HAL Id: hal-02602035
https://hal.inrae.fr/hal-02602035
Submitted on 16 May 2020
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Suivi hydrobiologique de la Durance : Étude DREAM
-Dynamique de REcolonisation des Algues et des
Macroinvertébrés suite aux lâchers d’eau - Résultats
macroinvertébrés - Année 2014
A. Morel, G. Archambaud-Suard
To cite this version:
A. Morel, G. Archambaud-Suard. Suivi hydrobiologique de la Durance : Étude DREAM - Dynamique de REcolonisation des Algues et des Macroinvertébrés suite aux lâchers d’eau - Résultats macroin-vertébrés - Année 2014. irstea. 2015, pp.68. �hal-02602035�
Etude réalisée avec le concours financier de l’Agence de l’Eau RM&C
UNITE DE RECHERCHE HYDROBIOLOGIE
AOUT 2015
Adrien MOREL
Gaït ARCHAMBAUD-SUARD
Irstea
Centre d'Aix en Provence
3275 route de Cézanne - CS 40061
13182 AIX EN PROVENCE
Suivi hydrobiologique de la Durance
Etude DREAM
Dynamique de REcolonisation des
Algues et des Macroinvertébrés
suite aux lâchers d’eau
Résultats macroinvertébrés - Année 2014
Les auteurs remercient Bernard DUMONT pour ses conseils et son aimable relecture.
Cette étude a fait l’objet du stage de master professionnel d’Alexandre PUJOL soutenu en septembre 2014 à l’université d’Aix-Marseille. Les résultats de l’année 2014 ont également fait l’objet d’une présentation orale au colloque ISRivers en juin 2015 à Lyon.
1
Suivi hydrobiologique de la Durance
Etude DREAM :
Dynamique de REcolonisation des
Algues et des Macroinverte bre s suite
aux la chers d’eau
Re sultats macroinverte bre s de l’anne e 2014
Septembre 2015
Sommaire
1 Contexte de l’étude 4
1.1 Actions de restaurations hydrologiques en Durance 4
1.1.1 « Régime réservé » : modulation saisonnière des débits réservés 4
1.1.2 Lâchers de décolmatage 5
1.2 Suivis hydrobiologiques des lâchers et étude de la dynamique de recolonisation 6
1.3 Contexte hydro-climatique 7
1.3.1 Evolution temporelle des débits lors des sept premiers mois de 2014 7
1.3.2 Evolution des températures lors des sept premiers mois de 2014 8
1.4 Orientations de l’étude DREAM pour l’année 2014 9
2 Matériels et méthodes 10
2.1 Stations d’étude 10
2.2 Campagnes d’échantillonnage 12
2.3 Protocole de prélèvement 12
2.3.1 Zones de prélèvement 12
2.3.2 Méthode de prélèvement et description des variables d’habitat 12
2.3.3 Tri et identification des macroinvertébrés 14
3 Résultats 16
3.1 Approches globales 16
3.1.1 Données hydrologiques et thermiques 16
3.1.1.1 Historique hydrologique des campagnes 16
3.1.1.2 Historique thermique des campagnes de prélèvement 17
3.1.2 Données mésologiques 19
3.1.2.1 Variabilité des conditions d’habitat entre les ambiances d’écoulement 19
3.1.2.2 Degrés de colmatage des substrats 21
3.1.2.2.1 Bilan par ambiances d’écoulement 21
3.1.2.2.2 Évolution temporelle du colmatage par ambiances d’écoulement 23
3.1.3 Bilan faunistique global et par station d’étude 24
3.1.3.1 Bilans par station 24
3.1.3.2 Bilan des principaux groupes faunistiques 25
3.1.4 Structures faunistiques observées dans les trois ambiances d’écoulement 28
3.1.4.1 Richesse et composition faunistique des ambiances d’écoulement 28
3.1.4.2 Effectifs et structures des peuplements des ambiances d’écoulement 31
3.2 Dynamique temporelle des peuplements 33
3.2.1 Evolution des structures des peuplements entre les campagnes 33
3.2.1.1 Richesses taxonomiques par échantillon 33
3.2.1.2 Effectifs par échantillon 34
3.2.1.3 Richesses taxonomiques et effectifs relatifs par campagne et par station 34
3.2.1.4 Indices de similarités structurelles entre campagne - station 36
3.2.1.5 Relation entre effectifs et richesses des échantillons 37
3.2.2 Contribution des ambiances d’écoulement dans les processus de recolonisation 40
3.2.3 Dynamique de croissance des taxons EPT 45
3.2.3.1 Structuration globale en classes de tailles 45
3.2.3.2 Hypothèse de l’électivité taxonomique dans un site 46
3.2.4 Réponses fonctionnelles à l’évolution du degré de colmatage des substrats 49
3.2.4.1 Démarche analytique 49
3.2.4.2 Variations des abondances relatives des interactions fonctionnelles crées en lien
avec le colmatage 52
1
Table des figures
Figure 1 : Evolution des débits au pas de temps horaire (tracé bleu) et des débits moyens mensuels (rectangles bleus) de la
Durance en aval du barrage de l’Escale entre le 1er janvier et le 31 juillet 2014. L’axe des ordonnées est représenté avec une échelle logarithmique. Les débits maximaux atteints lors des épisodes de crues sont indiqués au-dessus de la courbe. La boite à moustache dans la partie droite concerne les données de la période considérée. Source : Edf. ... 7
Figure 2 : Evolutions des températures au pas de temps horaire (tracé rouge) et des moyennes journalières (tracé noir) et
mensuelles (rectangles rouges) des eaux de la Durance environ 900 m en aval du barrage de l’Escale (longitude : 6,0076905 ; latitude : 44,077718 en degrés décimaux) entre le 1er janvier et le 31 juillet 2014. La boite à moustache dans la partie de droite concerne les données de la période considérée. Source : SMAVD. ... 8
Figure 3 : Représentation simplifiée du protocole d‘estimation du colmatage du substrat. La première colonne correspond
aux codes de colmatage (chiffres en rouge), la seconde colonne correspond aux classes exprimées en pourcentage et la dernière colonne schématise le substrat en coupe latérale avec l’épaisseur des dépôts de surface, la cohésion des éléments minéraux et l’intensité de la bouffée turbide engendrée lors de la déstructuration du substrat. ... 14
Figure 4 : Distribution des débits horaires des 30 jours précédant chaque campagne de prélèvement. L’axe des ordonnées
est représenté avec une échelle logarithmique. ... 16
Figure 5 : Distribution des températures horaires des 30 jours précédant chaque campagne de prélèvement. ... 18 Figure 6 : Distributions des valeurs de vitesses maximales dans la colonne d’eau en fonction des ambiances d’écoulements
pour les deux stations d’étude. Les trois zones grisées correspondent aux valeurs ciblées de vitesse de surface pour chaque ambiance (A1 : gris foncé entre 30 et 50 cm/s ; A2 : gris intermédiaire entre 70 et 90 cm/s ; A3 : gris clair pour des vitesses > 100 cm/s) ... 19
Figure 7 : Distribution des granulométries moyennes par ambiance d’écoulement pour les deux stations d’étude. Les codes
granulométriques utilisés sont : 1 –vase (< 4 µm), 2 –limon (> 4 µm), 3 –sable fin (> 62 µm), 4 –sable grossier (> 0,5 mm), 5 – gravier fin (> 2 mm), 6 –gravier grossier (> 8 mm), 7 –cailloux fin (> 16 mm), 8 –cailloux grossier (> 32 mm), 9 –pierre fine (> 64 mm), 10 –pierre grossière (> 128 mm), 11 –bloc fin et moyen (> 256 mm), 12 –bloc grossier (>1024 mm), 13 –bloc très grossier (> 2048 mm), 14 –végétal. ... 20
Figure 8 : Distributions des vitesses de courant mesurées à 3 cm du fond par ambiance d’écoulement pour les deux stations
d’étude. ... 21
Figure 9 : Fréquence d’occurrence des trois classes de colmatage observées en 2014 par ambiance d’écoulement pour les
deux stations d’étude. Pour rappel : classe 1 – colmatage estimé entre 0 et 25 % ; classe 2 – entre 25 et 50 % ; classe 3 – entre 50 et 75 %. ... 22
Figure 10 : Variation de l’indice moyen de colmatage des ambiances d’écoulement par campagne et indice moyen par
campagne pour les deux stations d’étude. ... 23
Figure 11 : Richesses taxonomiques (figure de gauche) et effectifs bruts (figure de droite) des peuplements agrégés par
station d’étude et décomposition par groupes faunistiques. ... 27
Figure 12 : Richesses taxonomiques (figure de gauche) et effectifs bruts (figure de droite) des peuplements agrégés par
ambiance d’écoulement et par station d’étude et décomposition par groupes faunistiques. ... 28
Figure 13 : Distributions des valeurs de richesses taxonomiques par échantillons en fonction des ambiances d’écoulement
pour les deux stations d’étude. ... 29
Figure 14 : Distributions des effectifs par échantillons en fonction des ambiances d’écoulement pour les deux stations
d’étude. Les axes des ordonnées sont représentés avec une échelle logarithmique. ... 31
Figure 15 : Distribution des richesses taxonomiques de chaque échantillon par campagne de prélèvement pour les deux
stations d’étude. Pour rappel : C1 – 27/01/2014 ; C2 – 17/03/2014 ; C3 – 16/04/2014 ; C4 – 20/05/2014. ... 33
Figure 16 : Distribution des effectifs de chaque échantillon par campagne de prélèvement pour les deux stations d’étude.
