Échapper au conflit sexuel : le cas de Gerris buenoi
Mémoire
Maxime Pineaux
Maîtrise en biologie
Maître ès sciences (M. Sc.)
Québec, Canada
© Maxime Pineaux, 2016
Échapper au conflit sexuel : le cas de Gerris buenoi
Mémoire
Maxime Pineaux
Sous la direction de :
III
RÉSUMÉ
Un conflit sexuel sur le taux d’accouplement est nuisible aux femelles à cause du harcèlement subi et des accouplements superflus. Chez les patineurs, on sait que les femelles résistent aux mâles au cours de luttes pré-copulatoires. Cependant, lutter est aussi coûteux pour elles et en nature les femelles sont plus souvent vues fuyant au large ou se cachant à l’extérieur de l’eau. L’objectif principal de ce projet est de tester si ces comportements sont des moyens alternatifs de résistance aux mâles chez Gerris buenoi. Par ailleurs, il est généralement accepté que les mâles ont très peu de coûts engendrés par les interactions reproductives, mais des études récentes suggèrent le contraire. Un objectif secondaire est de voir s’ils fuient pour éviter la compétition entre mâles pour l’accès aux femelles. Une première expérience s’intéressait simultanément au comportement de fuite des femelles et des mâles. Comme attendu, la probabilité de fuite des femelles et des mâles augmentait avec la proportion de mâles présents dans la population. Pour les femelles, la fuite est donc un moyen de résistance alternatif à la lutte. Étonnamment, les femelles souvent harcelées n’étaient pas celles qui fuyaient, car possiblement trop affaiblies pour être capables de fuir. La probabilité de fuite des mâles augmentait aussi avec la proportion de mâles et les fuyards étaient ceux qui s’étaient rarement accouplés. Ils étaient probablement peu compétitifs et la fuite leur permettait de limiter la compétition entre mâles. La seconde expérience a révélé que certaines femelles se cachaient constamment plus fréquemment à l’extérieur de l’eau que d’autres. Ces différences interindividuelles se maintenaient dans le temps et dans différentes intensités de conflit sexuel, indiquant que ce comportement est un phénotype constant. Se cacher permettait aux femelles de diminuer le harcèlement des mâles, ce qui pourrait leur permettre d’augmenter leur succès reproducteur. Ce projet apporte un regard plus réaliste des interactions intersexuelles existantes chez les patineurs et en particulier des moyens qu’ont les femelles pour justement limiter ces interactions, un aspect du conflit sexuel qui a été très rarement étudié dans le passé.
IV
ABSTRACT
Intersexual interactions linked to sexual conflict over mating rate can be highly detrimental to the fitness of female water strider. It is well established that females resist males mating attempts by engaging in costly pre-copulatory struggles. In nature, females often avoid struggling with males by escaping or hiding off the water, but most studies on sexual conflict have focused on struggles. The main objective of this project was to investigate whether escaping and hiding are alternative female resistance means in Gerris
buenoi. Regarding males, it is generally accepted that they incur few costs associated with
reproductive interactions but recent studies suggest the opposite. A secondary objective was to explore whether males can escape from harsh conditions imposed by male-male competition for access to females. In a first experiment, we simultaneously investigated female and male escape behavior. Probability of females to escape increased with the proportion of males in the population but contrary to predictions heavily harassed females were not escaping more often. As expected, the probability that a female escapes increased with the proportion of males in the population, indicating that escape is an alternative means of resistance to avoid costly conditions imposed by sexual conflict. Contrary to predictions, heavily harassed females were not escaping more often, indicating that they were probably too weak to escape. The probability that a male escapes also increased with the proportion of males, and the males escaping had spent less time mating. These males could be poorly competitive and may have escaped to avoid potential costs imposed by male-male competition. In a second experiment, our results reveal that females differ consistently in their tendency to hide off the water. The tendency to hide was stable over time and across different sexual conflict intensities so that this behaviour can be considered as a consistent phenotype. Importantly, hiding allowed females to lower the harassment they endured, suggesting that this behavioural trait may improve their reproductive success. This project contributes to a better understanding of intersexual interactions in water striders and female resistance means to actually avoid these interactions, an aspect previously neglected in sexual conflict studies.
V
TABLE DES MATIÈRES
RÉSUMÉ ... III ABSTRACT ... IV TABLE DES MATIÈRES ... V LISTE DES TABLEAUX ... VII LISTE DES FIGURES ... VIII REMERCIEMENTS ... X AVANT-PROPOS ... XII
CHAPITRE 1 : Introduction générale ... 1
Le côté femelle du conflit sexuel ... 2
Le côté mâle du conflit sexuel ... 4
Conflit sexuel et personnalité animale ... 5
Les patineurs comme modèles ... 6
Objectifs, hypothèses et prédictions ... 9
CHAPITRE 2: Escaping sexual conflict and male-male competition in Gerris buenoi ... 11
Abstract ... 12
Résumé ... 13
Introduction ... 14
Materials and methods ... 16
Results ... 19 Discussion ... 20 Perspectives ... 23 Tables ... 24 Figures ... 26 Supplementary Material ... 27
CHAPITRE 3: Behavioural consistency in female resistance to male harassment in a water strider species .. 32
Abstract ... 33
Résumé ... 34
Introduction ... 35
Materials and methods ... 38
VI Discussion ... 43 Perspectives ... 46 Tables ... 48 Figures ... 52 Supplementary Material ... 53 CONCLUSION ... 60
ANNEXE A : Persistance des mâles et personnalité animale ... 69
VII
LISTE DES TABLEAUX
Chapitre 2
Table 2.1 Ethogram of observed pre-copulatory behaviours in Gerris buenoi.…………....24 Table 2.2 Probability of escaping as a function of sex-ratio and individual traits for (A)
females and (B) males of Gerris buenoi. β estimates are presented with associated standard errors for explanatory variables of the best model selected on the basis of AICc (Akaike
