HAL Id: hal-02790459
https://hal.inrae.fr/hal-02790459
Submitted on 5 Jun 2020
HAL is a multi-disciplinary open access archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Distributed under a Creative Commons Attribution - NonCommercial - NoDerivatives| 4.0 International License
Développement d’une approche de méta-modélisation du
développement et de la croissance foliaire des graminées
fourragère
Arthur Couturier
To cite this version:
Arthur Couturier. Développement d’une approche de méta-modélisation du développement et de la croissance foliaire des graminées fourragère. Sciences agricoles. 2018. �hal-02790459�
Développement d’une approche de
méta-modélisation du développement
et de la croissance foliaire des
graminées fourragère
Couturier Arthur
2017-2018
Master 2 Mention Biologie Végétale (BV) Parcours Qualité des Productions Spécialisées Mémoire de Fin d’Études
Sous la direction de MM.
Louarn Gaëtan et Combes Didier
Membres du jury Gerhard Buck-Sorlin | Enseignant référent Jérémy Clotault | Représentant du parcours Didier Peltier | Responsable du Master
Soutenu publiquement à Angers le : 17 septembre 2018
L’auteur du présent document vous autorise à le partager, reproduire, distribuer et communiquer selon les conditions suivantes :
- Vous devez le citer en l’attribuant de la manière indiquée par l’auteur (mais pas d’une manière qui suggérerait qu’il approuve votre utilisation de l’œuvre).
- Vous n’avez pas le droit d’utiliser ce document à des fins commerciales. - Vous n’avez pas le droit de le modifier, de le transformer ou de l’adapter.
Consulter la licence creative commons complète en français : http://creativecommons.org/licences/by-nc-nd/2.0/fr/
Ces conditions d’utilisation (attribution, pas d’utilisation commerciale, pas de modification) sont symbolisées par les icônes positionnées en pied de page.
Je tiens tout d’abord à remercier Gaëtan Louarn et Didier Combes pour m’avoir choisi pour ce stage et pour m’avoir encadré tout au long de ces 6 mois. Pour l’ensemble des conseils et des connaissances que vous m’avez apportés.
Merci à toute l’équipe URP3F pour l’accueil et la bonne humeur quotidienne durant ces semaines passées ensemble, et pour les nombreuses discussions à la pause café.
Je tiens à remercier particulièrement Simon Rouet, pour m’avoir montré qu’il y avait plein de chose à faire dans la région poitevine.
Je tiens également à remercier Ela Frak et Abraham Escobar-Gutiérrez pour m’avoir proposé une thèse et ainsi continuer l’aventure dans l’équipe URP3F
Merci à Nabilla Roy pour m’avoir supporté dans le même bureau, même par temps de canicule.
Je tiens à remercier Sébastien Pommies et Jean-Marie Girardeau pour m’avoir dépanné de nombreuse fois pour des problèmes informatiques.
Merci au gardien de nuit de l’INRA pour m’avoir sauvé la vie en retrouvant mes lunettes de vue, que je pensais perdu à jamais.
Merci à tout les stagiaires ayant été présent sur la période de mon stage pour les bons moments passés.
Un grand merci à Hélène Bobille pour m’avoir aidé à trouver ce stage.
Un grand merci à tout les gens du bâtiment stagiaire. Nicolas, Virginie, Marion, Fatemeh, Xiaogang, Marc-Antoine, Anaël et ceux dont le nom m’échappe parce que je suis un piètre colocataire.
Merci à tout mes ami·e·s et à ma famille, qui me regardaient bizarrement après que je leur ait dit que je travaillais sur des plantes virtuelles.
Je tiens également à remercier l’équipe du Dietrich qui m’a occupée de nombreuses soirée et qui m’a permis de découvrir beaucoup d’œuvres que je n’aurais pas vu ailleurs.