Pour rappel : C1 – 27/01/2014 ; C2 – 17/03/2014 ; C3 – 16/04/2014 ; C4 – 20/05/2014. Les axes des ordonnées sont représentés avec une échelle logarithmique. ... 34
Figure 17 : Richesses (figure de gauche) et effectifs relatifs (figure de droite) des peuplements agrégés par campagne de
prélèvement et par station d’étude et décomposition par groupes faunistiques... 35
Figure 18 : Représentation du logarithme des effectifs en fonction des richesses taxonomiques de chaque échantillon pour
les deux stations d’étude, et droites de régression linéaire au niveau stationnel (droites noires continues) et au sein de chaque ambiance d’écoulement (droites colorées en pointillés)... 38
Figure 19 : Station D0BIS : distributions des valeurs d’indice de similarité de Whittaker (à gauche) et de Jaccard (à droite)
2
boites à moustaches correspondent aux groupes établis à partir des résultats des tests de comparaison de moyennes de
Wilcoxon (« ns » : non significatif). ... 41
Figure 20 : Station D1 : distributions des valeurs d’indice de similarité de Whittaker (à gauche) et de Jaccard (à droite) entre les prélèvements d’une ambiance lors d’une campagne t et tous ceux de la campagne t+1. Les lettres au-dessus des boites à moustaches correspondent aux groupes établis à partir des résultats des tests de comparaison de moyennes de Wilcoxon (« ns » : non significatif). ... 42
Figure 21 : Distributions par site des effectifs bruts (figures en haut ; axes des ordonnées en logarithme) et des effectifs relatifs (figures en bas) des classes de taille (exprimées en mm) des individus PET en excluant le genre Hydropsyche (figures de gauche) et de tous les PET (figures de droite). ... 45
Figure 22 : Exemples de la distribution des effectifs bruts (figures de gauche) et relatifs (figures de droite) par classes de tailles (exprimées en mm) de trois espèces d’éphéméroptères pour les deux stations d’étude. ... 46
Figure 23 : Distribution des effectifs bruts (figures de gauche) et relatifs (figures de droite) des classes de taille (exprimées en mm) d’Oligoneuriella rhenana entre les campagnes de prélèvement et pour les deux stations d’étude. ... 47
Figure 24 : Schéma conceptuel utilisé pour la création de quatre interactions de traits fonctionnels sélectionnés vis-à-vis d’une problématique de colmatage des substrats par les sédiments fins. ... 49
Figure 25 : Station D0BIS : évolution par campagne des abondances relatives des quatre interactions fonctionnelles. Les lettres au-dessus des boites à moustaches correspondent aux groupes établis à partir des résultats des tests de comparaison de moyennes de Wilcoxon (« ns » : non significatif). ... 52
Figure 26 : Station D1 : évolution par campagne des abondances relatives des quatre interactions fonctionnelles. Les lettres au-dessus des boites à moustaches correspondent aux groupes établis à partir des résultats des tests de comparaison de moyennes de Wilcoxon. ... 53
Figure 27 : Station D1 : évolution des abondances relatives des interactions fonctionnelles par ambiances et par campagne. ... 54
Table des tableaux et cartes
Carte 1 : Bassin de la Durance. ... 4Tableau 1 : Modalités prévisionnelles des lâchers d'eau dans le tronçon de l'Escale-Cadarache. ... 5
Carte 2 : Localisation des stations d’étude D0BIS et D1 dans le tronçon de l’Escale-Cadarache. ... 10
Photo 1 : Stations D0BIS à gauche et D1 à droite en 2014 (photos : Irstea). ... 11
Tableau 2 : Dates des campagnes de prélèvements réalisées en 2014 et intervalles de temps entre deux campagnes consécutives. ... 12
Tableau 3 : Bilans des occurrences de dépassements de valeurs seuils par les des débits horaires durant des périodes de 30 jours précédant chacune des campagnes (744 valeurs). ... 17
Tableau 4 : Antériorité en nombre de jours de l’épisode de crue du 19 janvier 2014 aux campagnes de prélèvements. ... 17
Tableau 5 : Bilan global et par station des effectifs bruts, des densités, des richesses taxonomiques et du nombre de taxons représenté par plus d’un individu. ... 24
Tableau 6 : Bilans global et par station des effectifs, des richesses taxonomiques et du nombre de taxons représenté par plus d’un individu pour les principaux groupes faunistiques. Les effectifs et les richesses sont exprimés en valeurs brutes et en valeurs relatives (Tableau 5). ... 25
Tableau 7 : Indices de Jaccard (partie inférieure gauche), richesses taxonomiques (cases grisées) et nombre de taxons communs (partie supérieure droite) entre les entités ambiance - station. Les valeurs maximales d’indice de chaque ambiance – station sont figurées en bleu et en gras. ... 30
Tableau 8 : Indices de Whittaker-Fairbanks entre chaque couple d’ambiance – station. Les valeurs maximales d’indice sont figurées en bleu et en gras. ... 32
Tableau 9 : Indices de Whittaker-Fairbanks entre chaque couple de campagne – station. Les valeurs d’indice de deux campagnes consécutives sont indiquées en gras et en bleu. Les valeurs grisées correspondent au couplage des entités de stations différentes. Les valeurs soulignées et en gras correspondent aux maximums observés pour chaque entité. ... 36
Tableau 10 : Indices de Jaccard entre chaque couple de campagne – station. Les valeurs d’indice de deux campagnes consécutives sont indiquées en gras et en bleu. Les valeurs grisées correspondent au couplage des entités de stations différentes. Les valeurs soulignées et en gras correspondent aux maximums observés pour chaque entité. ... 36
Tableau 11 : Caractérisation des traits et des quatre interactions pour chaque taxon. Les taxons surlignés en orange se rencontrent simultanément dans les deux premières modalités, les taxons surlignés en bleu dans les deux dernières. ... 51
4
1 Contexte de l’étude
La Durance est l’un des principaux affluents du Rhône dans sa partie aval. Depuis le début du XXème
siècle, cette rivière en tresse a été fortement aménagée pour des besoins de production hydroélectrique et d’irrigation agricole principalement. Depuis le barrage de Serre-Ponçon jusqu’à sa confluence avec le Rhône, sept retenues au fil de l’eau constituent l’essentiel de la production hydroélectrique en Durance. Au total, ce sont plus de 200 km du linéaire du cours moyen et aval de la Durance qui sont soumis à un régime hydrologique réservé.
Carte 1 : Bassin de la Durance.
1.1 Actions de restaurations hydrologiques en Durance
La gestion des ouvrages hydroélectriques en France est soumise à de nombreuses obligations réglementaires. L’article L214-18 du Code de l’Environnement, modifié par la loi sur l’eau et les milieux aquatiques du 30 décembre 2006, définit les obligations en matière de débits réservés
applicables aux aménagements hydroélectriques au plus tard au 1er janvier 2014.
1.1.1 « Régime réservé » : modulation saisonnière des débits réservés
EDF, gestionnaire de la chaine hydroélectrique en moyenne Durance, a proposé un relèvement du débit réservé réglementaire dans le cadre de cette législation. En complément de ce relèvement, une modulation saisonnière des débits réservés a été instaurée. Celle-ci consiste à laisser transiter
1/14ème du module en été et 1/20ème en période hivernale dans les tronçons court-circuités. Ce
5 2007 jusqu’à la fin de l’année 2010 dans le cadre du renouvellement de la concession de Sainte-Tulle, puis son application a été étendue à l’ensemble des tronçons court-circuités en 2014.