weight: for females, w = 0.43; for males w = 0.69). A positive effect means an increase in the probability of escaping. The female-biased sex-ratio is used as reference in all models...25
Table 2.S1 Principal components analysis of pre-copulatory behaviours endured by females
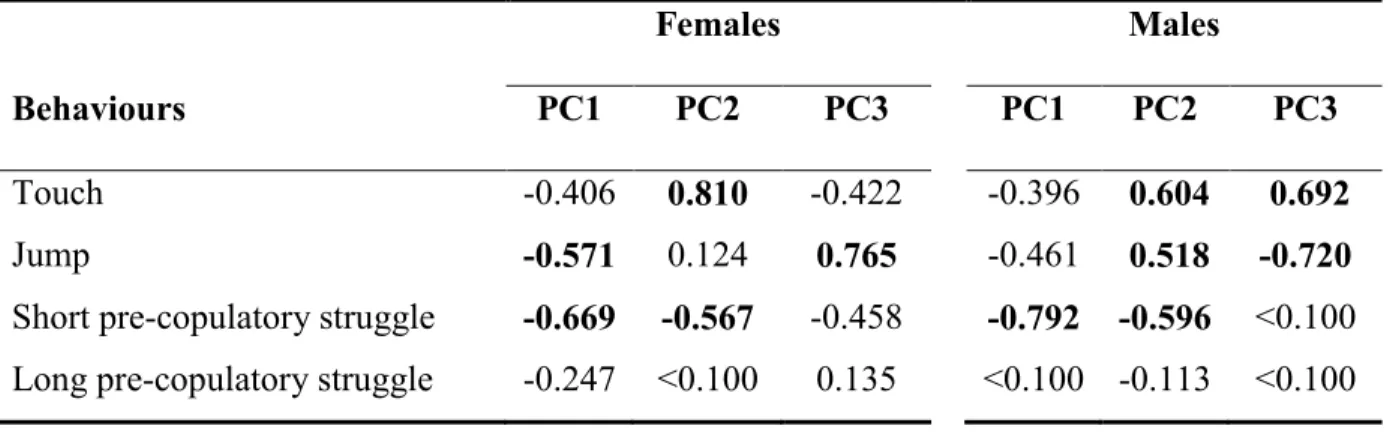
and performed by males of Gerris buenoi. Collectively, the three principal components explain 80.9% of the variance in the female data set and 85.2% of the variance in the male data set. PC loadings > 0.50 are in bold.………..………….….27
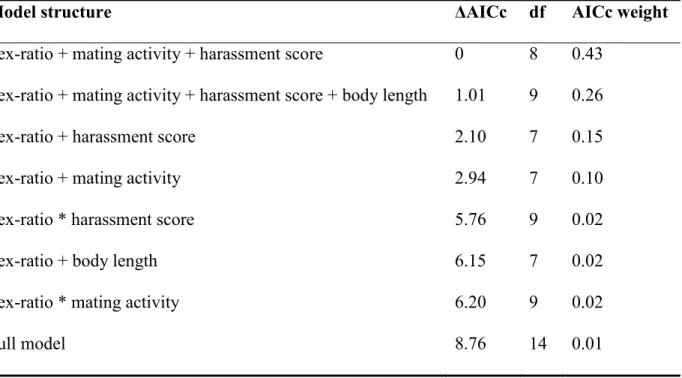
Table 2.S2 AICc model selection table with GLMMs predicting escape behaviour of (A)
females and (B) males of Gerris buenoi. Random effects were included in all models (i.e. the individual effect nested in the pool effect and the observation day effect).………28
Chapitre 3
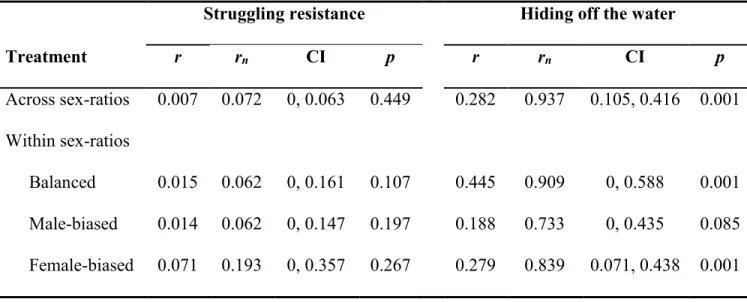
Table 3.1 Ethogram of observed pre-copulatory behaviours in Gerris buenoi..…………....48 Table 3.2 Repeatability values for struggling resistance and the tendency to hide off the water
of Gerris buenoi females………..……….…………49
Table 3.3 Generalized linear mixed models for predictors of (A) female struggling resistance
and (B) female tendency to hide off the water in Gerris buenoi. The balanced sex-ratio was the reference level for the sex-ratio effect.……….………50
Table 3.S1 Repeatability values for the tendency to hide off the water for a brief moment
(during spot checks) of Gerris buenoi females.……….53
Table 3.S2 Generalized linear mixed models for predictors of female tendency to hide off
the water for a brief moment (during spot checks) in Gerris buenoi. The balanced sex-ratio was the reference level for the sex-ratio effect..……….……54
Table A1 Mesures de répétabilité de l’agressivité, de l’efficacité à lutter et de l’efficacité à
entrer en lutte des mâles chez Gerris buenoi.………....74
Table A2 Modèles linéaires mixtes et généralisés prédisant (A) l’agressivité des mâles chez Gerris buenoi, (B) leur efficacité à lutter (C) et leur efficacité à entrer en lutte. Le sex-ratio
VIII
LISTE DES FIGURES
Chapitre 2
Figure 2.1 Probability of escaping in different sex-ratio treatments for females (A) and males
(B) of G. buenoi. The total numbers of females and males observed in the female-biased, balanced, and male-biased sex ratios were 82 and 27, 57 and 55, and 30 and 89, respectively. Bars represent standard errors. Different letters within sex represent significant differences among groups..………..26
Figure 2.S1 Number of escape events observed in males and females of Gerris buenoi (A)
all sex-ratios combined, (B) in the female-biased sex-ratio, (C) in the balanced sex-ratio and (D) in the male-biased sex-ratio.………..…..29
Figure 2.S2 Number of pre-copulatory struggles observed in males and females of Gerris buenoi (A) all sex-ratios combined, (B) in the female-biased sex-ratio, (C) in the balanced
sex-ratio and (D) in the male-biased sex-ratio.………..30
Figure 2.S3 Mean mating activity of males and females of Gerris buenoi (A) all sex-ratios
combined, (B) in the female-biased sex-ratio, (C) in the balanced sex-ratio and (D) in the male-biased sex-ratio.………..…………..31
Chapitre 3
Figure 3.1 Correlation between mean female Gerris buenoi struggling resistance and (A)
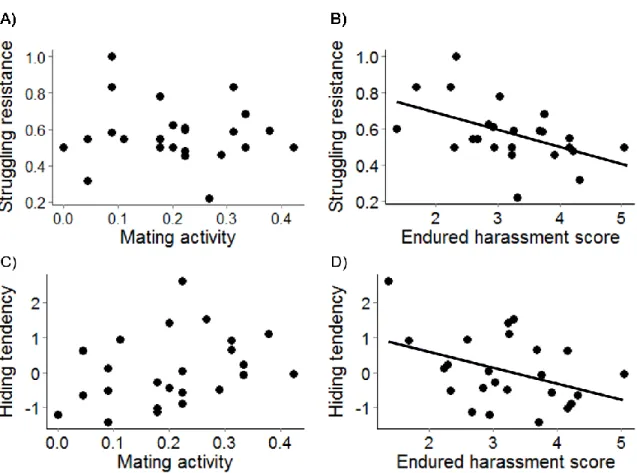
mean mating activity (R = - 0.08, P = 0.72), and (B) mean endured harassment (R = -0.50, P = 0.01); overall tendency of individual female to hide off the water (BLUP) and (C) mean mating activity (R = 0.33, P = 0.11), and (D) mean endured harassment (R = - 0.41, P = 0.05).………...………52
Figure 3.S1 Number of pre-copulatory struggles endured by females of Gerris buenoi (A)
for the whole experiment, (B) in the female-biased sex-ratio, (C) in the balanced sex-ratio and (D) in the male-biased sex-ratio.……….………...………55
Figure 3.S2 Mean endured harassment scores of experimental Gerris buenoi females (A) for
the whole experiment, (B) in the female-biased sex-ratio, (C) in the balanced sex-ratio and (D) in the male-biased sex-ratio.……….………...56
Figure 3.S3 Mating activity of experimental Gerris buenoi females (A) for the whole
experiment, (B) in the female-biased sex-ratio, (C) in the balanced sex-ratio and (D) in the male-biased sex-ratio.………..………..57
Figure 3.S4 Correlation between female average struggling resistance and female overall
tendency to hide off the water (BLUP) in Gerris buenoi (R = -0.02, P = 0.92)..…………..58
Figure A1 Corrélation entre l’activité d’accouplement moyenne des mâles chez Gerris buenoi et (A) leur agressivité (BLUP) (R = 0.52, p = 0.01), et (B) leur efficacité à lutter (R =
IX
The art of dueling is knowing when to fight, when to flee and when to play dirty.
X
REMERCIEMENTS
La première personne que je dois remercier pour son aide à l’accomplissement de ma maîtrise, mais pas seulement, est Julie Turgeon, ma directrice de recherche. C’est elle qui m’a fait découvrir le monde de la recherche, un monde qui m’intéressait depuis plusieurs années déjà. Je me souviens du premier jour où j’ai posé un pied à l’Université Laval, à la recherche, un peu tardivement, d’un professeur avec lequel faire mon initiation à la recherche. Julie était mon premier choix mais je pensais que mes chances étaient faibles étant donné le retard avec lequel je m’y prenais et le fait que j’étais un parfait inconnu fraîchement débarqué sur le Nouveau-Continent. Je veux la remercier pour la confiance qu’elle m’a accordée à ce moment-là. Je dois vous faire part de mon étonnement lorsqu’elle m’a suggéré, quelques mois plus tard, l’idée de rester au Québec pour faire une maîtrise dans son laboratoire. Cette opportunité a clairement renforcé l’ambition que j’avais de poursuivre dans le monde de la recherche. Cette opportunité m’a aussi donné confiance en moi, qualité que je n’avais pas, loin de là, avant ce moment-là. Pendant les trois années qui ont suivi, Julie m’a transmis sa passion et sa rigueur scientifique, par les discussions que nous avons eues, par ses conseils, par son support permanent, par ses critiques et ses encouragements. Tout cela m’a permis de développer toutes les qualités nécessaires pour être un scientifique accompli, mais aussi pour être une personne à l’esprit ouvert. Julie, encore une fois, je te suis extrêmement reconnaissant pour la formation d’une qualité exceptionnelle à laquelle j’ai eu le droit.