R
E
M
E
R
C
IE
M
E
N
T
S
Sommaire
Introduction...5
1 Contexte et état de l’art...5
1.1. Contexte des prairies...5
1.2. Morphogenèse des graminées fourragères...5
1.3. Modèle de plantes... 6 1.3.1. Définitions... 6 1.3.2. Process-based models...7 1.3.3. Modèles structure-fonction...7 1.3.4. Méta-modélisation...8 2 Modèle L-grass... 8 3 Objectifs du stage... 10
Matériel & Méthodes...11
1 Plan d’expérimentation... 11
1.1. Choix des paramètres d’entrée...11
1.2. Choix des variables de sortie...12
1.3. Plan d’expérimentation des simulations pour la construction des méta-modèles...12
2 Construction des méta-modèles...13
2.1. Méta-modèles directs...13
2.2. Méta-modèles en cascade...13
3 Équations des méta-modèles...14
4 Évaluation des méta-modèles...16
4.1. Évaluation par des données indépendantes...16
4.2. Évaluation pour la réponse à la coupe...17
Résultats... 18
1 Résultats des simulations pour le plan de construction des méta-modèles...18
2 Méta-modèles des variables intermédiaires...19
2.1. Évaluation des méta-modèles de la taille des gaines sur des données indépendantes....19
2.2. Évaluation des méta-modèles du temps passé dans la gaine sur des données indépendantes... 20
3 Méta-modèles du phyllochrone...20
3.1. Qualité de l’ajustement des méta-modèles...20
3.2. Évaluation des méta-modèles sur des données indépendantes...21
Couturier Arthur| Développement d’une approche de méta-modélisation du développement et de la croissance foliaire des graminées fourragère 3
3.2.1. Données indépendantes sans coupe...21
3.2.2. Données indépendantes avec coupe...22
4 Méta-modèles de la taille du limbe...22
4.1. Qualité de l’ajustement des méta-modèles de la taille du limbe...22
4.2. Évaluation des méta-modèles de la taille du limbe sur des données indépendantes...23
4.2.1. Données indépendantes sans coupe...23
4.2.2. Données indépendantes avec coupe...24
Discussion...25
1 Résultats des simulations...25
2 Performance des modèles...25
2.1. Méta-modèles des variables intermédiaires...25
2.1.1. Méta-modèles de la taille de la gaine...25
2.1.2. Méta-modèles du temps passé dans la gaine...26
2.2. Méta-modèles du phyllochrone...26
2.3. Méta-modèles de la taille du limbe...27
Conclusion... 29
Bibliographie...30
Annexes...I Couturier Arthur| Développement d’une approche de méta-modélisation du développement et de la croissance foliaire des graminées fourragère 4
Introduction
1 Contexte et état de l’art
1.1.
Contexte des prairies
Les prairies sont des surfaces agricoles cultivées pour leur production de biomasse destinée à l’alimentation des ruminants, dans un but final de production de viande, de lait ou de tout autre produit issu de l’exploitation animale.
Selon l’Agreste, les prairies sont divisées en deux catégories :
– les prairies permanentes, comprenant les prairies naturelles productives, les prairies temporaires semées depuis plus de 6 ans et les prairies peu productives
– les prairies temporaires, composées d’au moins 20 % de graminées semées et ayant données moins de six récoltes.
Malgré cette différenciation principalement d’ordre temporelle, leur composition est similaire car étant composées de plantes fourragères pérennes, qu’elles soient des graminées ou des légumineuses. Cette composition est généralement multi-espèces et détermine la valeur agronomique de la prairie, en termes de rendement et de valeur nutritive.
La valeur agronomique des prairies est déterminée par le rendement et la valeur nutritive de la partie aérienne des plantes fourragères, le rendement est principalement déterminé par le nombre de feuilles ainsi que de leur taille (Lafarge and Durand, 2011).
Du fait de leur mode de culture, les prairies sont sujettes à de nombreuses coupes qui ont pour effet de modifier le couvert végétal et donc les interactions inter-espèces, notamment au niveau de la compétition pour la lumière. Ces interactions inter-espèces ont pour effet de modifier le couvert végétal et donc la valeur agronomique des prairies.
Les graminées sont les espèces végétales principales des prairies, l’étude de leur morphogenèse incluant les interactions inter-espèces apparaît donc comme importante afin de comprendre le fonctionnement des prairies.
1.2.
Morphogenèse des graminées fourragères
La morphogenèse correspond à « l’ensemble des processus qui aboutissent à la mise en forme d’un végétal ou d’un organe » (Lafarge and Durand, 2011).
