont des hormones de satiété et seraient impliquées dans le contrôle de la prise alimentaire à court terme (Bradford et al., 2008). Les protéines et les matières grasses alimentaires induisent l’expression et la sécrétion de CCK, avec des concentrations maximales dans le sérum 30 minutes en post-prandial (Choi et al., 2000; Allen et al., 2005). La CCK pourrait affecter la satiété en réduisant la motilité intestinale (augmentant la distension intestinale et la production de propionate dans le rumen) et en agissant directement sur le système nerveux central (Allen et al., 2005). Il a été démontré que les vaches nourries avec une diète riche en matières grasses ont une prise alimentaire inférieure et
une concentration plasmatique de CCK plus élevée que les vaches nourries avec une diète témoin (Choi et Palmquist, 1996). Aussi, la CCK endogène est un régulateur anorexigène qui induit la satiété chez les vaches nourries avec des rations riches en gras (Choi et al., 2000), en diminuant le taux de passage et stimulant ainsi les récepteurs du RR (Allen, 2000b). Parallèlement, des essais ont été réalisés afin d’évaluer les effets factoriels d’une diète à la fois riche en fibres et en matières grasses. Il n’y a pas eu d’interaction entre les effets du gras et des fibres sur la prise alimentaire dans quatre essais (Jerred et al., 1990; Klusmeyer et al., 1991; Tackett et al., 1996; Choi et Allen, 1999) sur six (Grant et Weidner, 1992; Elliott et al., 1995). Lorsqu’il y avait des interactions, les effets anorexigènes de la matière grasse ont été moins importants dans les rations contenant des teneurs élevées en fibres (Allen, 2000b).
Les matières grasses contenues dans les aliments stimuleraient également les sécrétions de GLP-1 et d’insuline (Holst et Gromada, 2004). Une augmentation de la concentration en insuline affecte le métabolisme hépatique en promouvant l’oxydation du propionate au détriment des sentiers gluconéogéniques. Selon Allen et al. (2005), la fin du repas est probablement reliée à ces évènements, jumelée à une absorption rapide de propionate par le foie.
1.1.5.3. L’oxydation hépatique
Le foie est un senseur d’énergie majeur puisqu’il métabolise une variété de carburants dérivés de la diète et des tissus périphériques. Un signal anorexigène ou orexigène, relié à l’oxydation hépatique, est envoyé au cerveau, dépendamment du statut énergétique de l’animal (Allen, 2014). Cette idée selon laquelle la prise alimentaire est contrôlée par un signal du foie au cerveau, défini par l’oxydation des différents carburants, est appelée la théorie de l’oxydation hépatique. Plusieurs carburants peuvent être oxydés ou peuvent stimuler l’oxydation de l’acétyl-CoA dans le foie des ruminants, dont les acides gras libres non-estérifiés, les acides aminés, le lactate, le glycérol et le propionate (Allen, 2014). Contrairement aux espèces monogastriques, le glucose n’est pas utilisé comme source d’énergie par le foie et son captage hépatique est minimal puisque l’activité de la glucokinase est très basse (Ballard, 1965). Puisque les glucides sont transformés en acides gras volatils (AGV) par le microbiote ruminal chez les ruminants, le foie sert en quelque sorte de manufacture de glucose. Le propionate est son précurseur principal. En plus d’être converti en
glucose, le propionate peut être oxydé et stimuler l’oxydation de l’acétyl-CoA dérivé des autres carburants (Allen, 2000b). Comme le glucose chez les non-ruminants, le propionate est hypophagique (Allen, 2014).