1.1.2 Lâchers de décolmatage
Les résultats de l’expérimentation Sainte-Tulle ont également permis de mettre en évidence l’influence négative sur le potentiel d’habitat du colmatage des substrats minéraux durant des périodes d’absence de crues morphogènes. Ainsi, une mesure de gestion supplémentaire a été mise en application sur l’ensemble des tronçons de moyenne Durance depuis l’année 2014. Il s’agit de réaliser des lâchers d’eau (ou « pulses hydrauliques ») afin de décolmater les substrats des habitats lotiques. Le principal objectif est de nettoyer ces substrats avant la période de ponte pour favoriser la fraie d’espèces piscicoles cibles utilisant les supports minéraux pour leur ponte. Cette mesure devrait donc permettre d’améliorer le recrutement de ces espèces. Des retombées positives secondaires sur d’autres compartiments biologiques aquatiques ainsi que sur des facteurs abiotiques sont attendues : favoriser certains taxons d’invertébrés aquatiques par l’intermédiaire d’une augmentation de la qualité d’accueil des substrats ; améliorer les échanges entre la nappe d’accompagnement et le lit de la Durance.
Les modalités des lâchers d’eau ont été ajustées aux particularités de chaque tronçon soumis au régime réservé. Les principales modalités retenues pour le tronçon de l’Escale-Cadarache sont présentées à titre d’exemple dans le Tableau 1.
Tableau 1 : Modalités prévisionnelles des lâchers d'eau dans le tronçon de l'Escale-Cadarache.
Période Augmentation Débit
maximal Diminution
Volume
maximal Durée totale
Entre début février et mi-mars Par palier Durant 1 h30 70 m 3 /s Progressive Durant 3h00 3,8 hm 3 10h00
L’intensité du lâcher doit permettre une remise en suspension des sédiments déposés sans pour autant engendrer une mobilisation des éléments minéraux de granulométries importantes, ni être pénalisante pour la sécurité des tiers. La définition de l’hydrogramme des lâchers (intensité, durée totale, modalités des paliers de montée et de baisse, …) a été établie à partir de retours d’expériences effectuées dans le tronçon de la Saulce à St-Lazare en l’adaptant en fonction des morphologies plus larges et plus en tresses dans les tronçons en aval.
La période de réalisation est définie selon les espèces de poissons ciblées par tronçon. Pour le tronçon de l’Escale-Cadarache, les espèces ciblées étaient à l’origine les cyprinidés d’eaux vives, mais depuis l’année 2015, la présence d’Apron (Zingel asper) est prise en considération. La période de réalisation du lâcher est donc prévue dans ce tronçon entre le début du mois de février et le 15 mars. La réalisation d’un lâcher est toutefois conditionnée à l’absence d’épisode hydrologique de même niveau que le lâcher dans les mois précédents et en l’absence de prévision de crue dans les 15 jours suivant la date prévue du lâcher.
6
1.2 Suivis hydrobiologiques des lâchers et étude de la dynamique de
recolonisation
Afin de quantifier l’influence de ces lâchers sur les biocénoses aquatiques, de nombreux compartiments biologiques ainsi que des paramètres physiques du milieu sont suivis. De par le fort caractère indicateur des conditions du milieu qu’ils présentent et leur importance dans le réseau trophique, les macroinvertébrés aquatiques font l’objet d’un suivi particulier et complémentaire dans le cadre de l’expérimentation des lâchers.
L’étude DREAM1 a donc pour objectif de dégager les mécanismes de réponses du macrobenthos
vis-à-vis des lâchers d’eau dans le tronçon de l’Escale-Cadarache, en particulier en étudiant les processus de recolonisation des habitats lotiques par les macroinvertébrés suite à ces lâchers. Le principe est de suivre les peuplements d’invertébrés à un pas de temps serré et adapté à leur cycle vital. L’intégration des aspects pouvant influencer cette dynamique de recolonisation est nécessaire ; ainsi le développement du périphyton sur les substrats est considéré comme un élément primordial dans la compréhension des processus de recolonisation des macroinvertébrés. Ce compartiment biologique qu’est le périphyton fait l’objet d’un suivi en parallèle dont les résultats ne sont pas présentés dans ce rapport. Les résultats présentés concernent exclusivement les données de prélèvements des macro-invertébrés collectées durant les cinq premiers mois de l’année 2014. Sur le plan de la définition de la méthodologie à adopter pour le suivi d’une gestion hydrologique expérimentale, certaines préconisations avaient déjà été avancées en conclusion de
l’expérimentation de régime réservé à Ste-Tulle2.
L’objectif fonctionnel de gestion apporté par les lâchers de décolmatage est le maintien de la capacité d’accueil des substrats dans les habitats lotiques, principalement pour favoriser la reproduction d’espèces de poissons rhéophiles durant leur période de fraie. Le compartiment benthique est donc suivi en tant qu’indicateur de la qualité d’habitat des zones lotiques. « La
diversité des taxons d’invertébrés benthiques en cours d’eau étant dominée par les insectes, il convient donc d’adapter la production de connaissances à l’échelle du déroulement des cycles biologiques majoritairement monovoltins (une génération par an) » 2.
De plus, la période d’étude devrait commencer « assez précocement, dès la fin de l’hiver, afin de bien prendre en compte la dynamique de production primaire algale qui monte en puissance
proportionnellement à l’allongement de la période diurne »2. Le plan d’échantillonnage a été défini
afin que la fréquence des campagnes de prélèvements soit courte (entre un mois et un mois et demi), rythmée dans une chronique saisonnière.
Les habitats ciblés sont les milieux lotiques et les populations d’invertébrés rhéophiles sont essentiellement visées par les protocoles d’échantillonnage. Les prélèvements se réduisent aux mésohabitats dont les vitesses de courant de surface sont supérieures ou égales à 25 cm/s ; des réplicas sont effectués dans des gammes de gradient de vitesse différentes. Les granulométries des habitats prospectés doivent être homogènes. Les taxons appartenant aux ordres des plécoptères,
1
DREAM : Dynamique de REcolonisation des Algues et Macroinvertébrés
2
Renouvellement de la concession de St-Tulle. Etude expérimentale d’un régime réservé en moyenne Durance. Etat initial 2005-2007 / essai 2008-2011. Macro-invertébrés benthiques. Mars 2013. UR HYAX, Irstea Aix en Provence
7 des éphéméroptères et des trichoptères (PET) font l’objet en laboratoire de mesure des tailles des larves afin de distribuer leurs effectifs au sein de classes de taille.
1.3 Contexte hydro-climatique
1.3.1 Evolution temporelle des débits lors des sept premiers mois de 2014
Les débits représentés dans la figure 1 ont été obtenus à partir des données de débits de déversés du barrage de l’Escale (fournies par Edf) auxquelles sont ajoutées, en période d’absence de déversés, les valeurs théoriques de débits réservés dans le tronçon Escale-Cadarache. Ces valeurs théoriques de
débits réservés dans ce tronçon sont de 6,1 m3/s (soit 1/20ème du module) d’octobre à mars, et de 8,7
m3/s (soit 1/14ème du module) d’avril à septembre.
Les valeurs de débits présentées dans la figure 1 ne reflètent pas les conditions hydrologiques qui ont eu cours au sein de nos stations d’étude puisqu’elles n’intègrent pas les apports du bassin-versant intermédiaire entre le barrage de l’Escale et les stations d’étude. Elles permettent néanmoins d’approximer les tendances hydrologiques, depuis que le seul limnimètre dans le tronçon de l’Escale installé au seuil de la Brillanne (DREAL PACA) a été rendu inutilisable par la cassure de cet ouvrage depuis le mois de mai 2013.