Chez les graminées, la talle est le point central de la morphogenèse. Une talle de graminée est constituée de différentes couches de feuilles enroulées les unes sur les autres. Les feuilles de Couturier Arthur| Développement d’une approche de méta-modélisation du développement et de la croissance foliaire des graminées fourragère 5
Figure 1 : Schéma des processus d’auto-régulation des graminées. Repris de (Verdenal, 2009)
graminées sont constituées de 3 parties distinctes : le limbe, la gaine et la ligule, cette dernière faisant office de jonction entre les 2 autres parties.
La morphogenèse des graminées peut être décrite par l’hypothèse d’auto-régulation (Verdenal et al., 2008). Cette hypothèse décrit la morphogenèse des graminées au travers d’une boucle de contrôle entre les différentes parties des feuilles déclenchant mutuellement leur croissance et déterminant leur taille (fig.1) :
– La longueur des limbes est régulée par la longueur de la gaine des feuilles précédentes, car les feuilles doivent a minima avoir une taille égale à la taille de la gaine afin de recevoir de la lumière.
– Le déclenchement de la croissance d’une feuille est régulé par l’émergence du limbe de la feuille précédente.
– Le déclenchement du tallage est régulé par l’ontogenèse des feuilles, et peut également être réprimé par l’environnement lumineux lorsque le ratio R:FR (R = rouge clair, FR = rouge sombre) dépasse un seuil dépendant du génotype de la plante.
– En cas de défoliation, l’ensemble de cette boucle de développement est modifiée puisqu’un coupe provoque une modification de la taille des gaines.
La taille des feuilles et leur nombre, qui permettent de décrire le rendement de la prairie, sont donc déterminés par les processus d’auto-régulation des graminées. La description de ces processus par la modélisation permet de tenir compte de l’ensemble des paramètres de morphogenèse des graminées afin de prédire la valeur agronomique des prairies.
1.3.
Modèle de plantes
1.3.1. Définitions
Selon le Larousse, un modèle est défini comme une « représentation schématique d’un processus », mais cette représentation schématique n’est pas une fin en soi puisque celle-ci doit répondre à un ou plusieurs objectifs donnés.
Dans le cadre de la recherche scientifique, un modèle est défini comme une représentation simplifiée et communicable de la réalité, afin d’étudier un phénomène même si celui-ci est partiellement reproduit (Legay, 1997).
On peut dénombrer 5 principaux objectifs à la modélisation, chaque modèle a pour but de répondre à un ou plusieurs de ces points :
Couturier Arthur| Développement d’une approche de méta-modélisation du développement et de la croissance foliaire des graminées fourragère 6
1. La prévision
2. La structuration des connaissances 3. La gestion des connaissances 4. L’analyse d’hypothèses 5. L’exploration des possibles
La prévision a pendant longtemps été l’objectif de la modélisation, poussée par une volonté d’améliorer les rendements des productions agricoles. Ainsi, avant même l’apparition de la modélisation assistée par ordinateurs, les chercheurs et les chercheuses ont tenté·e·s de créer des modèles capables de représenter au mieux la réalité afin de prévoir de manière précise la réponse du rendement face à différentes conditions (Vos et al., 2007). Avec l’apparition de technologie permettant de réaliser des calculs toujours plus complexe, les méthodes de modélisation ont pu être améliorés pour se rapprocher au maximum de la réalité. De plus, différents types de modèles ont vu le jour afin de répondre à différents objectifs.
Dans le domaine de la biologie végétale, 2 types de modèles mécanistes sont majoritairement utilisés pour atteindre à ces objectifs : les « Process-based models » (PBM) et les modèles structure-fonction (FSPM) (Vos et al., 2007). Ces types de modèles font opposition aux modèles empiriques, qui ne tiennent pas compte des processus de développement.
1.3.2. Process-based models
Les « Process-based models » (PBM) ont principalement été utilisés pour modéliser la relation entre le rendement d’une culture et ses conditions environnementales. Dans ces modèles l’unité de référence est la parcelle. Ces modèles ont pour but de représenter les principaux processus physiologiques mis en jeu sur un couvert végétal de façon mécaniste et déterministe, avec par exemple les travaux de Thornley (1998) sur la photosynthèse.