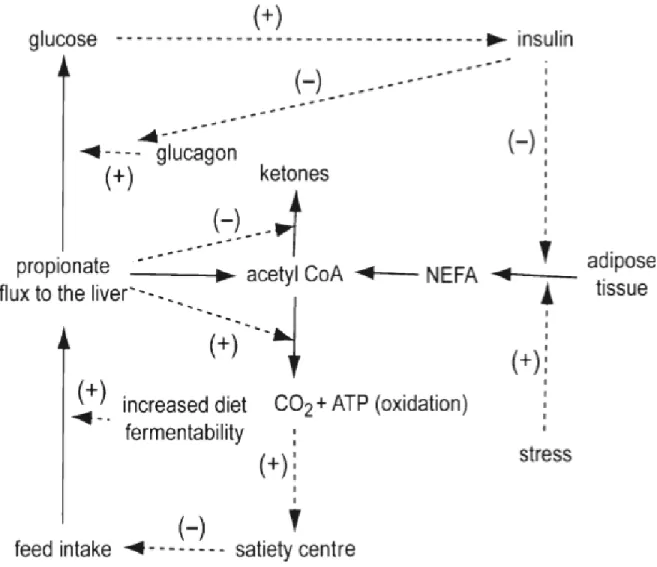
Une fois captés par les hépatocytes, les acides gras non-estérifiés peuvent être oxydés dans les mitochondries ou estérifiés et entreposés en triacylglycérols pour être oxydés plus tard ou exportés en lipoprotéines à très basse densité (Remesy et al., 1986). Les vaches laitières ont une capacité limitée à exporter le triacylglycérol sous forme de lipoprotéines à très basse densité. En début de lactation, elles sont en balance énergétique négative et subissent de la lipolyse de façon importante, elles sont donc sujettes à développer de la stéatose hépatique (Zhu et al., 2000; Allen, 2014). Si l’acide gras non-estérifié est oxydé, chaque cycle de -oxydation libère un acétyl-CoA, qui lui, à son tour, peut être soit oxydé ou exporté en corps cétonique ou en acétate (Allen, 2014). La figure 1.1 présente un schéma qui résume la théorie de l’oxydation hépatique.
Figure 1.1. Le modèle expliquant le contrôle de la prise alimentaire par la théorie de l’oxydation hépatique. Le propionate peut être utilisé par le foie pour produire du glucose en utilisant des ATP, ou peut être oxydé dans le CAT (Cycle acide tricarboxylique) par l’acétyl-CoA, et ceci produit des ATP. L’acétyl-CoA produit par la -oxydation des acides gras peut être oxydé dans le CAT ou être exporté en cétones. Une baisse de la concentration d’insuline, une augmentation de l’insulino-résistance, ou un stress augmentent la lipolyse, ce qui augmente le pool d’acétyl-Coa par la -oxydation des AGNE. Le propionate qui augmente après un repas stimule l’oxydation des acétyl-Coa en CO2, ce qui produit des ATP et ainsi induit la satiété. Tiré de Allen et al. (2009).
Il est important de comprendre que, selon la théorie de l’oxydation hépatique, c’est la capacité d’un carburant à stimuler l’oxydation hépatique (oxydation de l’acétyl-CoA ou du carburant en-soi) qui le rend hypophagique. De ce fait, le propionate est davantage hypophagique que l’acétate. En effet, le captage des AGV du sang au foie est dépendant de la présence de leur acétyl-CoA synthétase respective, et la propionyl-CoA synthétase est beaucoup plus active dans les
hépatocytes que l’acétyl-CoA synthétase (Ricks et Cook, 1981). En plus, le propionate est anaplérotique et stimule l’oxydation de l’acétyl-CoA dans le CAT contrairement à l’acétate (Allen et Piantoni, 2013).
Le glycérol, tout comme le propionate, est un précurseur du glucose mais il utilise un parcours métabolique différent, soit la voie du glycéraldéhyde-3-phosphate. Ainsi, les chances qu’il entre dans le cycle de l’acide tricarboxylique sont moindres, tout comme sa capacité oxydative (Allen, 2014). Les autres carburants anaplérotiques (ex. lactate et les acides aminés glucogéniques) peuvent aussi stimuler l’oxydation hépatique et leur effet anorexigène est ainsi corrélé à leur capacité oxydative respective.