Je veux aussi remercier toutes les personnes que j’ai côtoyées au sein du laboratoire de Julie Turgeon. Merci à Geneviève Parent pour m’avoir aidé à faire mes premiers pas en recherche dans le laboratoire, et pour sa bonne humeur contagieuse. Je remercie Éric Devost pour le temps qu’il m’a accordé et qui m’a permis de dompter R, mais aussi pour les discussions toujours pertinentes que nous avons eues. Merci à Pierre Duchesne pour l’aide qu’il m’a apporté dans mon traitement de données, mais aussi pour son humour fin. Je dois remercier Jeanne Piette pour ses commentaires sur mes présentations et pour nos discussions pertinentes et moins pertinentes. Je remercie aussi Joséphine Magnan, Ariane Blier-Langdeau et Gabriel Piette-Lauzière pour l’aide qu’ils m’ont apporté sur le terrain lors de mes expériences.
XI
De manière plus personnelle, je veux remercier mes parents et ma sœur, pour leur soutien et pour l’attention qu’ils m’ont porté depuis le début. Je suis vraiment reconnaissant envers mes parents de m’avoir rendu curieux et de m’avoir donné le goût de la science. Je veux les remercier de m’avoir poussé à donner le meilleur de moi-même et d’aller toujours de l’avant. En termes de soutien, je dois aussi remercier chaleureusement mes amis d’enfance, Antoine, Tony, Guillaume, Laurine et Élodie. L’attention qu’ils m’ont porté, mais aussi leurs calembours de haute voltige, ont réussi à me réconforter dans les moments les plus difficiles. Pour terminer cette note personnelle, il m’est impossible de ne pas remercier Agatha, sans qui j’aurai probablement eu beaucoup de mal à rester à flots ces derniers mois. Aussi incongru que cela puisse paraître, je crois que ce ne sont pas les ornithorynques qui auront le plus marqué mon périple australien. Merci pour ton aide, pour tes délicieux crobars, pour ton utilisation désuète de la langue française, mais surtout pour le soutien infaillible que tu m’as accordé.
Enfin, je tiens à remercier le CRSNG pour les fonds de recherche accordés à ma directrice de recherche.
XII
AVANT-PROPOS
Le présent mémoire est composé d’une introduction et d’une conclusion en français ainsi que de deux chapitres, chacun écrit sous la forme d’un article scientifique et rédigé en anglais. Le premier article (deuxième chapitre) est prévu d’être soumis à Behavioral Ecology, et le second article (troisième chapitre) à Behavioral Ecology and Sociobiology. Les auteurs de ces deux articles sont Julie Turgeon, ma directrice de recherche, et moi-même. Le design des expériences, l’échantillonnage, l’analyse des données et l’écriture des manuscrits ont été principalement, mais non-exclusivement, à ma charge.
L’expérience qui a abouti à l’écriture du troisième chapitre de ce mémoire a apporté des résultats pour les femelles et les mâles, mais seuls les résultats concernant les femelles sont rapportés et discutés dans le troisième chapitre. Les résultats concernant les mâles font quant à eux l’objet de l’Annexe A. Cette séparation vient du fait que les résultats concernant les femelles ont apporté beaucoup de nouveauté, et étaient difficilement conciliables dans un même article avec les résultats obtenus pour les mâles.
1
CHAPITRE 1 : Introduction générale
La reproduction est essentielle dans la vie de tous les organismes. Lorsqu’elle est sexuée, l’union de gamètes provenant d’une femelle et d’un mâle aboutit à la production d’une autre génération. La reproduction a longtemps été vue comme une coopération entre les deux sexes, mais les intérêts des mâles et des femelles dans la reproduction sont en réalité rarement les mêmes (Parker 1979). L’anisogamie est à la base de cette divergence d’intérêts : les mâles produisent un grand nombre de petits gamètes pauvres en ressources, alors que les femelles produisent une faible quantité de gros gamètes riches en énergie (Bateman 1948). De cette anisogamie découlent des différences dans l’histoire de vie des deux sexes, mais aussi dans leur comportement et dans leur morphologie. Il en résulte des interactions intersexuelles et intra-sexuelles de différents types qui créent des variations dans le succès reproducteur des individus. Ces variations sont à la base de la sélection sexuelle. Les femelles ont parfois des préférences cachées pour certains des attributs possédés par les mâles, et ceux-ci peuvent exploiter ces préférences. Cette exploitation peut se faire au détriment des femelles, ce qui peut conduire à la mise en place d’un conflit sexuel (Chapman et coll. 2003). Un conflit sexuel se met en place lorsqu’il existe des pressions de sélection antagonistes entre les mâles et les femelles quant à l’énergie investie dans la reproduction (Parker 1979, 2006). Ce conflit s’exprime sur des traits partagés par les deux sexes. Ces traits partagés peuvent résulter d’interactions entre les mâles et les femelles, comme le taux d’accouplement, la durée des copulations ou la fécondité (Chapman et coll. 2003; Perry & Rowe 2015). Lorsque le conflit sexuel est sur le taux d’accouplement, les mâles et les femelles n’ont pas les mêmes intérêts quant à la décision de s’accoupler. Les mâles vont généralement chercher à s’accoupler le plus souvent possible parce que leur succès reproducteur apparaît comme limité par le nombre d’accouplements. Lorsque c’est le cas, les mâles vont typiquement développer des traits de persistance qui leur permettent de gagner des accouplements (Arnqvist & Rowe 2005). Il existe de nombreux exemples de traits de persistance, qui vont du pénis perforateur chez les insectes à insémination traumatique (Tatarnic et coll. 2006), aux comportements de harcèlement mis en évidence dans une grande diversité de taxons (Magurran & Seghers 1994; Réale et coll. 1996; Shine et coll. 2001; Le Gaillard et coll. 2005). Ces traits de persistance ont souvent un impact négatif sur le fitness
2
des femelles, notamment en entraînant une diminution de leur fécondité (Rönn et coll. 2006; Eldakar et coll. 2011; Makowicz & Schlupp 2013) ou en augmentant leur mortalité (Réale et coll. 1996; Clutton-Brock & Langley 1997; Shine et coll. 2001; Le Gaillard et coll. 2005). Si le conflit sexuel affecte négativement le fitness des femelles, il entraîne aussi une diminution du fitness de l’ensemble de la population. Le conflit sexuel peut donc théoriquement mener à l’extinction de la population, mais ce risque d’extinction peut cependant être empêché par des mécanismes antagonistes (Hardin 1968; Rankin et coll. 2011).
Le côté femelle du conflit sexuel
Parmi les mécanismes pouvant empêcher une extinction de la population, les femelles peuvent développer des traits de résistance antagonistes aux traits de persistance des mâles. Ces traits de résistance peuvent simplement entraîner une diminution des coûts associés à la persistance des mâles (Rowe et coll. 2005; Michels et coll. 2015). Ils peuvent aussi conduire à une baisse du fitness des mâles en permettant aux femelles de limiter leur taux d’accouplement, menant éventuellement à une coévolution antagoniste entre la résistance des femelles et la persistance des mâles (Arnqvist & Rowe 2005). Les traits de résistance des femelles ont été bien moins explorés que les traits de persistance des mâles. Du côté des femelles, les évidences concernent surtout des traits morphologiques (Arnqvist & Rowe 2002; Eberhard 2004; Tatarnic et coll. 2006; Green et coll. 2013), alors qu’il semblerait que les traits de résistance soient plus souvent comportementaux que morphologiques (Eberhard 2004; Perry & Rowe 2015). La résistance comportementale a l’avantage d’être plus plastique que la résistance morphologique, ce qui permet aux femelles de rapidement s’adapter aux conditions locales. Cet avantage est particulièrement important lorsque l’économie du conflit sexuel dépend des conditions environnementales ou sociales (i.e., le risque de prédation : Rowe 1994; le sex-ratio : Le Gaillard et coll. 2005; la densité : Baguette et coll. 1996).
Il existe différents types de résistance comportementale. Elle peut être physique, par un affrontement direct avec les mâles (Arnqvist 1992a; Blyth & Gilburn 2011), ou par dissuasion en montrant de l’agressivité (Forstmeier 2004; Stuart-Fox & Whiting 2005). Ces affrontements peuvent cependant être très coûteux pour elles (Le Gaillard et coll. 2005; Makowicz & Schlupp 2013; Watson et coll. 1998). Pour éviter ces coûts, les femelles peuvent
3
aussi résister aux mâles de façon plus indirecte. Ces formes de résistance indirectes sont beaucoup moins étudiées, mais des évidences montrent que les femelles peuvent par exemple accepter des accouplements superflus (i.e., polyandrie de commodité : Thornhill & Alcock 1983; Rowe 1992; Harano 2015). Les femelles peuvent aussi éviter à la fois les coûts liés au harcèlement et à l’accouplement en allant se cacher là où les mâles vont rarement (Bokides et coll. 2012; Schultze & Fincke 2013; Bots et coll. 2015), ou bien en s’enfuyant ou en dispersant de l’environnement où elles se trouvent (Shine et coll. 2005; Darden & Croft 2008; Eldakar et coll. 2009a).