1.3.3. Modèles structure-fonction
Les modèles structure-fonction (FSPM) ont pour rôle de décrire des plantes dans un espace tri-dimensionnel à des échelles bien plus précises, pouvant aller jusqu’à la description de chaque organe distinct (Evers et al., 2018). Ce type de modèle a pour rôle de tenir compte des processus physiologiques associés à l’architecture de la plante, ainsi la représentation de la plante dans l’espace est intrinsèquement liée aux processus décrit dans le modèle. De plus, la représentation spatiale des plantes permet l’intégration des compétitions entre les individus Couturier Arthur| Développement d’une approche de méta-modélisation du développement et de la croissance foliaire des graminées fourragère 7
pour les différentes ressources environnementales (nutriments, lumière…) dans l’espace et dans le temps.
Mais cette représentation réaliste vient avec un coût : en effet, le temps de calcul exigé pour l’ensemble des fonctions intégrés aux modèles est très supérieure au temps de calcul généralement exigé par les autres types de modèle.
C’est ce type de modèlisation qui est utilisé par le modèle L-grass.
1.3.4. Méta-modélisation
Un méta-modèles est un « modèle d’un modèle » (Kleijnen, 1987), construit à partir des sorties d’un modèle préexistant. Contrairement aux PBM et aux FSPM, les méta-modèles sont des modèles empiriques qui permettent une approximation des sorties du modèle d’intérêt tout en diminuant fortement le temps de calcul nécessaire (Jin et al., 2001).
Le coût énergétique et temporel nécessaire à l’utilisation en simulation des modèles FSPM est exponentiel, l’utilisation de méta-modèles afin de réduire le temps de calcul semble pertinente. Les méta-modèles faciliteraient également l’usage des modèles, notamment sur des ordinateurs moins puissants qui ne sont pas prévus pour la modélisation.
Parmi les principales techniques de méta-modélisation figurent les régressions polynomiales (Jin et al., 2001), les surfaces de réponses (Hammadi et al., 2012), les régressions locales, les régressions multiples adaptatives ou les approches par réseaux de neurones.
Les régressions polynomiales permettent d’établir une relation entre une variable dépendante et une variable indépendante. Les surfaces de réponses font la relation entre variables explicatives et les variables de réponse, afin d’obtenir une réponse optimale. Les régressions locales permettent une régression non paramétrique combinant des régressions linéaires avec des régressions non linéaires.
2 Modèle L-grass
Le modèle L-grass est un modèle structure-fonction des graminées fourragères est développé à l’INRA depuis 2005. Il a d’abord été capable de simuler les processus d’auto-régulation de la morphogenèse aérienne des graminées fourragères d’une prairie en prenant en compte les conditions de lumière et les conditions de coupe (Verdenal, 2009) ; il a par la suite été amélioré afin d’intégrer la morphogenèse racinaire (Migault, 2015).
Couturier Arthur| Développement d’une approche de méta-modélisation du développement et de la croissance foliaire des graminées fourragère 8
Figure 2 : Réponse du modèle L-grass à la variation du potentiel de morphogenèse C. Repris de (Verdenal, 2009)
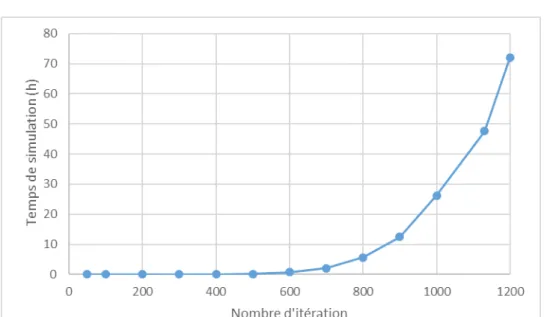
Figure 3: Evolution du temps nécessaire aux simulation en fonction du nombre d’itérations, à une densité de 100 plantes/m², un LAI critique de 1 et un potentiel de morphogenèse C de 0,5.
Le nom « L-grass » provient du mode de représentation des plantes dans ce modèle, car il s’appuie sur les L-systèmes. Les L-systèmes, développés par Aristid Lindenmayer (1968) et basés sur les travaux de linguistique de Noam Chomsky (1956), sont un système grammatical dont les éléments sont réécrits à chaque pas de temps, des objets présents en suivant des règles de productions précises. Dans le cas du modèle L-grass, l’ensemble des règles de réécritures sont disponibles en Annexe I.