Il est intéressant de mentionner que la cinétique de la digestion (ou fermentation) ruminale détermine le site et le degré de digestion des nutriments, ce qui affecte le type de carburants absorbés en fonction du temps et, par ricochet, la distribution temporelle de leur oxydation hépatique et les comportement alimentaires qui en découlent (Allen et al., 2009). De ce fait, la rétention du digesta dans les pré-estomacs des ruminants permet un approvisionnement constant d’amidon, d’acides gras à longues chaines et de protéines au petit intestin, ce qui augmente le délai d’absorption postprandiale. À l’opposé, les AGV sont rapidement produits et absorbés pendant les repas et induisent donc les signaux de satiété plus tôt (Allen et al., 2009).
1.1.6. Impact de mélanger des graminées avec des légumineuses sur la prise alimentaire
Une méta-analyse récente démontre que les performances de vaches laitières nourries avec différentes espèces de graminées sont comparables, à condition que la digestibilité de la matière organique soit similaire (Johansen et al., 2017a, b). Il est intéressant de constater que l’enjeu principal se situe au niveau de la digestibilité du fourrage, nonobstant ses autres caractéristiques. Parallèlement, il a été démontré, dans la littérature, que les performances laitières des vaches sont meilleures lorsqu’elles sont nourries avec de l’ensilage de légumineuses comparativement à de l’ensilage de graminées (Thomas et al., 1985; Hoffman et al., 1998; Dewhurst et al., 2003b; Johansen et al., 2017b). Dans deux de ces travaux, des comparaisons de prise alimentaire et de performance laitière ont été faites avec des rations contenant des graminées seules, des
légumineuses seules et des mélanges (Dewhurst et al., 2003b; Johansen et al., 2017b). Les résultats de ces essais démontrent que les rations composées de mélanges fourragers (graminées et légumineuses) influencent positivement ces paramètres, et ce, devant les rations de graminées seules. Les travaux d’un autre chercheur ont aussi démontré que la CVMS et la production laitière sont supérieures avec des rations composées de fourrage de légumineuses (Steinshamn, 2010). Les légumineuses contiennent moins de fibres que les graminées et elles se dégradent plus rapidement dans le rumen à cause de la différente position physique de la lignine (Buxton et Redfearn, 1997; Johansen et al., 2017b). Selon les auteurs d’études sur le sujet, c’est ce qui explique majoritairement les différences au niveau des performances laitières des vaches (Buxton et Redfearn, 1997; Johansen et al., 2017b). En somme, pour un même mode de conservation, les ruminants consomment davantage de légumineuses et mélanges fourragers que de graminées seules. Il devient donc avantageux d’ajouter des légumineuses pour optimiser la prise alimentaire et la production laitière des vaches en lactation.
Cela dit, bien que l’ajout de légumineuses dans les rations soit intéressant, l’ensilabilité de ces dernières l’est un peu moins. L’ensilabilité d’une plante est positivement reliée à sa teneur en sucres solubles, négativement reliée à son pouvoir tampon, et elle est optimale entre 35 et 40 % de MS pour les graminées et les légumineuses (Berthiaume et al., 1998; Albrecht et Beauchemin, 2003). Leur concentration élevée en protéine, jumelée à des concentrations relativement élevées en acides organiques et en cations confèrent aux légumineuses un plus grand pouvoir tampon (McDonald et al., 1991; Albrecht et Beauchemin, 2003). Le pouvoir tampon détermine la capacité d’un fourrage à résister à des changements de pH. Or, dans le processus d’ensilage, il est souhaitable que le pH diminue rapidement pour éviter le développement des bactéries butyriques au détriment des bactéries lactiques (Jaster, 1995). De ce fait, un pouvoir tampon élevé n’est pas favorable dans ce contexte.
Aussi, la teneur en sucres rapidement fermentescibles affecte l’ensilabilité d’un fourrage, puisque ces sucres sont le substrat nécessaire à la production d’acide lactique et donc, par ricochet, au déclin du pH (Owens et al., 1999). Les graminées contiennent généralement davantage de sucres solubles que les légumineuses (McDonald et al., 1991; Owens et al., 1999; Pelletier et al., 2010). L’ensilage de légumineuses nécessite une gestion plus serrée : récolter les fourrages à une bonne
teneur en MS, exclure l’oxygène rapidement du silo pour minimiser la respiration aérobie et appliquer une vigilance particulière à l’ouverture du silo sont toutes des stratégies à adopter pour maximiser la qualité d’un ensilage (Albrecht et Beauchemin, 2003). Elles doivent être appliquées plus rigoureusement pour les légumineuses (Albrecht et Beauchemin, 2003).