La fuite des femelles face au conflit sexuel a été très rarement investiguée. Les quelques études qui s’y sont intéressées ont rapporté que les femelles fuyaient lorsque l’intensité de harcèlement des mâles était élevée. Cette forte intensité de harcèlement peut être due à un biais du sex-ratio vers les mâles (Darden & Croft 2008) ou bien à une élévation de la densité (Baguette et coll. 1996). Les femelles peuvent aussi fuir lorsqu’il y a des mâles particulièrement agressifs dans leur environnement (Eldakar et coll. 2009a, 2010).
En plus d’être dépendante des conditions environnementales, la décision de partir ou de rester dépend aussi du phénotype des individus. Tous les individus d’une même population n’ont pas la même réponse face aux conditions locales, et dans la majorité des cas, seule une fraction de la population s’en va. Il existe donc souvent des différences phénotypiques entre les fuyards et ceux qui restent. Ces différences peuvent être liées à l’âge des individus (Bowler & Benton 2005), à leur morphologie (Lawrence 1987; Shine et coll. 2006), ou même à leur comportement (Cote et coll. 2010). Dans le cadre du conflit sexuel, ces différences pourraient aussi être liées aux expériences passées des femelles. Malheureusement, les études qui rapportent des évidences de la fuite des femelles face au conflit sexuel se sont principalement intéressées à l’effet de l’environnement social (sex-ratio, densité). Les quelques études qui se sont intéressées à des mécanismes plus proximaux ont rapporté que les femelles qui s’en allaient pouvaient être celles qui avaient subi le plus de harcèlement (Eldakar et coll. 2010). Au contraire, les femelles les plus harcelées étaient parfois trop affaiblies et incapables de s’enfuir (Le Gaillard et coll. 2005).
4 Le côté mâle du conflit sexuel
L’extinction d’une population causée par le conflit sexuel peut aussi être empêchée si les mâles ont des coûts associés à l’accouplement ou au harcèlement (Rankin et coll. 2011). Les études sur le conflit sexuel sont en très grande majorité concentrées sur les coûts existants chez les femelles et sur les avantages en termes de fitness associés à la persistance des mâles (révisé par Perry & Rowe 2015). Par contre, les coûts potentiels des mâles associés au harcèlement ou à l’accouplement ont été très peu explorés. Cette absence d’intérêt vient du fait qu’il est généralement accepté que les mâles ont peu de coûts associés au conflit sexuel (Arnqvist 1997; Watson et coll. 1998). Des évidences récentes ont quand même rapporté l’existence de coûts à harceler pour les mâles, via une diminution de la condition corporelle (Macowicz & Schlupp 2013) ou du succès reproducteur (Devost & Turgeon 2016).
Les mâles peuvent aussi avoir des coûts associés à la compétition entre mâles pour l’accès aux femelles. L’intensité de la compétition entre mâles et celle du conflit sexuel sont très étroitement liées, notamment parce qu’elles fluctuent de la même façon avec certaines conditions environnementales ou sociales, comme le sex-ratio ou la densité d’une population (Kokko & Rankin 2006; Head & Brooks 2006; Macowicz & Schlupp 2013). Ces deux forces ont pourtant été rarement étudiées simultanément. Lorsque la compétition entre mâles s’intensifie, les mâles peuvent avoir des coûts liés aux confrontations physiques entre mâles (Qvarnström 1997; Savage et coll. 2005). L’intensification de la compétition entre mâles peut aussi entraîner des coûts plus indirects liés à la diminution du succès d’accouplement des mâles, notamment à cause de la baisse des opportunités d’accouplement, mais aussi à cause de l’augmentation de la réticence des femelles (Jirotkul 1999).
Pour éviter ces coûts, les mâles pourraient eux aussi s’enfuir et rejoindre un environnement plus favorable où ils pourraient éventuellement augmenter leur succès d’accouplement. Les quelques rares études qui se sont intéressées à la fuite des mâles ont rapporté que leur probabilité de fuite augmente lorsque le sex-ratio est biaisé vers les mâles, et donc lorsque la compétition entre mâles est intense (Lawrence 1987; Croft et coll. 2003; Trochet et coll. 2013). Tout comme pour les femelles, la majorité de ces études se sont intéressées à l’effet de l’environnement social, ce qui ne permet pas de connaître les particularités phénotypiques des fuyards. Quelques études ont tout de même rapporté que les
5
mâles qui fuyaient le plus étaient souvent ceux qui étaient les moins compétitifs ou qui avaient le moins de succès avec les femelles (Lawrence 1987; Shine et coll. 2006; Okada et coll. 2007).
Conflit sexuel et personnalité animale
Comme dit plus haut, il existe toujours des différences phénotypiques entre les individus d’une même population. Ces différences interindividuelles peuvent être comportementales. Si ces différences sont maintenues au cours du temps et/ou dans différents contextes, on peut considérer le comportement en question comme un trait individuel, plus communément appelé un trait de personnalité (Sih et coll. 2004; Réale et coll. 2007; Bell et coll. 2009; Briffa & Weiss 2010). Un trait de personnalité a une base génétique plus ou moins élevée, et est donc en partie transmis de génération en génération, offrant ainsi la possibilité à ce trait d’être éventuellement sélectionné (Dohm 2002). La personnalité animale a déjà été mise en lien avec de nombreux processus biologiques comme l’immunité, les invasions biologiques, ou la sélection sexuelle (Fogarty et coll. 2011; Sih et coll. 2012; DiRienzo et coll. 2015). Le lien entre la personnalité animale et le conflit sexuel a lui été fait presque exclusivement du côté des mâles. Des différences interindividuelles dans le niveau de persistance comportementale des mâles ont été mises en évidence, notamment au niveau de leur agressivité (Chang & Sih 2013; Montiglio et coll. 2016; Wey et coll. 2015a). Le maintien de ces différences interindividuelles dans l’agressivité des mâles a par contre été investigué uniquement dans le temps, et pas dans différents contextes, comme par exemple différentes intensités de conflit sexuel. Il est possible que les mâles puissent plastiquement changer de niveau d’agressivité en fonction de l’environnement social, et notamment du nombre de femelles disponibles pour l’accouplement par rapport au nombre de compétiteurs mâles (Dingemanse et coll. 2010).
À l’opposé, personne ne s’est jamais intéressé à voir s’il pouvait y avoir des différences constantes dans le niveau de résistance comportementale des femelles. On sait que la résistance comportementale des femelles est affectée par une grande variété de facteurs environnementaux, comme le risque de prédation (Han & Jablonski 2010), la densité (Baguette et coll. 1996) ou le sex-ratio (Darden & Croft 2008). Par contre, on ne sait pas s’il y a aussi une composante intrinsèque à la résistance comportementale des femelles, ou
6
autrement dit s’il y a des femelles plus résistantes que d’autres quel que soit le contexte environnemental ou social. Si tel est le cas, les femelles plus résistantes devraient bénéficier d’un fitness relativement plus élevé que les autres femelles de la population. Cette augmentation du fitness pourrait notamment être causée par une limitation des interactions intersexuelles coûteuses, comme les accouplements ou le harcèlement. En plus d’être un trait de personnalité, la résistance comportementale pourrait être aussi plastique (Dingemanse et coll. 2010). Cette plasticité permettrait aux femelles de s’adapter rapidement aux conditions locales. Par exemple, en réponse à une hausse de l’intensité du conflit sexuel, les femelles pourraient être plus réticentes à s’accoupler pour éviter les coûts associés à l’accouplement (Muhlhauser & Blanckenhorn 2002). Au contraire, elles pourraient aussi être moins résistantes face aux assauts des mâles pour éviter les coûts liés au harcèlement (Thornhill & Alcock 1983).