L’auto-régulation de la morphogenèse aérienne est simulée à partir de 3 règles :
1. Un phytomère est produit lorsque la feuille du phytomère précédent émerge de la pseudotige.
2. La longueur finale d’une feuille et de sa gaine dépend de la période de temps écoulé entre l’apparition du phytomère et l’émergence de la feuille.
3. Si les conditions environnementales lumineuses sont adéquates lors de l’émergence de la feuille, le bourgeon axillaire du phytomère est différencié en méristème apical. Sinon le bourgeon est avorté.
Un des principaux intérêts de L-grass réside dans ses propriétés émergentes. En effet, contrairement à des modèles qui nécessitent un grand nombre de paramètres d’entrée, L-grass est capable de générer un grand nombre de sorties qui décrivent la morphogenèse des graminées à partir d’un nombre réduit de paramètres d’entrée.
Malgré un faible nombre de paramètre d’entrée, L-grass est capable de simuler une large gamme d’espèces de plantes fourragères, notamment par le paramètre de potentiel de morphogenèse C dont les variations provoque une modification du nombre de feuille et de leur longueur (fig.2).
L-grass est également capable d’anticiper les modifications provoquées par la coupe avec seulement 2 paramètres d’entrée correspondants à la hauteur et à la date de coupe.
Les sorties du modèle décrivent la vitesse de croissance, avec le nombre de feuilles et le nombre de talles produits, et décrivent les propriétés des feuilles, avec la taille du limbe et de la gaine ainsi que le temps passé dans la gaine.
Couturier Arthur| Développement d’une approche de méta-modélisation du développement et de la croissance foliaire des graminées fourragère 9
L-grass permet de produire un grand nombre de sortie mais son temps de simulation suit un profil exponentiel en fonction du nombre d’itération demandé (fig.3). L’utilisation de la méta-modélisation permettrait de contourner ce problème.
Malgré la capacité de L-grass à simuler la partie racinaire des graminées, la méta-modélisation effectuée au cours de ce stage se focalisera sur la partie aérienne puisque c’est cette partie des plantes qui décrit la valeur agronomique des prairies.
3 Objectifs du stage
Un des principaux problèmes actuel de L-grass porte sur le temps de calcul nécessaire pour réaliser des simulations sur une longue période (1 an et plus). L’objectif principal de ce stage est donc de construire des méta-modèles capables de prédire les sorties du modèle qui caractérisent la valeur agronomique des prairies sous différentes conditions environnementales (réponses à la coupe et à la lumière) et pour une large gamme d’espèces de graminées. Ces méta-modèles auront pour but afin de réduire le temps de calcul nécessaire pour les simulations tout en conservant les informations produites par L-grass, et ainsi permettre une optimisation de la conception de prairies multi-espèces à l’aide de méta-modèles capable de simuler rapidement une large gamme d’espèces et de conditions de culture.
Pour simuler une large gamme d’espèces graminées, une série de simulations est effectuée. Différents méta-modèles sont construits pour chaque paramètre de sortie et sont ensuite comparés et évalués entre-eux et par rapport aux sorties de L-grass.
Couturier Arthur| Développement d’une approche de méta-modélisation du développement et de la croissance foliaire des graminées fourragère 10
Matériel & Méthodes
1 Plan d’expérimentation
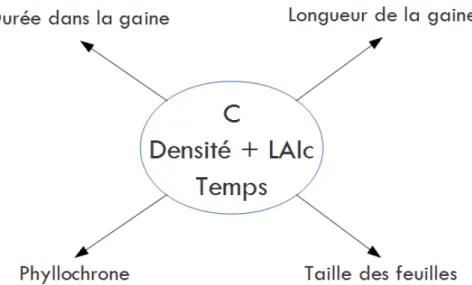
Dans ce contexte, le principal objectif de la méta-modélisation est de prédire différentes variables de sorties qui décrivent la morphogenèse de la partie aérienne des graminées. Pour cela il faut au préalable choisir les paramètres d’entrée qui seront testées à différentes valeurs ainsi que les variables de sortie.
1.1.