1.2. Impact du mode de conservation sur la prise alimentaire
1.2.1. Longueur de la fibre
Pour l’animal, la fibre a pour fonction de maximiser la mastication, la rumination et la salivation (Mertens, 2000). Lorsque les fibres sont en excès dans les rations, la densité énergétique est réduite, la CVMS est diminuée et la productivité est à la baisse. À l’inverse, lorsque les fibres sont insuffisantes dans les rations, plusieurs symptômes risquent d’apparaître, allant d’une mauvaise fermentation ruminale jusqu’à l’acidose sévère ou même la mort de l’animal (Mertens, 1997). Malgré le fait que le gras dans le lait soit un paramètre facilement mesurable, une alimentation pauvre en fibres peut potentiellement affecter négativement la santé d’un ruminant sans pour autant causer une chute de gras du lait significative (Mertens, 1997; Zebeli et al., 2006). Le gras du lait ne peut donc pas servir d’informateur unique de la teneur en fibres efficaces d’un aliment. La teneur en NDF seule ne peut pas être utilisée à cette fin non plus, car il est une mesure des caractéristiques chimiques de la fibre, alors que les caractéristiques physiques, telles que la densité ou la longueur des particules, ne sont pas prises en compte par ce paramètre (Mertens, 1997). Pour pallier ce problème, Mertens (1997) a introduit les concepts de fibre NDF physiquement efficace (peNDF) et fibre NDF efficace (eNDF). La peNDF est reliée aux caractéristiques physiques de la fibre qui influencent la mastication et la stratification biphasique du contenu ruminal (tapis de particules longues flottant sur le liquide ruminal rempli de plus petites particules). La eNDF est plutôt reliée à la somme des habiletés d’un aliment à remplacer un fourrage dans une ration afin de maintenir efficacement la production de gras dans le lait.
La longueur optimale de la fibre dans un ensilage doit favoriser à la fois la fermentation dans le silo et la fibre efficace (peNDF) maximale pour la vache (Albrecht et Beauchemin, 2003). Un ensilage coupé finement facilite la compaction dans le silo pour réduire les espaces d’air entre les
particules et améliore l’accès aux sucres solubles par les bactéries lactiques. En contrepartie, un ensilage contenant des fibres plus longues maximise le contenu en fibres efficaces pour le rumen et favorise ainsi son bon fonctionnement (Shinners et al., 1993; Albrecht et Beauchemin, 2003). Aussi, il a été démontré que, pour une même teneur en ADF du fourrage à la mise en silo, les ensilages contenant des fibres plus longues contiennent davantage de fibres ADF que les ensilages composés de fibres plus petites (Ruppel, 1993). Ceci pourrait s’expliquer par la phase aérobie plus importantes pour les fibres longues, résultant en une plus grande utilisation des sucres solubles par les bactéries aérobies et, par ricochet, une diminution de digestibilité et d’énergie pour ces fourrages (Albrecht et Beauchemin, 2003). Si les fibres plus longues ne peuvent pas se compacter autant que les fibres courtes, il y a davantage d’oxygène autour de ces fibres, augmentant de ce fait la période aérobie.
Au niveau de la prise alimentaire, les résultats des études sont divergents. Kononoff et Heinrichs (2003a) ont démontré que la prise alimentaire des vaches augmente avec une diminution de longueur des particules de luzerne offerte sous forme d’ensilage demi-sec, d’autres ne démontrent aucun effet entre la longueur des fibres et la prise alimentaire (Yang et al., 2001; Beauchemin et al., 2003; Bhandari et al., 2007). Dans les travaux de Krause et Combs (2003), la réduction de la longueur de la fibre influence négativement la prise alimentaire des vaches en lactation. Même si une conclusion claire est difficile à tirer de ces travaux, il n’en demeure pas moins que les particules de fourrage trop petites peuvent réduire la mastication, la production de salive et le pouvoir tampon du rumen, ce qui augmente les risques d’acidose ruminale subclinique (Mertens, 1997; Stone, 2004). Il est donc important de trouver l’équilibre entre une longueur de fibre assez courte pour maximiser sa fermentation dans le silo et suffisamment longue pour atteindre une quantité adéquate de peNDF, afin de stimuler la mastication et établir une stratification biphasique du contenu ruminal (Mertens, 2000).