Les patineurs comme modèles
Les patineurs (Gerridae) sont parmi les exemples les plus connus de l’existence d’un conflit sexuel sur le taux d’accouplement (Arnqvist 1992a; Rowe et coll. 1994; Arnqvist & Rowe 2005; Perry & Rowe 2015). Les mâles harcèlent fréquemment les femelles pour tenter de s’accoupler avec elles (Rowe 1992; Rowe et coll. 1994). Le harcèlement peut se traduire par de brefs contacts initiés par les mâles. Il peut aussi y avoir des contacts plus longs et plus intenses appelés luttes pré-copulatoires, au cours desquelles les mâles s’agrippent aux femelles pour les forcer à s’accoupler. Environ 15% des évènements de harcèlement mènent à une lutte pré-copulatoire (Rowe 1992, 1994). Ces luttes mènent quant à elles à un accouplement dans 8 à 22% des cas, notamment parce que les femelles résistent vigoureusement aux tentatives des mâles pour éviter tous les coûts associés à l’accouplement (i.e., augmentation du risque de prédation et forte dépense énergétique : Arnqvist 1989; Fairbairn 1993; Watson et coll. 1998). L’accouplement peut durer d’une dizaine de minutes chez Gerris buenoi à plusieurs heures chez d’autres espèces comme Aquarius remigis (Rowe 1992; Weigensberg & Fairbairn 1994). Il peut être suivi d’une garde post-copulatoire au cours de laquelle le mâle reste passif sur le dos de la femelle. Cette garde prend fin par une courte lutte à l’initiative de la femelle (Rowe et coll. 1994).
7
La résistance des femelles et la persistance des mâles ont été très bien décrites d’un point de vue morphologique. Certains traits morphologiques permettent aux femelles de repousser les assauts des mâles. C’est notamment le cas des épines connexivales situées au bout de leur abdomen qui leur permettent de réduire la proportion de luttes subies et la fréquence des copulations (Arnqvist & Rowe 1995; Arnqvist & Rowe 2002; Ronkainen et coll. 2005). Du côté des mâles, l’aplatissement de leur abdomen leur permet de s’accoupler plus fréquemment (Arnqvist & Rowe 2002). Il y a des évidences de l’existence d’une coévolution antagoniste entre ces traits de résistance et de persistance. Autrement dit, le niveau d’armement des mâles et des femelles est corrélé entre espèces (Arnqvist & Rowe 2002) et entre populations d’une même espèce (Gagnon & Turgeon 2011; Perry & Rowe 2012).
Outre ces traits morphologiques, il existe aussi des comportements de résistance et de persistance chez les patineurs. La très grande majorité des études sur le conflit sexuel s’est intéressée au comportement de résistance des femelles lors des luttes pré-copulatoires, en la mettant en lien avec certains traits morphologiques (Arnqvist 1992a; Rowe 1992; Arnqvist & Rowe 1995; Arnqvist & Rowe 2002; Ronkainen et coll. 2005; Gagnon & Turgeon 2011; Perry & Rowe 2012). Les luttes pré-copulatoires sont pourtant très coûteuses pour les femelles (Arnqvist 1989; Rowe 1994; Watson et coll. 1998). Lorsque le harcèlement est très intense, dans un sex-ratio biaisé vers les mâles par exemple, il est même plus coûteux de résister physiquement pendant ces luttes que d’accepter des accouplement superflus (i.e., polyandrie de commodité : Rowe 1992). La polyandrie de commodité peut apparaître comme un moyen indirect de résistance pour les femelles, mais cette stratégie ne leur permet pas d’éviter les coûts liés à l’accouplement.
Les femelles ne font pas que subir les interactions avec les mâles, elles peuvent aussi éviter l’affrontement direct et les coûts qui y sont associés. Ces moyens alternatifs de résistance ont été très peu étudiés comparativement à la résistance physique. Ce débalancement vient probablement du fait que beaucoup d’études sur la résistance des femelles ont été menées en laboratoire dans des conditions peu réalistes, avec peu d’individus, dans un espace réduit et/ou en s’intéressant à un seul sexe à la fois (Arnqvist & Rowe 1995; Ortigosa & Rowe 2003). Quelques études se sont tout de même intéressées à la
8
possibilité qu’ont les femelles de se cacher en dehors de l’eau, sur des débris végétaux ou sur les bords d’étang ou de rivière (Krupa et coll. 1990; Krupa & Sih 1993; Rowe et coll. 1994). Les mâles sont rarement aperçus en dehors l’eau, ce qui permettrait aux femelles d’éviter de se faire harceler ou de s’accoupler souvent (Rowe et coll. 1996). Il y a des différences entre les femelles dans leur tendance à aller en dehors l’eau, et ces différences sont constantes au cours du temps (Wey et coll. 2015b). Par contre, on ne sait pas encore si le fait d’aller en dehors de l’eau permet vraiment à ces femelles de se cacher des mâles et donc de limiter les évènements de harcèlement et d’accouplement qu’elles subissent. Si tel est le cas, cette stratégie pourrait permettre aux femelles d’augmenter leur fitness en évitant les coûts associés aux interactions intersexuelles. Finalement, plutôt que de se cacher, les femelles peuvent aussi fuir et quitter l’environnement dans lequel elles sont. Cette stratégie pourrait être plus payante, notamment parce que les femelles ne peuvent pas aller chercher des proies sur l’eau tout en restant cachées. Chez l’espèce de patineur lotique Aquarius remigis, les femelles fuient lorsqu’elles sont confrontées à des mâles particulièrement agressifs et les femelles qui se font le plus agresser s’en vont plus souvent (Eldakar et coll. 2009a, 2010). On pourrait aussi se demander si la probabilité de fuite des femelles est en plus de cela affectée par des changements plus globaux de l’environnement social, comme des variations du sex-ratio ou de la densité.
La persistance comportementale des mâles a récemment reçu beaucoup d’attention, et plusieurs traits comportementaux sont déjà connus comme permettant aux mâles de gagner des copulations. Chez Gerris gracilicornis, les mâles tapent sur l’eau pour attirer des prédateurs, poussant les femelles à s’accoupler avec eux (Han & Jablonski 2010, Han & Brooks 2015). Il existe aussi des différences dans le niveau d’agressivité des mâles chez
Aquarius remigis (Sih & Watters 2005; Eldakar et coll. 2010; Chang & Sih 2013). Être
agressif permet aux mâles de gagner des copulations, même si ce gain est relatif à certaines conditions (Eldakar et coll. 2010; Chang & Sih 2013; Wey et coll. 2015a). L’agressivité des mâles semble constante dans le temps, mais les méthodes utilisées pour mettre en évidence cette constance ne sont pas celles utilisées habituellement (i.e., répétabilité d’un trait). Par ailleurs, personne ne s’est pour l’instant intéressé à voir si le niveau d’agressivité des mâles était maintenu dans différents contextes, comme dans différentes intensités de conflit sexuel ou de compétition entre mâles.
9
Contrairement aux avantages de la persistance, les coûts des mâles associés aux interactions intersexuelles ont reçu très peu d’attention. Des évidences montrent tout de même que les mâles ont des coûts à harceler les femelles (Devost & Turgeon 2016). Les mâles pourraient aussi avoir des coûts associés à la compétition entre mâles, notamment lorsqu’il y a un biais du sex-ratio vers les mâles. Dans ces conditions, les mâles ont moins d’opportunités de s’accoupler, et leur activité d’accouplement est effectivement plus faible en moyenne (Arnqvist 1992b; Han & Brooks 2013; Wey et coll. 2015a). Les moyens qu’ont les mâles pour éviter ces coûts restent par contre inexplorés.
Objectifs, hypothèses et prédictions
L’objectif général de ce projet de recherche était de mettre en lumière certains aspects du conflit sexuel qui avaient été généralement délaissés, en particulier dans les études sur les patineurs. Pour remplir cet objectif, il était nécessaire de faire deux expériences.
La première expérience visait à s’intéresser au comportement de fuite des femelles et des mâles. Pour les femelles, nous avons testé l’hypothèse que la fuite était un moyen pour elles d’échapper aux mauvaises conditions qui leur étaient imposées par le conflit sexuel. Nous avions prédit que la probabilité de fuite des femelles devrait augmenter avec l’intensité du conflit sexuel. A une échelle plus individuelle, nous avions aussi prédit que les femelles qui devraient fuir seraient celles qui s’étaient accouplées le plus souvent et qui s’étaient fait le plus harceler. Du côté des mâles, nous avons testé l’hypothèse que la fuite leur permettrait d’échapper aux conditions délétères imposées par la compétition entre mâles pour l’accès aux femelles. Nous avions prédit que la probabilité de fuite des mâles devrait augmenter avec l’intensité de la compétition entre mâles. Et par ailleurs, les mâles qui devraient s’en aller devraient être ceux qui s’accouplent peu souvent.