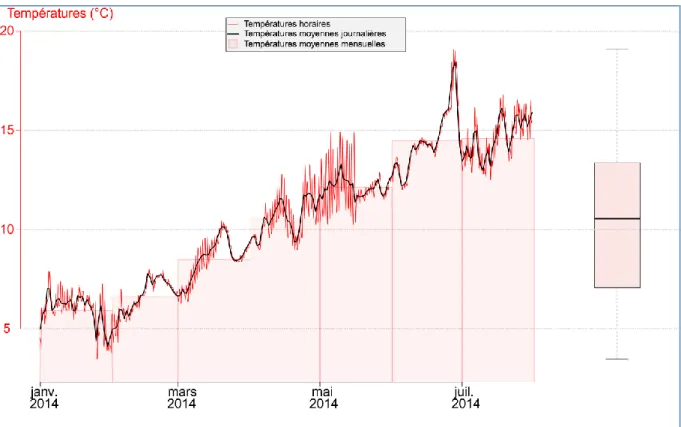
Figure 1 : Evolution des débits au pas de temps horaire (tracé bleu) et des débits moyens mensuels (rectangles bleus) de la Durance en aval du barrage de l’Escale entre le 1er janvier et le 31 juillet 2014. L’axe des ordonnées est représenté avec une échelle logarithmique. Les débits maximaux atteints lors des épisodes de crues sont indiqués au-dessus de la courbe.
La boite à moustache dans la partie droite concerne les données de la période considérée. Source : Edf.
En janvier 2014 et à la suite de forts épisodes de précipitations, un épisode de crue de forte intensité
s’est produit (Figure 1). Les débits enregistrés ont dépassé le seuil de 1000 m3/s lors de la journée du
19 janvier 2014 durant une période d’environ 4 heures. Une telle intensité de débit, largement supérieure à celle prévue lors d’un lâcher artificiel, a engendré une mobilisation sédimentaire et
8 donc un décolmatage complet des substrats. Le lâcher artificiel d’eau prévu a donc été annulé pour cette année. En conséquence, les objectifs de l’étude pour l’année 2014 ont été réajustés autour de la dynamique de recolonisation des macroinvertébrés suite à cet épisode de crue.
En raison d’opérations techniques sur l’ouvrage du barrage de l’Escale, les valeurs de débits déversés
entre le 26 février et le 2 juillet étaient proches de 18 m3/s. Ces débits, largement supérieurs aux
valeurs de débit réservé (6,1 m3/s et 8,7 m3/s), ont été observés durant les quatre campagnes de
prélèvements que nous avons effectuées.
1.3.2 Evolution des températures lors des sept premiers mois de 2014
La Figure 2 présente l’évolution des températures (horaires, journalières et mensuelles) enregistrées
entre le 1er janvier et 31 juillet 2014 à une distance de moins d’un kilomètre en aval du barrage de
l’Escale. Ces données sont issues d’enregistrements réalisés par le Syndicat Mixte d’Aménagement
de la Vallée de la Durance3 (SMAVD). Elles constituent les seules données thermiques disponibles
pour la Durance dans le tronçon Escale-Cadarache en 2014. Tout comme les données de débits présentées dans le chapitre précédent, ces températures n’ont pas été acquises au niveau des stations d’étude, mais plus de 20 km en amont de la station D0BIS. En absence d’enregistrement plus proche de nos stations d’étude, ces valeurs permettent tout de même d’observer les tendances d’évolution thermique dans ce tronçon de Durance.
Figure 2 : Evolutions des températures au pas de temps horaire (tracé rouge) et des moyennes journalières (tracé noir) et mensuelles (rectangles rouges) des eaux de la Durance environ 900 m en aval du barrage de l’Escale (longitude : 6,0076905 ; latitude : 44,077718 en degrés décimaux) entre le 1er janvier et le 31 juillet 2014. La boite à moustache dans
la partie de droite concerne les données de la période considérée. Source : SMAVD.
3
9
Au regard de l’évolution des températures entre le 1er janvier et le 15 juillet 2014 (Figure 2), une
augmentation quasi-continue est observée durant cette période. De fortes variations journalières sont observées durant certaines périodes. Le lieu d’enregistrement des températures situé 900 mètres en aval du barrage de l’Escale et donc à une distance limitant un réchauffement des eaux depuis leur sortie du barrage de l’Escale, pourrait expliquer ces valeurs peu élevées en début d’été.
1.4 Orientations de l’étude DREAM pour l’année 2014
Le lâcher de décolmatage a été annulé suite à l’épisode de crue qui s’est déroulé à la fin du mois de janvier 2014. Les objectifs de l’étude DREAM ont donc été réajustés vers le suivi de la dynamique de recolonisation des peuplements de macroinvertébrés benthiques suite à cet épisode en maintenant le plan d’échantillonnage initialement prévu. Dans le cadre de l’identification de l’influence des opérations de lâcher de décolmatage en Durance sur la macrofaune benthique, les résultats de l’étude en 2014 pourront servir de référence de dynamique de recolonisation suite à un épisode naturel décolmatant de forte intensité.
10
2 Matériels et méthodes
2.1 Stations d’étude
Deux stations dans le tronçon de l’Escale-Cadarache ont été suivies en 2014.
Carte 2 : Localisation des stations d’étude D0BIS et D1 dans le tronçon de l’Escale-Cadarache.
La station D0BIS4 se situe sur la commune d’Oraison, soit à plus de 26 km en aval du barrage de
l’Escale. En 2014, la morphologie de la Durance dans cette station était constituée d’un chenal unique.
La station D15 se situe sur la commune de Volx, à proximité de Manosque, environ 32 km en aval du
barrage de l’Escale. En 2014, une île de gravier séparait deux chenaux au niveau de la station.
4 Coordonnées en degrés décimaux : longitude = 5.884836 ; latitude = 43.88876 5
11
Photo 1 : Stations D0BIS à gauche et D1 à droite en 2014 (photos : Irstea).
Ces stations étaient toutes deux déjà suivies entre 2005 et 2011 dans le cadre de l’étude du
renouvellement de la concession de Ste-Tulle. Initialement, la localisation de ces stations avait été choisie afin d’encadrer le point de restitution du débit complémentaire nécessaire à l’expérimentation Ste-Tulle se situant entre la confluence de l’Asse et celle du Largue. Ces stations ont été conservées pour l’étude DREAM en raison des bonnes connaissances acquises sur leur fonctionnement biologique, et parce qu’elles encadrent les confluences de l’Asse, du Largue et de la Durance. Cela permet ainsi d’étudier un secteur amont, fonctionnant sous régime hydrologique réservé auquel s’ajoute les apports non-dérivés des bassins-versants intermédiaires, et un secteur en aval de l’Asse bénéficiant de la naturalité du débit d’un bassin-versant important.
L’Asse est l’un des principaux affluents de la Durance dans le tronçon de l’Escale-Cadarache. Cette rivière n’étant pas aménagée par des barrages, les épisodes de petites et moyennes crues écrêtées en Durance y sont plus fréquents. Toutefois, jusqu’en 2007, une digue fusible située dans la Durance entre la confluence de l’Asse et du Largue permettait de dériver une partie des apports du bassin-versant intermédiaire dont celui de l’Asse. La gestion de cette digue ayant été abandonnée depuis, l’intégralité des débits de l’Asse transitent naturellement dans la Durance.
12
2.2 Campagnes d’échantillonnage
Quatre campagnes d’échantillonnage ont été effectuées sur les stations D0BIS et D1 en 2014. Le Tableau 2 présente les dates des campagnes et les intervalles de temps entre celles-ci. Lors de chaque campagne, les deux stations ont été prospectées durant la même journée.
Tableau 2 : Dates des campagnes de prélèvements réalisées en 2014 et intervalles de temps entre deux campagnes consécutives.
Date Intervalle entre deux
campagnes (jours) Campagne 1 27/02/2014 Campagne 2 17/03/2014 18 Campagne 3 16/04/2014 30 Campagne 4 20/05/2014 34
2.3 Protocole de prélèvement
2.3.1 Zones de prélèvementPour suivre les effets d’un lâcher de décolmatage puis ceux des phénomènes de colmatage ultérieurs sur les habitats lotiques, le plan d’échantillonnage défini ne concerne que des habitats dont la vitesse d’écoulement de surface est supérieure à 30 cm/s.
Trois ambiances d’écoulements ont été suivies. Elles sont repérées par leurs vitesses d’écoulement de surface. Le principe du suivi est de mettre en lien les différences faunistiques et habitationnelles entre des ambiances d’écoulements contrastées et représentatives des habitats lotiques de la moyenne-Durance avec les conditions abiotiques, et particulièrement les conditions hydrauliques des habitats. Toutes ces ambiances ont été sélectionnées dans un faciès hydromorphologique de type « radier ».
Ces ambiances d’écoulements correspondent aux gammes de vitesses de surface suivantes :
Ambiance 1 (A1) : vitesses d’écoulement de surface entre 30 et 50 cm/s.