Choix des paramètres d’entrée
La valeur agronomique des prairies dépend principalement de sa composition botanique, il est important de choisir des paramètres qui permettent de simuler diverses variétés d’espèces fourragères afin de couvrir un large spectre de plantes pouvant être utilisées en association avec des légumineuses.
Comme décrit précédemment, le paramètre C est un facteur multiplicatif du potentiel de croissance, dont la variation permet de simuler différentes espèces de graminée. C’est donc un paramètre d’entrée important dans la construction de L-grass.
La compétition pour la lumière peut être évaluée à l’aide du LAI (Leaf Area Index ou Indice Foliaire), un paramètre qui exprime la surface foliaire du couvert par unité de surface de sol (m²/m²). Le LAI critique est défini par Brougham (1958) comme la quantité de surface foliaire nécessaire pour intercepter 95 % de la lumière incidente, dans ce rapport le LAI critique sera défini comme le seuil à partir duquel le tallage est ralenti. Le LAI critique est un paramètre qui dépend du génotype de la plante.
La densité de semis, exprimé en nombre de plantes par m², est un paramètre qui caractérise la distance entre chaque plante, une variation de la densité entraîne donc une variation du LAI local. C’est donc un second paramètre d’entrée qui rend compte de la compétition pour la lumière.
Les paramètres d’entrée choisis sont donc le C, le LAI critique (LAIc) et la densité (D) de semis.
Couturier Arthur| Développement d’une approche de méta-modélisation du développement et de la croissance foliaire des graminées fourragère 11
1.2.
Choix des variables de sortie
Les variables de sorties choisies doivent rendre compte de l’auto-régulation du Ray-grass.
La taille finale du limbe a été montrée comme dépendante de la taille de la gaine (Wilson and Laidlaw, 1985), ces 2 variables de sorties sont donc corrélées. Récupérer ces variables permettent donc de rendre compte d’une partie des processus d’auto-régulation du Ray-grass. En plus de la taille de la gaine, le temps passé par la feuille dans la gaine influence la longueur finale des limbes (Verdenal, 2009), la récupération de cette variable pourrait permettre une meilleure prédiction de la taille des limbes, couplée à la longueur de la gaine.
Comme décrit par Lafarge et al. (2005), la vitesse de tallage suit une boucle de rétro contrôle où une talle fille apparaît au niveau d’une feuille n lorsque la talle mère émet sa feuille n+3. Le tallage est également influencé par l’environnement lumineux, ce paramètre de sortie paraît pertinent pour caractériser l’auto-régulation du Ray-grass et les interactions inter-espèces vis-à-vis de la lumière.
Le phyllochrone détermine la vitesse de croissance des feuilles d’une talle, il est décrit comme régulier au cours du temps sur chaque talle d’un individu, mais avec des variations entre individus (Lafarge and Durand, 2011). Le phyllochrone est également coordonné par l’émergence des feuilles (Verdenal, 2009), une coupe au niveau des gaines peut donc modifier la date d’émergence d’une feuille, ce qui entraîne une modification du phyllochrone.
1.3.
Plan d’expérimentation des simulations pour la construction
des méta-modèles
Les simulations sont faites sur 1500 itérations, ce qui correspond dans le modèle L - grass à 1500 °J, et sur un couvert de 9 plantes. L-grass est capable de simuler un couvert de taille infini, les plantes simulées ne subissent donc pas d’effet de bordure.
Des gammes de valeurs sont choisies pour chacun des paramètres d’entrée :
• Le paramètre C est décrit par Verdenal (2009) comme un potentiel morphogénétique qui a un effet sur de multiples variables de sortie (longueur finale des feuilles, surface foliaire, vitesse de tallage…). Le choix des valeurs de C a été fait dans une gamme légèrement plus large que celle utilisée dans la thèse : les 10 valeurs utilisées vont de 0.5 à 2.75, avec un pas régulier de 0.25.
Couturier Arthur| Développement d’une approche de méta-modélisation du développement et de la croissance foliaire des graminées fourragère 12
Figure 5 : Schéma de construction de méta-modèles en cascade Figure 4 : Schéma de construction de méta-modèles de type direct
• La densité d’un semis prairial est généralement de 1000 plantes par m², le choix a été de prendre des valeurs de densité supérieures et inférieures à cette valeur afin de couvrir un large spectre de scenarii. 9 densités ont été testées : 100, 200, 400, 600, 800, 1111, 1300, 1500 et 2000 plantes/m².