Dans la deuxième expérience, nous voulions voir si certains comportements de résistance et de persistance étaient des traits propres aux individus (i.e., traits de personnalité). Pour la résistance des femelles, nous nous sommes intéressés à la résistance physique exprimée par les femelles lors des luttes pré-copulatoires, et aussi à la tendance à se cacher à l’extérieur de l’eau. Du côté des mâles, nous nous sommes intéressés à leur agressivité et à leur persistance pendant les luttes pré-copulatoires. Pour chacun de ces
10
comportements, nous voulions voir si (1) il existait des différences entre les individus, et si ces différences étaient maintenues au cours du temps et dans différentes intensités de conflit sexuel (i.e., si ce sont des traits de personnalités) ; (2) ils étaient plastiques avec l’intensité du conflit sexuel et avec l’expérience d’accouplement récente des individus ; (3) les traits de résistance des femelles leur permettaient de limiter le harcèlement qu’elles subissaient ainsi que leur activité d’accouplement ; et (4) les traits de persistance des mâles leur permettaient de s’accoupler plus souvent.
11
CHAPITRE 2: Escaping sexual conflict and
male-male competition in Gerris buenoi
For consideration in Behavioral Ecology.
Pineaux, Maxime1 and Turgeon, Julie1
1Departement de biologie, Université Laval, Québec, Québec, G1V 0A6, Canada
Keywords: Sexual conflict, escape, dispersal, resistance, harassment, water strider, sex-ratio, male-male competition.
12
Abstract
Intersexual interactions linked to sexual conflict over mating rate can be highly detrimental to the fitness of female water strider. In nature, females often avoid struggling with males by escaping, but most studies on sexual conflict have focused on struggles. Males could also escape and follow females to avoid male-male competition for access to females, but it is generally accepted that they incur no or few costs associated with competition. We used Gerris buenoi to investigate whether females and males can escape from harsh conditions imposed by sexual conflict and male-male competition, respectively. Three sex ratios were used to experimentally manipulate the intensity of sexual conflict and male-male competition. Escape behavior was monitored daily for 15 days. Intersexual interactions between individuals were recorded to gain a more proximal understanding of factors influencing escape behavior. As expected, the probability that a female escapes increased with the proportion of males in the population, indicating that escape is an alternative means of resistance to avoid costly conditions imposed by sexual conflict. Contrary to predictions, frequently harassed females were not escaping more often, indicating that they were probably too weak to escape. The probability that a male escapes also increased with the proportion of males, and the males escaping had spent less time mating. Males may have escaped to avoid potential costs imposed by male-male competition for access to females. Our study confirms the importance of conducting experiments in realistic conditions allowing for a wide range of behaviors in both males and females.
13
Résumé
Un conflit sexuel sur le taux d’accouplement peut être extrêmement nuisible aux femelles chez certaines espèces de patineurs. Elles peuvent lutter physiquement avec les mâles pour éviter les accouplements superflus. Cependant, lutter n’est pas non plus sans coût. En nature, les femelles évitent souvent ces luttes pré-copulatoires en fuyant, mais cette potentielle forme de résistance a reçu peu d’attention. Les mâles eux aussi pourraient fuir pour diminuer la compétition entre mâles pour l’accès aux femelles, mais il est généralement accepté qu’ils ont très peu de coûts associé à la reproduction. Nous avons utilisé Gerris
buenoi pour déterminer si les femelles et les mâles peuvent utiliser la fuite pour éviter les
conditions délétères imposées respectivement par le conflit sexuel et la compétition entre mâles. Trois sex-ratios ont été utilisés pour manipuler expérimentalement l’intensité du conflit sexuel et de la compétition entre mâles. Le comportement de fuite a été suivi quotidiennement pendant 15 jours et les interactions intersexuelles ont aussi été observées pour comprendre de façon plus proximale ce qui entraînait la fuite. Comme attendu, la probabilité de fuite des femelles augmentait avec la proportion de mâles présents dans la population. Étonnamment, les femelles qui fuyaient étaient celles qui s’étaient fait le moins harceler et qui avaient passé le moins de temps en accouplement. Les femelles fréquemment confrontées aux mâles étaient probablement trop faibles pour être capables de fuir. Les mâles aussi fuyaient plus lorsque le sex-ratio était biaisé vers les mâles, et les fuyards étaient ceux qui avaient passé peu de temps en accouplement. La fuite pourrait être un moyen pour eux d’éviter de potentiels coûts liés à la compétition entre mâles et d’éventuellement augmenter leur opportunité de s’accoupler. Cette étude confirme l’importance de faire des observations dans des conditions réalistes et à la fois sur les mâles et les femelles.
14
Introduction
Sexual conflict arises when reproductive interactions affect male and female fitness in various ways (Parker 1979, 2006). In pre-copulatory sexual conflict, the interests of males and females diverge over the decision to mate (Chapman et al. 2003; Rowe & Day 2006). Males typically benefit from multiple matings and develop traits to harass females and better persist during mating attempts. These persistence traits can be detrimental to female reproduction (Arnqvist & Rowe 2005), decrease female fecundity (Rönn et al. 2006; Eldakar et al. 2011; Iserbyt et al. 2013; Makowicz & Schlupp 2013) and increase female mortality (Réale et al. 1996; Clutton-Brock & Langley 1997; Shine et al. 2001; Le Gaillard et al. 2005). Theoretically, sexual conflict over mating could ultimately drive populations to extinction in a “tragedy of the commons” scenario (Hardin 1968) because behaviors that are advantageous to individual males can deplete the common resource (females) they compete for in a way that is detrimental to the population as a whole (Kokko & Rankin 2006). However, antagonistic mechanisms can prevent this outcome (Rankin et al. 2011).
First, female resistance can evolve in response to male harm. Female resistance traits are morphological, but also physiological or behavioral (reviewed by Perry & Rowe 2015). Behavioral resistance is certainly more plastic than morphological resistance, potentially allowing females to respond rapidly to local conditions. This plasticity could be particularly advantageous when the economics of mating are context-dependent, for example with sex-ratio or density (Perry & Rowe 2015). On the one hand, females can resist directly by physically interacting with males (Arnqvist 1992a; Blyth & Gilburn 2011) or by showing aggressiveness (Forstmeier 2004; Stuart-Fox & Whiting 2005). On the other hand, females can avoid male harassment by resisting more indirectly. They can accept superfluous copulations (Rowe 1992; Harano 2015), hide (Krupa & Sih 1993; Bokides et al. 2012; Schultz & Fincke 2013; Bots et al. 2015), escape or disperse away (Shine et al. 2005; Darden & Croft 2008; Eldakar et al. 2009a, 2010).
Escaping from harsh environmental conditions can be beneficial despite the elevated costs likely associated with this behavior (Bonte et al. 2012). Females may escape from a given environment when male harassment is intense, for example because of male-biased sex-ratio or high density (Clutton-Brock & Parker 1995; Baguette et al. 1996; Shine et al.
15
2005; Darden & Croft 2008). Females also escape to avoid particularly harmful males (Eldakar et al. 2009a, 2010). However, females may be too weak to escape when confronted with high sexual conflict intensity (Le Gaillard et al. 2005). Resisting by means of escaping has not been studied as much as physical resistance, probably because most studies on sexual conflict have been conducted under laboratory conditions in limited space with few individuals, and over short periods of time (Arnqvist & Rowe 1995; Ortigosa & Rowe 2003). All aspects of female resistance may not be fully expressed under these conditions.
Second, a “tragedy of the commons” scenario can also be prevented if males incur costs from harassment or mating events (Rankin et al. 2011). However, very few studies have investigated and documented the negative effects of harassment and mating on male fitness (Green & Madjidian 2011; but see Makowicz & Schlupp 2013; Devost & Turgeon 2016). Competition among males over access to females may be costly for males. Male-male competition increases when sexual conflict over mating rate becomes more intense, for example under male-biased sex-ratio or high density (Kokko & Rankin 2006). Fights between males requires energy and sometimes result in physical injuries (Gergits & Jaeger 1990; Qvarnström 1997; Savage et al. 2005). Strong male-male competition also decreases mating opportunities for a majority of males.
It is expected that less competitive males would escape from harsh conditions and eventually enjoy higher mating and reproductive success in a more favorable environment. A few studies have indeed shown that males escape more from male-biased groups, where male-male competition is more intense (Lawrence 1987; Croft et al. 2003; Trochet et al. 2013). However, very few studies have also investigated if the escaping males were the most or the less competitive (Lawrence 1987; see also Shine et al. 2006; Okada et al. 2007).