Ambiance 2 (A2) : vitesses d’écoulement de surface entre 70 et 90 cm/s.
Ambiance 3 (A3) : vitesses d’écoulement de surface supérieures à 100 cm/s.
Lors de chaque campagne, trois échantillons ont été effectués au sein de chaque ambiance d’écoulement. Au total, 9 échantillons de macrofaune benthique ont été prélevés par station et par campagne.
2.3.2 Méthode de prélèvement et description des variables d’habitat
Les échantillons ont été effectués à l’aide d’un filet Surber de surface d’1/10ème m² et équipé d’un
filet d’une taille de vide de maille de 250 µm. Les échantillons ont ensuite été fixés dans une solution contenant 4 % de formaldéhyde.
13 En parallèle de l’échantillonnage des macroinvertébrés, les paramètres suivants, descriptifs des conditions des micro-habitats, ont été relevés :
la granulométrie du substrat6 en distinguant le substrat de surface et celui en sous-couche
(granulométrie dominante, granulométrie des éléments les plus grossiers, présence de granulométrie accessoire),
la structure et la stabilité des éléments du substrat,
la présence de végétation aquatique et l’importance de son recouvrement et de son
développement,
le développement du biofilm,
la présence de dépôts et leur nature (par exemple pour des dépôts de limon),
la hauteur d’eau totale,
la vitesse de courant à 3 cm du fond,
les vitesses de courant à 0,2 ; 0,4 et 0,8 fois la hauteur d’eau,
l’estimation visuelle du colmatage de surface du substrat.
Pour ce dernier paramètre d’habitat : le colmatage de surface du substrat par des sédiments fins ou
des limons, a été estimé en appliquant le protocole décrit dans une note technique du Cemagref7.
Cette méthode consiste à attribuer un code variant de 1 à 5, pouvant également être exprimé en classe de pourcentage, en fonction de plusieurs critères comme l’épaisseur des dépôts, la cohésion des éléments minéraux de surface et de sous-couche, l’intensité de la bouffée turbide dégagée par la déstructuration du substrat de surface (Figure 3).
6 Malavoi, J.R. et Souchon, Y. – 1989 : Méthodologie de description et quantification des variables
morphodynamiques d'un cours d'eau à fond caillouteux. Exemple d'une station sur la Filière (Haute Savoie). -
Rev. Géogr. Lyon 64: 252-259.
7 Description du substrat minéral et du colmatage. Note technique provisoire, 2005. Archambaud G., Giordano
14
Figure 3 : Représentation simplifiée du protocole d‘estimation du colmatage du substrat. La première colonne correspond aux codes de colmatage (chiffres en rouge), la seconde colonne correspond aux classes exprimées en pourcentage et la dernière colonne schématise le substrat en coupe latérale avec l’épaisseur des dépôts de surface, la cohésion des éléments minéraux et l’intensité de la bouffée turbide engendrée lors de la déstructuration du substrat.
2.3.3 Tri et identification des macroinvertébrés
Au laboratoire, les échantillons ont été tamisés sur une colonne de tamis dont le plus fin présente une taille de vide de maille de 650 µm. Cette taille de vide de maille présente l’intérêt de ne pas conserver les plus petites larvules souvent difficilement identifiables.
Les individus récoltés ont ensuite été identifiés à des niveaux de détermination variable selon les taxons, puis dénombrés. A l’exclusion des diptères, les insectes ont été identifiés au niveau du genre ou de l’espèce (notamment pour la famille des Baetidae (éphéméroptères)). Les diptères ont été identifiés aux niveaux taxonomiques de la famille, de la sous-famille ou de la tribu. Les taxons n’appartenant pas à la classe des insectes ont été identifiés au niveau de la sous-classe (par exemple les oligochètes), ou à des niveaux taxonomiques plus précis.
Afin d’adopter les mêmes règles pour effectuer des comparaisons valables, les niveaux de détermination de la liste faunistique finale ont été harmonisés et les différents stades larvaires regroupés par taxon (à l’exception des coléoptères de la famille des Elmidae dont les stades larvaires possèdent des préférendums écologiques variés).
15
Les individus appartenant aux ordres des PET8 ont été mesurés en classes de taille adaptées aux
tailles maximales de chaque taxon et reflétant les principaux stades larvaires. Les limites des classes de taille exprimées en mm sont les suivantes :
- [1 ; 3[, [3 ; 5[, [5 ; 7[, [7 ; 10[, [10 ; 15[, [15 ; 20[, [20 ; 25[ : pour tous les taxons sauf
Hydropsyche,
- [0 ; 5[, [5 ; 10[, [10 ; 15[, [15 ; 20[, [20 ; 25[ : pour les Hydropsyche.
8
16
3 Résultats
3.1 Approches globales
Sont d’abord présentées dans ce chapitre les données hydrologiques et thermiques des 30 jours précédant les campagnes de prélèvement, puis celles relatives à certaines variables d’habitats, telles que les conditions hydrauliques et les estimations de degré de colmatage des substrats. Ces résultats permettent de comparer les conditions observées au sein des trois ambiances d’écoulements. Puis, un premier bilan des principales métriques décrivant les structures faunistiques échantillonnées en 2014 est présenté, suivi par une comparaison taxonomique entre les ambiances d’écoulement.
3.1.1 Données hydrologiques et thermiques
3.1.1.1 Historique hydrologique des campagnes
Afin de caractériser les conditions d’hydrologie ayant influencé les peuplements de macroinvertébrés, le graphique suivant présente les distributions des valeurs de débits pour des périodes de 30 jours précédant chacune des campagnes. Cette durée de 30 jours reste arbitraire et a été définie dans l’objectif de refléter l’histoire récente des conditions abiotiques, l’expression biologique d’une communauté étant régulée par les processus de bio-intégration des conditions environnementales.
Figure 4 : Distribution des débits horaires des 30 jours précédant chaque campagne de prélèvement. L’axe des ordonnées est représenté avec une échelle logarithmique.
La Figure 4 permet de distinguer nettement les conditions de débits des 30 jours précédant la première campagne (27 février) de celles des trois autres campagnes : des débits supérieurs à 200
m3/s ont été enregistrés. Les campagnes C2 (17 mars), C3 (16 avril) et C4 (20 mai) ont été effectuées
17 Le Tableau 3 présente le nombre d’occurrence de dépassement vis-à-vis de six valeurs seuils par les débits à un pas de temps horaire au sein de ces périodes de 30 jours précédant chacune des campagnes de prélèvements. Ces valeurs reflètent la notion de fréquence de perturbation due aux épisodes de crues.
Tableau 3 : Bilans des occurrences de dépassements de valeurs seuils par les des débits horaires durant des périodes de 30 jours précédant chacune des campagnes (744 valeurs).
Campagnes de prélèvements C1 C2 C3 C4 Q > 20 m3/s 374 153 0 1 Q > 30 m3/s 235 71 0 1 Q > 50 m3/s 206 43 0 0 Q > 100 m3/s 41 0 0 0 Q > 200 m3/s 4 0 0 0 Q > 250 m3/s 3 0 0 0
Le nombre d’occurrences de dépassement de valeurs de débits élevés diminue progressivement d’une campagne à la suivante. La fréquence de perturbation engendrée sur les peuplements de macroinvertébrés est donc maximale lors de la campagne de février, puis diminue jusqu’à un niveau pouvant être considéré comme nul lors des campagnes C3 et C4.
Une métrique supplémentaire a été utilisée : il s’agit de l’antériorité de l’épisode de crue le plus intense du début de l’année 2014 (celui du 19 janvier 2014) aux campagnes de prélèvements. Les valeurs sont présentées dans le Tableau 4.
Tableau 4 : Antériorité en nombre de jours de l’épisode de crue du 19 janvier 2014 aux campagnes de prélèvements.
Campagnes
C1
C2
C3
C4
Dates 27/02/2014 17/03/2014 16/04/2014 20/05/2014
Antériorité 38 jours 56 jours 86 jours 120 jours
Ces valeurs d’antériorité représentent l’éloignement des observations faunistiques dont nous disposons à un épisode de forte morphogénèse. Les campagnes de prélèvements constituent ainsi différentes images dans les étapes de dynamique de recolonisation et de résilience des peuplements benthiques.
3.1.1.2 Historique thermique des campagnes de prélèvement
De manière identique à la démarche adoptée pour caractériser l’histoire hydrologique récente des campagnes de prélèvement, la Figure 5 illustre la distribution des températures horaires sur une période de 30 jours précédant chacune des campagnes.