1.2.2. Propriétés physico-chimiques
En plus de la longueur des fibres, d’autres caractéristiques peuvent influencer la fermentation dans le silo et, par conséquent, le produit final qui sera offert comme aliment aux vaches laitières. La matière sèche du fourrage est un bon exemple. L’ensilage peut être conservé plus ou moins
humide, en fonction du temps de séchage imposé à la plante au champ. Un ensilage peut être entreposé humide (30 % MS), préfané (30 à 40 % MS) et demi-sec (40 à 60 % MS). En fonction de l’humidité dans le silo, la fermentation affectera les caractéristiques du fourrage, notamment l’azote et les glucides.
En premier lieu, les légumineuses subissent des changements lors du séchage et de la récolte. La concentration en protéines brutes est habituellement plus élevée pour des ensilages demi-secs de luzerne que pour des foins de luzerne. Cette différence est possiblement reliée à une plus grande perte de feuilles durant la récolte des fourrages de foin comparativement à la récolte d’ensilage des légumineuses (Broderick, 1995; Boutin, 2009). Chez les graminées, le contenu en PB semble être similaire entre un ensilage et un foin (Boutin, 2009), la perte de feuilles au champ est plus négligeable pour cette famille fourragère (Sullivan, 1973). Étant donné que les fourrages destinés à l’ensilage préfané sont récoltés plus humides et donc plus tôt dans le contexte du chantier de récolte que pour les ensilages demi-secs et le foin, il est adéquat de penser que les pertes reliées au séchage sont moindres.
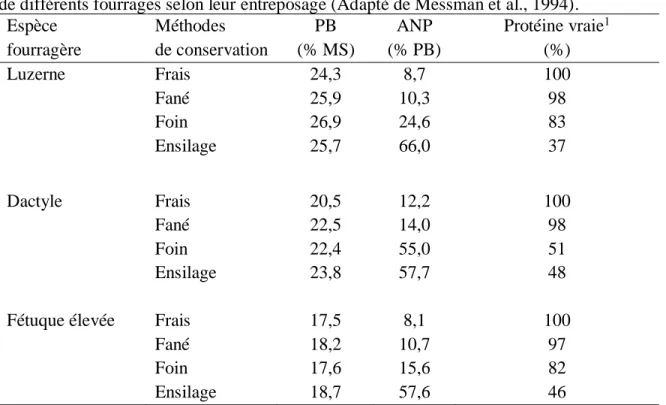
Au niveau des pertes durant l’ensilage, plus le fourrage est sec au moment de la mise en silo, plus les pertes au champ augmentent alors que les pertes lors de l’entreposage diminuent, et vice versa (Collins et Sheaffer, 1996). Une fois la plante coupée, l’activité enzymatique dans la plante cause une augmentation de la proportion d’azote non-protéique (ANP ; peptides, acides aminés libres, amides, nucléotides et l’ammoniac) ; cette proportion peut atteindre jusqu’à 35 % de l’azote total (Rotz et Muck, 1994, Martineau, 2007). Cette protéolyse enzymatique se poursuit jusqu’à la mise en silo ou lorsque la teneur en MS de la plante atteint 60 % (Martineau, 2007). Une fois dans le silo, l’activité microbienne poursuit ce processus biochimique et c’est pourquoi les concentrations en ANP sont plus élevées dans l’ensilage que dans le foin (Messman et al., 1994). Le tableau 1.2 indique les différentes valeurs d’ANP selon l’espèce fourragère et son entreposage.