Water striders (Gerridae) provide some of the best examples of sexual conflict over mating rate (Arnqvist 1992a; Rowe et al. 1994; Arnqvist & Rowe 2005; Perry & Rowe 2015). In many species, males frequently harass females to gain matings, while females strongly resist unnecessary and costly matings (e.g. increased predation risk and energy expenditure; Arnqvist 1989; Fairbairn 1993; Watson et al. 1998). Water strider females resist by various means. They engage in vigorous pre-copulatory struggles to dislodge males (Rowe 1992). However, resisting males is more costly than mating when harassment is intense so that
16
females may accept to mate multiply for convenience (Rowe 1992). Second, but less frequently investigated, females can resist by hiding off the water to reduce male harassment (Krupa et al. 1990; Krupa & Sih 1993; Rowe et al. 1994; Pineaux & Turgeon, chapitre 3). Finally, females may escape from particularly aggressive males by moving to habitat patches where less aggressive males aggregate (Eldakar et al. 2009a, 2010). Male water striders supposedly incur very little costs from mating, whether caused by sexual conflict or male-male competition (Arnqvist 1997; Watson et al. 1998). However, harassment can reduce male-male reproductive success in G. buenoi, indicating that it may be a costly behavior (Devost & Turgeon 2016). Although male-male competition has received very little attention, empirical evidence shows that males have less opportunities to mate when sex-ratio is male-biased (Arnqvist 1992b; Han & Brooks 2013; Wey et al. 2015a).
Using the water strider species G. buenoi in a realistic experimental setting, we simultaneously tested the hypothesis that females tend to escape from harsh conditions imposed by sexual conflict, while male escape from conditions increasing male-male competition. For females, we therefore expect that the probability of escape will increase with sexual conflict intensity. We also predict that females more frequently harassed and mated will be more likely to escape. For males, we expect that the probability of escape will increase with male-male competition intensity. We also predict that less competitive males gaining fewer matings will be more likely to escape. Our results show that females escape from high sexual conflict intensity instead of opting for other presumably costlier means of resistance. However, frequently mated and heavily harassed females were not escaping more often. Males escaped more from male-biased groups, and males with low mating activity escaped more, possibly to decrease male-male competition.
Materials and methods
Sample collection and experimental setup
Overwintered G. buenoi adults were collected from a pond in a bog near Quebec City (Sainte-Catherine-de-la-Jacques-Cartier; 46.524N, 71.342W) on the 9th of May 2014, just
after the pond thawed. The experiment was conducted in this bog, near the pond banks. There were three sex-ratio treatments (males:females): female-biased (1:3), balanced (1:1), and
17
male-biased (3:1). These conditions represent increasing intensities of sexual conflict, and this range of sex-ratios is observed in nature. Five replicates were performed per sex-ratio.
We formed groups of 20 individuals and we placed each group in a large wading pool (diameter: 1.5 m; surface: 1.77 m²; water depth: 15 cm) near the pond bank. Body length was measured and insects were individually marked on the pronotum with two dots of enamel paint (Paint Pen, ©Metron Marker). Each pool was separated in two sections by anchoring a wall (100 x 30 cm) such that the larger section covered 70% of the pool. Water striders could escape from the large section through a small opening in the wall (3 cm). Escapees could move back to the large section but few individuals did so (unpublished data). We covered the pools with nets to prevent overwintered winged adults from flying away. A few individuals managed to leave, but there were always at least 18 individuals remaining in each pool. Density and sex-ratio were kept constant by replacing missing individuals. We put a piece of peat on a floating platform on each side of the wall to provide resting sites, and we provided a rain shelter (upside down Styrofoam container) in the main section. Three frozen crickets were added at the end of the day in each pool as food.
Behavioral observations
Escape behavior was monitored every morning for 15 consecutive days. Before 9:00, the location of each individual in the pool was recorded. As all individuals were originally located in the main section of the pool, water striders were classified as having escaped if they were observed on the smaller section of the pool. All escaped individuals were then put back in the main section of the pool after this daily check. Thus, escape behavior was coded as a binary variable (escape/no escape), and each individual had one binary observation per day.
To gain a more proximal understanding of what drives escape behavior, we also performed 5-min continuous observation sessions to record pre-copulatory interactions and mating status of each individual located in the main section of the pool. Our goal was to investigate how escape behavior recorded at any given day could be explained by intersexual interactions during the previous day. Observations were performed twice to five times per day depending on weather conditions. Data were collected between 10:30 and 15:30 and observations were separated by at least one hour. Only the main section of the pool was
18
observed; the behavior of escapees was not observed. Recording mating status allowed us to estimate the daily mating activity of each individual as the proportion of sessions during which an individual was observed mating on a particular day.
Pre-copulatory interactions were recorded (see Table 2.1 for description). Touch (T), jump (J), short pre-copulatory struggle (SS), and long pre-copulatory struggle (LS) were considered to represent increasing levels of harassment intensity (Eldakar et al. 2010; Chang & Sih 2013). Based on these behaviors, we calculated a daily harassment score for each individual. This score measures the harassment performed by males or the harassment endured by females. We used two estimation methods. First, we designed a formula similar to that of Eldakar et al. (2009b) (see Eq. 1) in which the number of weighted aggressive interactions was divided by the number of times the individual was observed on that day. Endured/performed harassment score =𝐿𝑆(4)+𝑆𝑆 (3)+𝐽 (2)+𝑇 (1)𝑁𝑜𝑏𝑠𝑒𝑟𝑣𝑎𝑡𝑖𝑜𝑛𝑠 (1)
Admittedly, the estimates of energy expenditure for each kind of pre-copulatory interactions are somewhat subjective. Consequently, we also performed a principal component analysis (PCA), treating sexes separately. For males and females, PC axis 1 reflected global harassment (see Supplementary Material Table 2.S1). Since weighted and PCA scores were highly correlated (Pearson correlation coefficient: males, r = -0.91, P < 0.001; females, r = -0.95, P < 0.001), we only used the former for statistical analyses.
Statistical analyses
We used generalized linear mixed models (GLMM) to study the impact of sex-ratio, mating activity, and harassment on escape behavior. We constructed one set of GLMM per sex, using data from 169 females and 171 males. Number of days of observation per individual ranged from 1 to 15 (mean: 14 days). For each sex, the best model was determined using model selection based on corrected Akaike Information Criteria (AICc) (Burham & Anderson 2002; Mazerolle 2006). As indicated above, we used mating activity and harassment score on a given day as potentially explanatory variable for the escape behavior recorded the next morning.
19
Models for males and females were very similar to each other, except that harassment score reflects performed or endured harassment for males and females, respectively. The full model was as follows:
Escape behavior ~ sex-ratio + mating activity + (performed or endured) harassment score + body length + first-order interactions + pool [individual identity] (random) + day of observation (random) + error
Individual identity was nested within pool identity and included as a random effect to prevent pseudoreplication. Likewise, the day of observation was considered a random effect to prevent temporal pseudoreplication. A binomial distribution and a logit link were used in the models.
All statistical analyses were performed with R (R Development Core Team 2008). The LME4 R package (Bates et al. 2008) was used to run all models. For each sex, the model with the highest Akaike weight was selected as the best model using the AICcmodavg R package (Mazerolle 2015).
Results
The sex-ratio treatment was effective in increasing female endured harassment and decreasing mean male mating activity. Mean endured harassment score per female per day was higher in the male-biased sex-ratio (mean ± SE: 2.19 ± 0.21) than in balanced (1.06 ± 0.09) and female-biased sex-ratios (0.53 ± 0.03; Supplementary Material Fig. 2.S2). This confirms that the male-biased treatment resulted in higher harassment intensity. Additionally, males had a lower mating activity in the male-biased sex-ratio (0.19 ± 0.30) than in the two other sex-ratios (female-biased: 0.45 ± 0.35; balanced, 0.44 ± 0.35; Supplementary Material Fig. 2.S3) indicating that male mating opportunities decreased with the proportion of males in the pool.
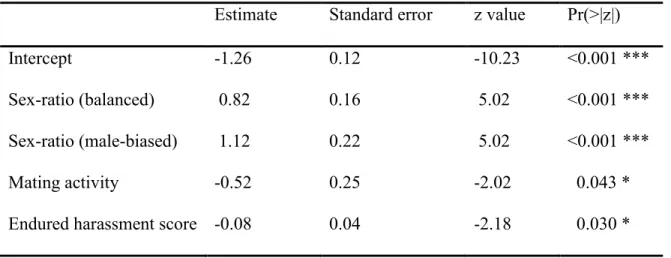
For females, the best model indicated that escape behavior was explained by sex ratio, mating activity and endured harassment (Table 2.2A and see Table 2.S2A in the Supplementary Materials for all models). Females escaped more in male-biased and balanced sex-ratios than in the female-biased sex-ratio (Table 2.2A, Fig. 2.1A). Surprisingly, the
20
probability that a female escape significantly decreased when endured harassment score and mating activity was high (Table 2.2A). These results indicate that less frequently harassed females and females that had a lower mating activity tended to escape more this day.