18
Figure 5 : Distribution des températures horaires des 30 jours précédant chaque campagne de prélèvement.
Les distributions des températures enregistrées sur les 30 jours précédant les campagnes montrent une augmentation continue. Ce signal saisonnier fort est donc attendu comme conditionnant l’apparition taxonomique de taxons à développement larvaire printanier et, dans un second temps, estival.
En conclusion, l’histoire hydrologique récente des campagnes de prélèvements permet de distinguer trois groupes : (i) la première campagne, avant laquelle des conditions de nervosité hydrologique
ont eu cours ; (ii) la seconde campagne, avant laquelle les débits ont dépassé les 50 m3/s ; (iii) et les
deux dernières campagnes effectuées dans des périodes de stabilité hydrologique malgré le dépassement des valeurs de débits réservés théoriques.
L’accroissement continu des températures des eaux place les campagnes de prélèvements dans une dynamique printanière caractéristique.
Une difficulté interprétative des données faunistiques va alors résider dans la distinction de l’influence de ce signal thermique saisonnier et de celui pouvant être résumé par l’antériorité de l’épisode de crue du 19 janvier 2014 et les histoires hydrologiques récentes.
19
3.1.2 Données mésologiques
Dans un premier temps, les variables relatives aux conditions hydrauliques au sein des ambiances d’écoulements sont présentées, suivies par les résultats d’estimation du degré de colmatage des substrats.
3.1.2.1 Variabilité des conditions d’habitat entre les ambiances d’écoulement
Dans ce chapitre, les conditions d’habitats sont comparées entre les ambiances d’écoulement à partir des valeurs de vitesses mesurées lors des prélèvements benthiques et des compositions granulométriques des substrats.
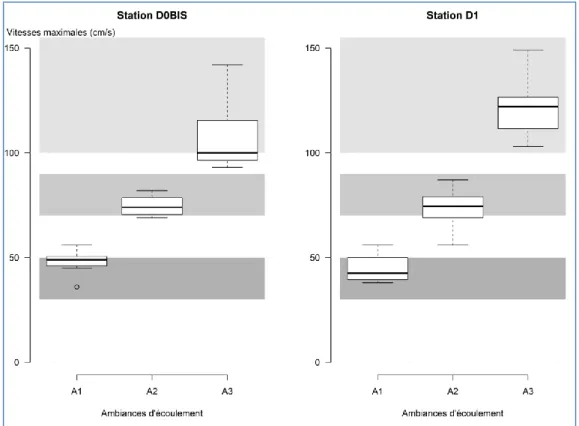
La Figure 6 présente la distribution des valeurs maximales de vitesses de courant mesurées au sein de la colonne d’eau, de chaque prélèvement, par ambiance hydraulique et pour les deux stations d’étude. Pour rappel, les vitesses de courant ont été mesurées à 3 cm du fond, puis à 0,2, 0,4 et 0,8 fois la hauteur d’eau. Les vitesses maximales obtenues à partir des mesures des vitesses à ces quatre hauteurs ne correspondent donc pas à la vitesse des écoulements de surface, mais permettent d’étudier la loticité des places de prélèvement.
Figure 6 : Distributions des valeurs de vitesses maximales dans la colonne d’eau en fonction des ambiances d’écoulements pour les deux stations d’étude. Les trois zones grisées correspondent aux valeurs ciblées de vitesse de surface pour chaque ambiance (A1 : gris foncé entre 30 et 50 cm/s ; A2 : gris intermédiaire entre 70 et 90 cm/s ; A3 : gris
clair pour des vitesses > 100 cm/s)
Bien que les ambiances d’écoulement ciblées par le protocole soient définies in-situ par les vitesses de courant de surface, ces valeurs maximales de vitesse dans la colonne d’eau apparaissent globalement en concordance avec la définition des limites de classes des ambiances. Dans l’ambiance A3, les vitesses maximales dans la colonne d’eau sont plus élevées en D1 qu’en D0BIS.
A partir de la description de la taille des éléments granulométriques et de leur recouvrement relatif au sein de la surface échantillonnée, une granulométrie moyenne par Surber peut être obtenue
20 (Figure 7). Pour rappel, tous les échantillons ont été effectués dans des faciès géomorphologiques de type « radier ».
Figure 7 : Distribution des granulométries moyennes par ambiance d’écoulement pour les deux stations d’étude. Les codes granulométriques utilisés sont : 1 –vase (< 4 µm), 2 –limon (> 4 µm), 3 –sable fin (> 62 µm), 4 –sable grossier (> 0,5
mm), 5 –gravier fin (> 2 mm), 6 –gravier grossier (> 8 mm), 7 –cailloux fin (> 16 mm), 8 –cailloux grossier (> 32 mm), 9 – pierre fine (> 64 mm), 10 –pierre grossière (> 128 mm), 11 –bloc fin et moyen (> 256 mm), 12 –bloc grossier (>1024 mm),
13 –bloc très grossier (> 2048 mm), 14 –végétal.
Les résultats présentés en Figure 7 illustrent le fait que l’ensemble des échantillons effectués en 2014 ont été réalisés dans des structures granulométriques comparables, variant du caillou fin à grossier jusqu’aux blocs fins.
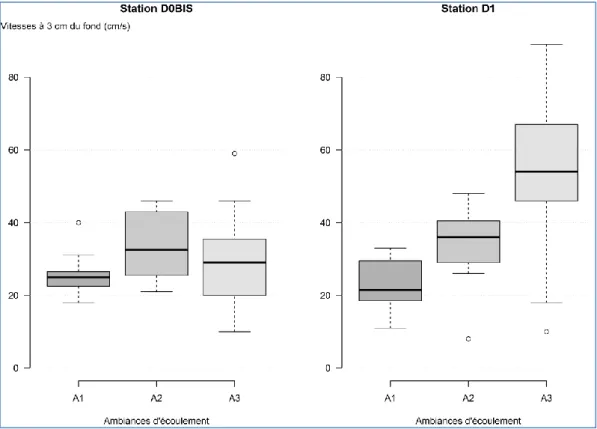
Les éléments granulométriques prélevés dans la station D1 apparaissent légèrement plus petits, et plus homogènes en taille, que ceux de la station D0BIS. Pour la station D0BIS, une plus faible variabilité dans la taille des éléments prélevés dans l’ambiance 3 est observée, les éléments étant principalement représentés par des pierres fines et des pierres grossières pour cette ambiance. La Figure 8 présente la distribution des vitesses de courant mesurées à 3 cm du fond lors de chaque prélèvement par ambiance d’écoulement. Ces vitesses de fond permettent d’appréhender les conditions hydrauliques régnant dans la strate inférieure de la rugosité du fond.
21
Figure 8 : Distributions des vitesses de courant mesurées à 3 cm du fond par ambiance d’écoulement pour les deux stations d’étude.
Logiquement, les valeurs de vitesses de fond (Figure 8), mesurées à 3 cm de hauteur d’eau, sont nettement inférieures aux vitesses de surface utilisées pour définir les ambiances d’écoulement ciblées.
En comparant les distributions de ces valeurs entre les deux stations, les conditions hydrauliques de fond apparaissent semblables dans les ambiances A1 et A2 entre les stations. La principale différence est observée dans l’ambiance A3 : les vitesses de fond sont supérieures dans la station D1. La taille des éléments granulométriques de l’ambiance A3, légèrement supérieure dans la station D0BIS par rapport à D1 (Figure 7), induit que la rugosité de fond y soit plus importante, et donc diminue les écoulements au contact des substrats. De plus, les vitesses maximales dans la colonne d’eau étant plus élevées dans l’ambiance A3 en D1 (Figure 6), il apparait logique que les vitesses de fond mesurées y soit plus importantes.
L’hypothèse d’une présence accrue de taxons rhéophiles dans l’ambiance A3 de la station D1 par rapport à D0BIS peut donc être avancée.