• Le LAI critique du Ray-grass est de 3 (Simon and Lemaire, 1987). La gamme de valeur pour ce paramètre n’est pas supérieure à cette valeur afin d’observer l’influence de ce paramètre sur le développement des plantes, les résultats pour des LAI supérieurs forment un plateau et ont des résultats semblables. La gamme pour le LAI critique est donc : 1, 1.5, 2, 2.5, 3.
Chaque scénario lancé a une valeur de C, de D et de LAIc différente, pour un total de 450 simulations (10 C .
Les simulations sont effectuées à l’aide d’un script batch (Annexe II) permettant le lancement successif de chaque scénario.
2 Construction des méta-modèles
Deux méthodes de méta-modélisations sont réalisées et comparées : une méthode de construction de méta-modèles directe et une méthode de construction en cascade.
Pour des raisons de contraintes temporelles, la construction des méta-modèles aura pour but de prédire le phyllochrone et la taille des limbes selon ces 2 méthodes.
2.1.
Méta-modèles directs
La première méthode, dite « directe » consiste à construire des méta-modèles capable de prédire chacune des variables de sortie au cours du développement de la graminée, à partir des paramètres d’entrée et du temps (fig.4).
Ce type de construction permet de faire des méta-modèles indépendants les uns des autres, mais ne rendent pas compte des processus d’auto-régulation présent dans L-grass.
Les méta-modèles de type « direct » sont construits pour le phyllochrone, la taille des limbes et la taille des gaines, cette dernière étant nécessaire à la méthode de construction de méta-modèles « en cascade ».
2.2.
Méta-modèles en cascade
La seconde méthode consiste à une construction de méta-modèles en cascade (fig.5).
Le point de départ de cette méthode est le modèle direct de la taille des gaines. Un second méta-modèle permettant de prédire le temps passé dans la gaine est construit partir des tailles Couturier Arthur| Développement d’une approche de méta-modélisation du développement et de la croissance foliaire des graminées fourragère 13
de gaines prédites et des paramètres d’entrée. Enfin les méta-modèles des tailles de limbe et des phyllochrone sont construits en parallèle à partir des temps passés dans la gaine prédits par les précédents méta-modèles et des paramètres d’entrée.
À l’inverse de la méthode directe, ce mode de construction des méta-modèles permet de rendre compte de l’auto-régulation du Ray-grass.
3 Équations des méta-modèles
Dans le modèle L-grass, les variables morphogénétiques sont des propriétés émergentes issues de calculs reproduisant la morphogenèse potentielle ainsi que la réponse à la lumière, supposée indépendante de ce potentiel.
L’équation générale de chaque variable morphogénétique au cours du temps peut alors être écrite de la manière suivante :
Morphogenèsevariable(t)=Potentiel Morphogénétique(t)×Réponseà lalumière
Le potentiel morphogénétique est caractérisé par le paramètre C et la réponse à la lumière par le LAI critique (LAIc) et la densité (D), cette équation peut donc être détaillée pour l’ensemble des variables d’intérêt.
Le profil des équations des méta-modèles directs en fonction du temps (t) est le suivant :
Phyllochrone(t ,C , LAIc , D)=f (t ,C)×f (LAIc)×f (D)
Taille du limbe(t ,C , LAIc , D)=f (t ,C)×f (LAIc)×f (D)
Taille de la gaine(t ,C , LAIc , D)=f (t ,C)×f (LAIc)×f (D)
Temps dans la gaine(t ,C , LAIc , D)=f (t ,C)×f (LAIc)×f (D)
Vitesse de tallage(t ,C , LAIc , D)=f (t ,C)×f (LAIc)×f (D) De la même manière, le profil des équations des méta-modèles en cascade est :
Le temps (t) correspond au temps phyllochronique, ce temps est caractérisé par le rang sur lequel se situe la feuille observée.
On s’attend à ne pas trouver d’effet de la densité et du LAIc sur la taille du limbe et la taille de la gaine puisque ces variables ne sont pas couplés au rayonnement dans la construction de L - grass.