Tableau 1.2. Teneurs en protéines brutes (PB), en azote non protéique (ANP) et en protéine vraie de différents fourrages selon leur entreposage (Adapté de Messman et al., 1994).
Espèce fourragère Méthodes de conservation PB (% MS) ANP (% PB) Protéine vraie1 (%) Luzerne Frais 24,3 8,7 100 Fané 25,9 10,3 98 Foin 26,9 24,6 83 Ensilage 25,7 66,0 37 Dactyle Frais 20,5 12,2 100 Fané 22,5 14,0 98 Foin 22,4 55,0 51 Ensilage 23,8 57,7 48
Fétuque élevée Frais 17,5 8,1 100
Fané 18,2 10,7 97
Foin 17,6 15,6 82
Ensilage 18,7 57,6 46
1Pourcentage de protéine vraie restante relativement à l’échantillon frais.
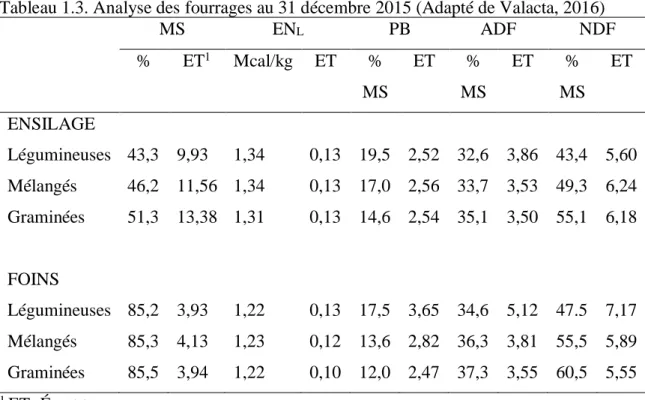
En ce qui a trait aux différences pour les fibres ADF et NDF entre les différents types d’ensilage, les analyses publiées par Valacta (2016) (Tableau 1.3) démontrent clairement que la concentration moyenne en fibres NDF et ADF des ensilages est inférieure à celle des foins, nonobstant la famille fourragère. Ces résultats vont dans le même sens que les explications de Rotz et Much (1994) au sujet de la hausse de concentration des fibres durant le séchage et l’entreposage des fourrages. Les fourrages conservés sous forme de foin à moins de 20 % de MS sont très stables durant l’entreposage.
Aussi, la digestibilité de la MS diminue durant la fermentation et l’entreposage (pour les ensilages), puisque les constituants digestibles sont utilisés pour la respiration et la protéine subit une protéolyse. Pour le foin, la baisse de digestibilité peut davantage être expliquée par la perte de constituants plus digestibles, comme les feuilles, durant la récolte (Savoie, 1988). La perte de GNF et la diminution en digestibilité de la MS induisent une diminution en énergie des fourrages (Rotz et Muck, 1994). Le tableau 1.3 résume bien ce concept.
Tableau 1.3. Analyse des fourrages au 31 décembre 2015 (Adapté de Valacta, 2016) MS ENL PB ADF NDF % ET1 Mcal/kg ET % MS ET % MS ET % MS ET ENSILAGE Légumineuses 43,3 9,93 1,34 0,13 19,5 2,52 32,6 3,86 43,4 5,60 Mélangés 46,2 11,56 1,34 0,13 17,0 2,56 33,7 3,53 49,3 6,24 Graminées 51,3 13,38 1,31 0,13 14,6 2,54 35,1 3,50 55,1 6,18 FOINS Légumineuses 85,2 3,93 1,22 0,13 17,5 3,65 34,6 5,12 47.5 7,17 Mélangés 85,3 4,13 1,23 0,12 13,6 2,82 36,3 3,81 55,5 5,89 Graminées 85,5 3,94 1,22 0,10 12,0 2,47 37,3 3,55 60,5 5,55 1 ET=Écart type
Au niveau de la prise alimentaire pour les ensilages, plusieurs analyses de résultats antérieurs démontrent une relation positive entre la teneur en MS d’un fourrage et la prise alimentaire (Offer et al., 1998; Steen et al., 1998; Wright et al., 2000; Huhtanen et al., 2007). En parallèle, les relations existantes entre la CVMS, la teneur en fibres et la digestibilité des fourrages sont moins stables, probablement à cause des produits de fermentation dans les silos (Huhtanen et al., 2002). En effet, Huhtanen et al. (2002) ont démontré qu’il existe une relation inversement proportionnelle entre l’ingestion d’ensilage et la concentration en azote ammoniacal, en acide lactique, en AGV totaux, en acides gras individuels et acides fermentaires totaux (acide lactique et AGV). À l’opposé, seulement la teneur en glucides solubles a été reliée positivement à la consommation de l’ensilage (Martineau, 2007).