3.1.2.2 Degrés de colmatage des substrats
3.1.2.2.1 Bilan par ambiances d’écoulement
La Figure 9 présente les fréquences d’occurrence des trois classes de colmatage observées au cours des campagnes de l’année 2014 par ambiance d’écoulement. Ces estimations de degrés de colmatage ont été obtenues à partir du protocole d’estimation visuel détaillé dans le paragraphe 2.3.2. Pour chaque ambiance dans une station donnée, ces fréquences d’occurrence sont calculées à partir de l’ensemble des données sur les quatre campagnes, soit :
22 (3 échantillons / campagne) x 4 campagnes = 12 prélèvements
Figure 9 : Fréquence d’occurrence des trois classes de colmatage observées en 2014 par ambiance d’écoulement pour les deux stations d’étude. Pour rappel : classe 1 – colmatage estimé entre 0 et 25 % ; classe 2 – entre 25 et 50 % ; classe 3 –
entre 50 et 75 %.
A partir des résultats présentés dans la figure ci-dessus, il apparait en premier lieu que la classe de colmatage maximale observée entre février et mai 2014 est la classe 3. Cela traduit des degrés de colmatage qui sont restés modérés, certainement dus à la forte mobilisation sédimentaire crée par l’épisode de crue morphogène du 19 janvier et à la limitation d’apports ultérieurs. Le maintien de débits supérieurs à la valeur théorique de débits réservé durant toute la période d’étude a certainement contribué de surcroit à limiter les dépôts sédimentaires, surtout dans les écoulements les plus rapides.
Au niveau des divergences entre les ambiances d’écoulement, il apparait que seules certaines estimations de colmatage dans l’ambiance la plus lente, l’ambiance A1 (vitesses de surface comprises entre 30 et 50 cm/s), ont été attribuées en classe 3. L’ambiance A2 a comporté seulement des degrés de colmatage limités aux deux premières classes. Les niveaux de colmatage de l’ambiance A3 sont restés nuls à faibles.
Seules les ambiances A1 présentent des divergences entre les stations. Les substrats de l’ambiance A1 de la station D1 ont été qualifiés en classe 3 plus fréquemment que ceux de la station D0BIS. Cela peut provenir de la structure de tressage en deux chenaux de cette station ; le débit, même s’il est supérieur à celui de D0BIS grâce aux apports du bassin-versant intermédiaire, y est partagé de manière inégale entre les deux chenaux, diminuant la puissance de l’écoulement et pouvant donc favoriser les dépôts sédimentaires. Ces niveaux de colmatage plus importants observés dans la station D1 pourraient également être dus à des apports supplémentaires de sédiments en provenance de la rivière de l’Asse.
23
3.1.2.2.2 Évolution temporelle du colmatage par ambiances d’écoulement
La Figure 10 présente l’évolution temporelle de la moyenne des indices d’estimation visuelle de colmatage par ambiance. A partir des classes de colmatage attribuées aux substrats prélevés, une valeur moyenne peut être estimée pour chaque ambiance.
Figure 10 : Variation de l’indice moyen de colmatage des ambiances d’écoulement par campagne et indice moyen par campagne pour les deux stations d’étude.
Les résultats présentés dans la Figure 10 permettent de comparer les évolutions des degrés de colmatage entre ambiance dans les deux stations d’étude.
Les degrés de colmatage les plus importants sont observés lors de la dernière campagne en D0BIS, et lors des deux dernières campagnes en D1. Cette dynamique de colmatage, à la suite d’un décolmatage complet présumé lors de la crue de la fin du mois de janvier, apparait donc plus précoce dans la station D1. Pour reprendre les éléments d’explication formulés préalablement, la Durance au niveau de la station D1 se compose de deux chenaux et reçoit les apports sédimentaires de la confluence de l’Asse.
En conclusion, les prélèvements réalisés durant les quatre campagnes de l’année 2014 ont été effectués en accord avec le protocole préalablement établi : les habitats lotiques ont été échantillonnés dans des gammes de vitesses en concordance avec les limites des ambiances d’écoulement ciblées ; les granulométries échantillonnées apparaissent homogènes.
La granulométrie de l’ambiance A3 dans la station D0BIS comportait des éléments plus grossiers que celle de la station D1, limitant la vitesse de courant de fond.
Les degrés de colmatage estimés durant l’ensemble des campagnes sont restés très limités. Dans l’ambiance A1, une légère augmentation du colmatage moyen est observée dès le mois d’avril en D1 et un mois plus tard en D0BIS.
24
3.1.3 Bilan faunistique global et par station d’étude
3.1.3.1 Bilans par station
Au total, 4 campagnes d’étude ont été effectuées sur les deux stations D0BIS et D1. Un total de 9 échantillons a été prélevé lors de chaque campagne par station. Le nombre total d’échantillons traités en 2014 s’élève donc à 72. Les listes faunistiques obtenues sont présentées en annexes. Un premier bilan des prélèvements de macroinvertébrés benthiques réalisés en 2014 est présenté dans le Tableau 5.
Tableau 5 : Bilan global et par station des effectifs bruts, des densités, des richesses taxonomiques et du nombre de taxons représenté par plus d’un individu.
Total
Station D0BIS
Station D1
Nombre d'individus 17461 2922 14539
Densité (ind. / m²) 2425 811 4038
Richesse taxonomique 52 34 47
Richesse > 1 ind. 35 26 33
En cumulant les résultats des deux stations d’étude lors des quatre campagnes de prélèvement, 17461 individus ont été prélevés en 2014 au sein des 72 échantillons. En ramenant cet effectif à la surface représentée par ces 72 échantillons, représentant chacun une surface de prélèvement de 0,1 m², une densité de près de 2425 individus / m² est obtenue en prenant en compte les deux stations d’étude.
Toujours au niveau global, 52 taxons ont été observés en 2014 dans l’ensemble des échantillons (diversité γ), dont 35 taxons étaient représentés par plus d’un individu, soit une proportion de taxons pouvant être qualifiés de « rare » avoisinant plus de 30 % de la richesse taxonomique.
Le pourcentage de taxons communs (indice de similarité de Jaccard) entre les peuplements des deux stations est de 55,8 %. En retirant les taxons représentés par un seul individu, ce pourcentage de similarité qualitative atteint 74 %.
5 taxons ont été rencontrés uniquement dans la station D0BIS : Dinocras (plécoptère), Dytiscidae (coléoptère), Onychogomphus (odonate), Limoniini (diptère) et Pisidium (gastéropode). Ils ont tous été représentés par un seul individu. La somme de leurs effectifs représente 0,001 % des effectifs de D0BIS.
18 taxons ont été rencontrés uniquement dans la station D1 : Euleuctra geniculata, Nemoura (plécoptères), Baetis pavidus, Procloëon, Ephemera, Heptagenia (éphéméroptères), Elmis, Esolus (coléoptères), Boyeria (odonate), Anthomyidae, Dolichopodidae, Dicranota, Pediciini, Eriopterini,
Stratiomyidae, Tabanidae (diptères), Dugesia (turbellarié) et Potamopyrgus (gastéropode). Les
taxons notés en gras étaient représentés par plus d’un individu. La somme de leurs effectifs représente 0,003 % des effectifs de D1.
Au niveau des différences entre stations d’étude, les peuplements de la station D1 apparaissent plus riches et nettement plus denses que ceux de la station D0BIS. Une densité presque cinq fois plus faible a été observée en D0BIS par rapport à celle de la station D1.
25
3.1.3.2 Bilan des principaux groupes faunistiques
Le Tableau 6 présente les effectifs, les richesses taxonomiques et le nombre de taxons représenté par plus d’un individu en valeurs brutes et en valeurs relatives par grands groupe faunistique. Sont distingués ici : les ordres appartenant à la classe des insectes, l’ensemble constitué par les EPT, et un regroupement des taxons n’appartenant pas à la classe des insectes.
Tableau 6 : Bilans global et par station des effectifs, des richesses taxonomiques et du nombre de taxons représenté par plus d’un individu pour les principaux groupes faunistiques. Les effectifs et les richesses sont exprimés en valeurs brutes
et en valeurs relatives (Tableau 5).