For males, the best model indicated that escape behavior was explained by sex ratio and mating activity, but not by performed harassment (Table 2.2B and see Table 2.S2B in the Supplementary Materials for all models). Males also escaped more in the male-biased sex-ratio than in the female-biased sex-ratio (Table 2.2B, Fig. 2.1B), suggesting that the probability that a male escape increased when the opportunity for mating decreases. This trend is also supported by the significant negative relationship between escape tendency and mating activity (Table 2.2B). Thus, males that had a low mating activity escaped more during this day.
Discussion
This study provides an overview of the escape behavior in the water strider species
G. buenoi. We conducted an experiment in semi-natural populations, we let individuals
interact freely over several days, and we recorded both females’ and males’ behavior. Based on solid past knowledge, we used different sex-ratios to experimentally manipulate sexual conflict and male-male competition intensities (Arnqvist 1992a, b; Rowe 1992; Han & Brooks 2013; Wey et al. 2015a). The sex-ratio treatment was effective as indicated by the higher mean harassment score endured by females and the lower mean male mating activity in the male-biased sex-ratio than in the female-biased sex-ratio. Very few studies have simultaneously examined escape behavior of both sexes as a consequence of sexual conflict and male-male competition (but see Le Gaillard et al. 2005; Shine et al. 2006; Trochet et al. 2013). All these features collectively allowed us to make meaningful observations.
Our primary objective was to test the hypothesis that female should escape more frequently when sexual conflict is intense. As expected, females escaped more in the male-biased sex-ratio than in the female-male-biased sex-ratio. Our results also indicated that females endured a higher harassment rate in the male-biased ratio than in the female-biased sex-ratio. Male harassment may reduce female fitness in many ways, ranging from physical harm to inhibition of foraging efficiency (Réale et al. 1996; Rönn et al. 2006; Eldakar et al. 2011;
21
Devost & Turgeon 2016). This indicates that females certainly incurred a much higher cost in the male-biased sex-ratio than in the female-biased sex-ratio. The above observations collectively support that females escaped to avoid harsh conditions imposed by sexual conflict. Escaping is certainly less costly for females than struggling or accepting superfluous copulations. Yet, this form of resistance is seldom considered in studies on sexual conflict. This is certainly due to the necessity of conducting an experiment in semi-natural conditions providing enough space to allow free individual movement for several days. Our experiment met those criteria, and two weeks of observations provided conclusive results.
Females incurring higher costs from harassment due to sexual conflict were not among those escaping during the rest of the day. This is surprising since it could be expected that heavily harassed females should escape rapidly to avoid future costly events (Eldakar et al. 2009a, 2010). An increase in female harm caused by males has also been associated with a trend for lower female escape rate in the common lizard Lacerta vivipara (Le Gaillard et al. 2005). However, contrary to our results, this pattern was observed at the population level and over a much longer period of time (several months). Our results therefore suggest that females were in poor condition shortly after mating and harassment events. Direct, physical pre-copulatory resistance and mating events are highly energetically demanding for females (Watson et al. 1998). Male harassment also decreases female feeding efficiency, which could in turn decrease body condition (Rowe et al. 1994). Instead of escaping, severely harmed females may have used a less costly form of resistance like hiding off the water. This behavior has already been proposed as a means to lower harassment costs for females (Krupa et al. 1990; Krupa & Sih 1993; Wey et al. 2015b). Most importantly, hiding could prevent these temporarily vulnerable females from re-mating too rapidly. Unfortunately, we did not record which females were hiding during the experiment. It is known that females hide more often when the sex-ratio is male-biased (Krupa & Sih 1993; Wey et al. 2015b), so it would be interesting to investigate whether females hide more often soon after mating.
Males also escaped more when the intensity of the sexual conflict was increased because of male-biased sex-ratio. This was revealed by the higher probability of escaping in the male-biased sex-ratio compared to the female-biased sex-ratio. For males, opportunities to mate decrease when the proportion of males in a population increases, as indicated by the
22
lower mean mating activity in the male biased ratio compared to the female biased sex-ratio. Our results are consistent with the idea that a male-biased sex-ratio impose competition costs on males. Male-male competition has rarely been investigated in systems where there is a sexual conflict over mating rate, and this is especially the case for water striders (but see Han & Brooks 2013; Wey et al. 2015a). This is probably due to the lack of readily observable interference competition such as male-male combats or territorial defense behaviors in most water strider species (but see Hayashi 1985). Yet, competitive mate search results in intense male-male scramble competition in water striders, particularly when sex-ratio is male-biased. We did not estimate the effects resulting from this behavior, but escaping could be a way for males to increase their mating probability. In milkweed beetles (Tetraopes tetraophthalmus), male may leave a male-biased patch for a female-biased patch (Lawrence 1987), thus likely securing better mating opportunities. Here, individual-based observations lead us to infer that escapees were probably also the less competitive males. Evidence for this pattern is scarce (Lawrence 1987; Shine et al. 2006; Okada et al. 2007) and it would be interesting to explore whether these less competitive males truly increase their mating/reproductive success through escaping.
We obtained unsuspected results regarding the balanced sex-ratio treatment in our experiment. We predicted that the balanced sex-ratio treatment should represent an intermediate intensity of sexual conflict and male-male competition. However, females did not escape more often in male-biased than in balanced sex-ratios, while the male tendency to escape was similar in balanced and female-biased ratios. It seems that the balanced sex-ratio treatment corresponded to high sexual conflict intensity and low male-male competition intensity. There was an intermediate level of harassment in the balanced sex-ratio, but mating activity was as high as in the male-biased sex-ratio. This may be due to the density used in this experiment. Indeed, sexual conflict intensity also increases with density (Arnqvist 1992a). Here, the density was low compared to other studies (Rowe 1992; Arnqvist & Rowe 1995; Devost & Turgeon 2016), but it remains higher than in natural populations. A much more complex and challenging experimental design would have been needed to disentangle the effects of density and sex-ratio on sexual conflict and male-male competition on escape behavior.
23
Perspectives
This experiment allowed for investigating the short-term response of males and females within controlled social contexts. Other studies have used experimental settings of similar spatial extent to study dispersal (Eldakar et al. 2009a, 2010). Here, we chose to describe the observed behavior as escape. Indeed, all such experiments are likely and necessarily simplifications of dispersal in truly natural populations. Dispersal is broadly defined as “an active or passive attempt to move from a natal or breeding site to another
breeding site” (Clobert et al. 2009). The definition of a “site” depends on the species and the
habitat structure, but for G. buenoi a pond might be considered as a unique site. However, given that overwintered G. buenoi can fly in the spring, it would be interesting to explore if individuals fly to disperse between ponds as a means for females to avoid sexual conflict and for male to lessen male-male competition. Using winged individuals would be a way to test more directly a better proxy of dispersal effect. In theory, dispersal can promote or inhibit runaway evolution between female resistance and male harassment via kin selection (Rankin 2011). The outcome depends on whether dispersal is male- or biased. If it is female-biased, as in this study, dispersal should inhibit the evolution of male harm because remaining males would compete with kin for mates (Rankin 2011; Faria et al. 2015).
Our experiment tested for the effect of the social environment on individual behavior and each individual was tested under a single sex-ratio. We do not know whether any one individual would have expressed the same behavior in a different context. Resistance could be a personality trait if the probability of escaping, struggling or hiding remains constant over time and across contexts such as sexual conflict intensity (Sih et al. 2004; Réale et al. 2007; Briffa & Weiss 2010; Dingemanse et al. 2010). The relative influence of intrinsic individual features and the social environment on female resistance remain to be ascertained.
24
Tables
Table 2.1 Ethogram of observed pre-copulatory behaviours in Gerris buenoi.
Behaviour Definition
Touch (T) Physical contact between a male and a female due to the voluntary movement of one individual towards another, but without any attempt to mount.
Jump (J) Physical contact where a male mounts a female but fails at grasping her with his forelegs.
Short pre-copulatory struggle (SS)
Physical contact lasting less than 3 s where a male mounts a female and succeeds at grasping her with his forelegs while attempting to copulate. The female often resists with rolls and somersaults. Long pre-copulatory
struggle (LS)