2. Influence des fourrages sur les paramètres ruminaux
2.1. pH ruminal
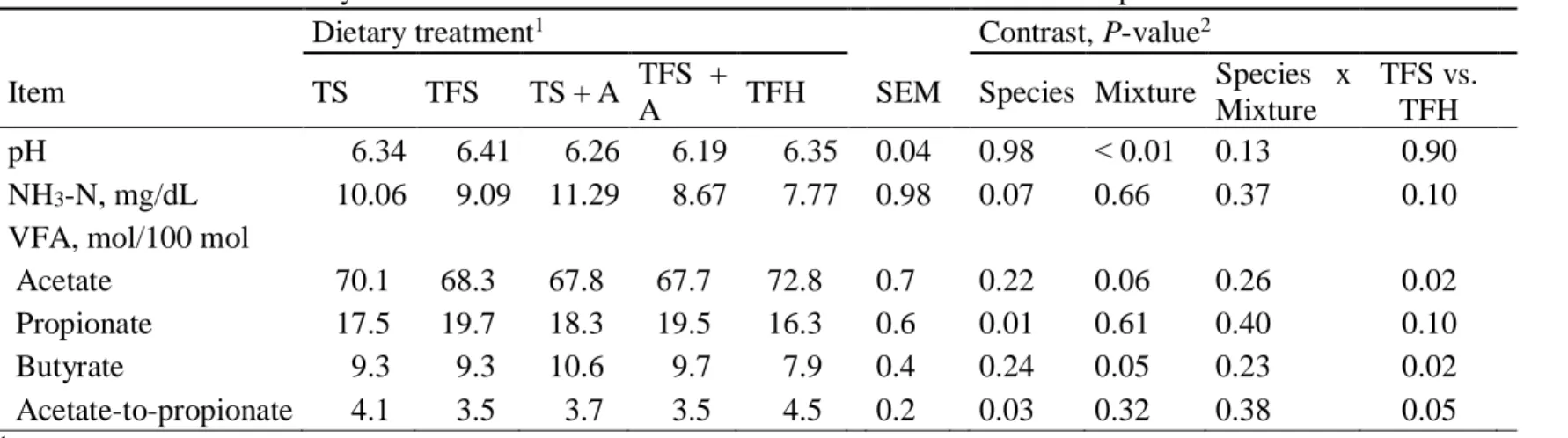
Bien que les ensilages de graminées et de légumineuses aient des caractéristiques propres à leur famille et à leur espèce, une fois ingérés et fermentés dans le rumen des vaches, ils ne semblent pas affecter différemment le pH ruminal. En effet, dans une étude de Dewhurst (2003a), alors que
six traitements composés d’ensilages de graminées seules, de légumineuses seules et de mélanges ont été comparés, les vaches ont maintenu un pH ruminal stable, nonobstant le traitement qui leur était attribué. Les mêmes résultats ont été obtenus dans les travaux d’autres chercheurs (Merry et al., 2006) alors que les vaches ont reçu une des cinq rations expérimentales composées de ray-grass sucré, de ray-ray-grass conventionnel, de trèfle rouge ou de mélange rayray-grass et trèfle rouge. Dans ces études, les rations étaient toutes à base d’ensilage et contenaient peu de concentré. Dans ces circonstances, il est possible de penser que la santé ruminale était favorisée.