Total
Station D0BIS
Station D1
bruts relatifs bruts relatifs bruts relatifs
INS
ECTES
Plécoptères Effectif 208 1.9% 85 2.9% 123 0.8% Richesse 6 11.5% 4 11.8% 5 10.6% Rich. >1ind 4 11.4% 2 7.7% 4 12.1% Ephéméroptères Effectif 4607 26.4% 1050 35.9% 3557 24.5% Richesse 16 30.8% 12 35.3% 16 34.0% Rich. >1ind 12 34.3% 11 42.3% 11 33.3% Trichoptères Effectif 587 3.4% 56 1.9% 531 3.7% Richesse 3 5.8% 3 8.8% 3 6.4% Rich. >1ind 3 8.6% 3 11.5% 3 9.1% Total PET Effectif 5402 30.9% 1191 40.8% 4211 29.0% Richesse 25 48.1% 19 55.9% 24 51.1% Rich. >1ind 19 54.3% 16 54.9% 18 54.5% Coléoptères Effectif 26 0.1% 2 0.1% 24 0.2% Richesse 4 7.7% 2 5.9% 3 6.4% Rich. >1ind 3 8.6% 0 0.0% 2 6.1% Odonates Effectif 2 0.0% 1 0.0% 1 0.0% Richesse 2 3.8% 1 2.9% 1 2.1% Rich. >1ind 0 0.0% 0 0.0% 0 0.0% Diptères Effectif 11569 66.3% 1668 57.1% 9901 68.1% Richesse 16 30.8% 9 26.5% 15 31.9% Rich. >1ind 9 25.7% 8 30.8% 9 27.3%HORS INSECTES
Effectif 462 2.6% 60 2.1% 402 2.8% Richesse 5 9.6% 3 8.8% 4 8.5% Rich. >1ind 4 11.4% 2 7.7% 4 12.1%En cumulant les résultats des deux stations (première colonne du Tableau 6), les diptères dominent en nombre d’individus et en richesse. L’ordre des éphéméroptères représente le second ordre dominant en effectif et en richesse.
Au regard des différences faunistiques entre les stations (deuxième et troisième colonnes du Tableau 6), les peuplements de la station D1 présentent des richesses plus élevées que ceux de D0BIS essentiellement pour les ordres des diptères et des éphéméroptères. Cependant, les valeurs de richesses des taxons représentés par plus d’un individu sont très proches entre les deux stations pour la quasi-totalité des groupes faunistiques. Le fait que les richesses soient plus élevées dans la station
26 D1 apparait donc principalement en lien avec la présence de taxons rares. De surcroit, les effectifs représentés par l’ensemble de ces taxons rares est de 0,2 % des effectifs totaux en D0BIS, et de 0,1 % de ceux de D1.
Les effectifs bruts des peuplements de D1 sont toujours supérieurs à ceux de D0BIS et cela pour l’intégralité des groupes faunistiques. Plusieurs hypothèses peuvent être avancées pour expliquer cette observation, parmi lesquelles :
- le fait que la présence de deux chenaux dans la station D1 (un seul chenal en D0BIS) pourrait
lui conférer un potentiel de désordre minimisé par l’étalement du flot et une dispersion spatiale accentuée de l’énergie de l’écoulement lors des épisodes de morphogénèse notamment ; cela enduirait une diminution de l’intensité de perturbation sur la macrofaune benthique. Cette explication semble peu probable car les niveaux supérieurs de densités en D1 par rapport à D0BIS sont constants depuis 2005, période durant laquelle cette station a présenté un caractère monochenal.
- la présence potentielle d’apports trophiques en amont de la station D1. Ces apports pouvant
provenir d’affluents comme le Largue et l’Asse ou de rejets d’eaux usées proches.
- La situation géomorphologique particulière de ce site, situé en aval des confluents de trois
nappes alluviales, Durance, Asse et Largue.
Au-delà de ce constat sur les densités, le résultat suivant apparait intéressant : pour ce qui est de la répartition des effectifs au sein des groupes faunistiques, les patrons observés apparaissent proches entre les stations avec une dominance, par ordre décroissant, des diptères (57 % des effectifs en D0BIS ; 68 % en D1) suivi par les éphéméroptères (35 % des effectifs en D0BIS ; 24 % en D1). Les autres groupes ne représentent qu’une faible proportion des effectifs (inférieure à 4 % des effectifs totaux de chaque station).
Cette observation est soutenue par la valeur d’indice de similarité de Whittaker-Fairbanks9 calculé
entre les deux peuplements globaux des deux stations : une valeur de 72 % de similarité est obtenue. L’indice de Whittaker-Fairbanks, identique à l’indice de Schoener mais antérieur, donne la mesure de la ressemblance entre les structures d’effectifs des populations composant les peuplements dans chacune des situations considérées. Fondé sur les proportions des effectifs, il est peu sensible aux effectifs les plus faibles dans les peuplements. On considère que ce type d’indice supérieur à 60% (ou 0,6 selon l’expression calculée) traduit une bonne ressemblance des structures d’effectifs. Les peuplements des stations D0BIS et D1 peuvent donc être qualifiés de similaires dans leur structure d’effectifs traduisant ainsi en partie la proximité géographique des stations d’étude et donc la ressemblance de structure faunistique.
En agrégeant les données par station, la Figure 11 présente graphiquement la décomposition des richesses taxonomiques et des abondances par groupe faunistique. Ces groupes sont composés de niveaux taxonomiques différents : ordre pour les insectes (plécoptères, éphéméroptères, trichoptères, coléoptères, odonates, diptères), sous-embranchement (crustacés), classe (turbellariés, gastéropodes) et sous-classe (oligochètes).
9 Whittaker R.H. and Fairbanks C.W., 1958. A study of plankton copepod communities in the Columbia basin,
27
Figure 11 : Richesses taxonomiques (figure de gauche) et effectifs bruts (figure de droite) des peuplements agrégés par station d’étude et décomposition par groupes faunistiques.
Une partie des résultats présentés dans le Tableau 6 sont représentés graphiquement dans la Figure 11. En observant les données des quatre campagnes d’échantillonnage effectuées en 2014 et en les agrégeant par station, les principaux résultats de comparaison inter-stationnelle des peuplements de macroinvertébrés sont :
- Les peuplements de D0BIS sont nettement moins denses que ceux de D1,
- Les peuplements de D0BIS apparaissent moins diversifiés taxonomiquement,
- Les effectifs importants en D1 sont principalement en lien avec des taxons appartenant aux
ordres des diptères, des éphéméroptères et dans une moindre mesure des trichoptères,
- En excluant les taxons rares (représentés par un seul individu), les valeurs de richesses sont
comparables entre les deux stations,
- Les structures de distribution des effectifs au sein des différents taxons apparaissent
comparables au sein des peuplements des deux stations d’étude.
En conclusion, les peuplements des deux stations apparaissent proches au regard de la composition taxonomique et des structures de distribution des effectifs, similarité certainement en lien avec la proximité géographique des sites.
La présence de taxons rares dans la station D1 permet le maintien de richesses plus fortes qu’en D0BIS.
En parallèle, les densités supérieures en D1 pourraient traduire des contextes locaux trophiques différents, ou bien une situation géomorphologique singulière dans la vallée alluviale. ces différences sont observées de façon récurrente depuis 2005.
28
3.1.4 Structures faunistiques observées dans les trois ambiances d’écoulement
La Figure 12 présente les valeurs de richesses taxonomiques et d’effectifs bruts des listes faunistiques agrégées par ambiance d’écoulement et par station d’étude avec la décomposition par principaux groupes faunistiques.
Figure 12 : Richesses taxonomiques (figure de gauche) et effectifs bruts (figure de droite) des peuplements agrégés par ambiance d’écoulement et par station d’étude et décomposition par groupes faunistiques.
A partir des richesses globales par ambiance et par station (Figure 12 à gauche), les valeurs observées varient peu entre les ambiances au sein d’une station donnée. La composition des richesses des principaux groupes faunistiques présentés est globalement semblable entre les ambiances.
Pour les deux stations, les effectifs globaux par ambiance (Figure 12 à droite) augmentent depuis l’ambiance d’écoulements les plus lents jusqu’à la plus rapide. En D0BIS, cette augmentation est surtout portée par les individus appartenant aux ordres des éphéméroptères et des diptères. En D1, le patron apparait plus complexe puisque ce sont les éphéméroptères qui apparaissent plus nombreux dans l’ambiance A2 comparativement à l’ambiance A1 ; puis le nombre de diptères est maximal dans l’ambiance A3 (4340 individus diptères l’ambiance A3 ; contre seulement 2728 et 2833 individus de diptères respectivement dans les ambiances A1 et A2). Les prises en considération des stades larvaires et des données relatives au périphyton permettront certainement de nuancer ces observations.
3.1.4.1 Richesse et composition faunistique des ambiances d’écoulement
Les deux figures suivantes présentent une comparaison par ambiance des mêmes métriques biologiques mais en s’intéressant à leurs variabilités au niveau de chaque échantillon. La Figure 13 présente les distributions des valeurs de richesses taxonomiques par échantillon en fonction des ambiances d’écoulement.