Au niveau de la longueur de la fibre, le concept de peNDF est utile pour mesurer l’efficacité de la fibre alimentaire sur la mastication, la salivation et son pouvoir tampon et, par conséquent, le pH ruminal (Yang et Beauchemin, 2009). Par contre, le peNDF ne prend pas en considération les différences de fermentabilité des aliments, ce qui peut affecter considérablement le pH du rumen (Yang et al., 2001; Krause et Combs, 2003; Yang et Beauchemin, 2009). En général, une mastication plus importante, générée par un peNDF alimentaire plus important, fait augmenter le pH ruminal (Krause et al., 2002, Yang et Beauchemin, 2007; 2009). Par contre, pour des rations qui contiennent peu de fourrage, le fait d’augmenter la longueur de la fibre n’atténue pas complètement les effets de baisses de pH causées par les aliments rapidement fermentescibles (Yang et Beauchemin, 2009). Autrement dit, les fibres longues sont bénéfiques pour la santé ruminale, mais lorsque les aliments qui composent les rations sont trop rapidement fermentescibles, les AGV produits peuvent s’accumuler et provoquer une chute du pH ruminal trop importante pour que celui-ci soit rétabli par l’ajout de fibres longues.
2.2. Azote
La prise alimentaire et la production laitière sont généralement supérieures avec des rations composées de légumineuses (Hoffman et al., 1998; Dewhurst et al., 2003b). Le taux de passage de la luzerne est aussi plus rapide que celui des graminées (Waghorn et al., 1989; Dewhurst et al., 2003a, b). Dans les études de Dewhurst (2003a, b), des ensilages de graminées et de légumineuses ont été comparées. Les rations étaient composées de ray-grass, de luzerne, de trèfle blanc et de trèfle rouge offerts seuls et en mélanges (ray-grass et trèfles rouge ou blanc). Tous les ensilages de légumineuses ont généré un flux d’azote non-ammoniacal (NAN) au duodénum plus important,
pouvant être attribué à une tendance à la hausse d’azote microbien. En effet, un taux de passage élevé est associé à une plus grande croissance microbienne (Pirt, 1965). Par contre, à cause de la digestibilité apparente de la matière organique significativement plus faible, la conversion de l’azote dégradable dans le rumen en azote microbien est possiblement assez basse. Le NAN est donc surement relié à une teneur plus élevée en azote non dégradé dans le rumen avec les rations de légumineuses, particulièrement à base de luzerne. Ces excès d’azote dégradable dans le rumen ont mené à une plus faible efficacité de l’utilisation de l’azote (Dewhurst et al., 2003a,b).
Par rapport au mode de conservation des fourrages, il a été démontré que, lorsque consommé par des vaches laitières, le foin résulte en une concentration ruminale en azote ammoniacal plus faible qu’avec de l’ensilage (Broderick, 1995). Ceci pourrait s’expliquer par la plus grande solubilité de la protéine et la dégradation partielle de l’azote protéique en azote non-protéique (ANP) dans l’ensilage comparativement au foin (Martineau, 2007).
2.3. Acides gras volatils
Afin de comparer les performances laitières des vaches nourries avec deux types de graminées, Fisher et al. (1993) ont conduit une étude avec des rations à base d’ensilages de dactyle et de fétuque élevée. La nature de l’ensilage n’a pas eu d’effet sur le pourcentage molaire d’acides gras volatils.
Dans les travaux de Dewhurst (2003a), l’inclusion de luzerne dans les rations à base de graminées a eu l’effet d’augmenter les proportions molaires des acides iso-butyrique, iso-valérique et n-valérique dans le rumen en comparaison aux rations de graminées seules. Ceci pourrait s’expliquer par les teneurs en protéines dégradables dans le rumen plus importantes pour les rations à base de mélanges fourragers. De plus, les rations à base de légumineuses ont fait diminuer les proportions molaires en propionate dans le rumen en comparaison aux rations à base de graminées (Beever et al., 1986; Dewhurst et al., 2003a). Dans les travaux d’autres chercheurs, les rations à bases de légumineuses ont fait augmenter les concentrations d’AGV et les proportions molaires en acétate en comparaison aux rations à bases de graminées (Vanhatalo et al., 2009).