Couturier Arthur| Développement d’une approche de méta-modélisation du développement et de la croissance foliaire des graminées fourragère 14
Temps dans la gaine(t ,Taillede la gaine ,C , LAIc , D)=Tailledelagaine(t ,C , LAIc , D)×f (C)×f (LAIc)×f (D)
Phyllochrone(t ,Temps dansla gaine ,C , LAIc, D)=TempsDansLaGaine(t ,C , LAIc , D)×f (C)×f (LAIc)×f (D)
Taille du limbe(t ,Temps dansla gaine ,C , LAIc , D)=TempsDansLaGaine(t ,C , LAIc , D)×f (C)×f (LAIc)×f (D)
On s’attend également à un phyllochrone régulier en fonction du temps phyllochronique pour chaque simulation.
Afin d’obtenir des méta-modèles précis, différentes méthodes de méta-modélisations ont été utilisées :
a) Un modèle polynomial emboîté, les coefficients de chaque paramètre suivant eux-mêmes un polynôme en fonction d’un second paramètre.
Par exemple, un modèle de degré 4 en fonction du rang dont chaque coefficient suis un polynôme de degré 3 en fonction de C permet de créer un méta-modèle de degré 16 :
f(rang ,C)=(a1.C3+a2.C2+a3.C+a4).rang4 +(b1.C3+b2.C2+b3.C+b4). rang3 +(c1.C 3 +c2.C 2 +c3.C+c4).rang 2 +(d1.C 3 +d2.C 2 +d3.C+d4).rang +(e1.C3 +e2.C2 +e3.C+e4)
Les modèles polynomiaux sont construits à l’aide de la fonction « nls » du package Rstudio « stats ».
b) Un modèle de surface de réponse (RSM), réalisé à l’aide du package Rstudio « RSM », et suivant l’équation suivante :
f(rang ,C)=a + b. Rang + c.C + d . Rang .C + e . Rang2 +f .C2
Les modèles polynomiaux sont construits à l’aide de la fonction « rsm » du package Rstudio « rsm ».
c) Un modèle de régression locale (LOESS), contrairement aux précédents modèles, le méta-modèle de régression locale ne repose pas sur une équation mathématique mais sur un tableau de valeurs prédites à partir des valeurs observées.
Le modèle « Brain-Cousens hormesis » utilisé pour le modèle « modelLimbe1.2 »à la formule suivante :
f(rang)=c+ d−c+f . rang
1+eb(ln (rang)−ln(e))
Le choix de cette formule est due à la capacité de cette équation à suivre une diminution après avoir atteint l’asymptote de la sigmoïde, à l’inverse d’autres courbes donc la valeur reste figée après avoir atteint un plateau.
Couturier Arthur| Développement d’une approche de méta-modélisation du développement et de la croissance foliaire des graminées fourragère 15
Tableau 1: Nomenclature utilisé pour les méta-modèles construit à partir des données d’ajustement produites par L-grass.
Tableau 1: Nomenclature utilisé pour les méta-modèles construit à partir des données d’ajustement produites par L-grass.
Variable Type de méta-modèle Méthode de construction Nom attribué
Temps passé dans la gaine cascade ModeleTDG2.1 Temps passé dans la gaine cascade polynome du 2nd degré ModeleTDG2.2 Temps passé dans la gaine cascade LOESS ModeleTDG2.3 Taille de la gaine directe polynome ModeleGaine1.1 Taille de la gaine directe RSM ModeleGaine1.2 Taille de la gaine directe LOESS ModeleGaine1.3 Taille du limbe directe polynome ModeleLimbe1.1 Taille du limbe directe Brain-Cousens ModeleLimbe1.2 Taille du limbe directe RSM ModeleLimbe1.3 Taille du limbe directe LOESS ModeleLimbe1.4 Taille du limbe cascade polynome du 1er degré ModeleLimbe2.1 Taille du limbe cascade ModeleLimbe2.2 Taille du limbe cascade LOESS ModeleLimbe2.3 Phyllochrone directe polynome ModelePhyllochrone1.1 Phyllochrone directe RSM ModelePhyllochrone1.2 Phyllochrone directe LOESS ModelePhyllochrone1.3 Phyllochrone cascade polynome ModelePhyllochrone2.1 Phyllochrone cascade RSM ModelePhyllochrone2.2 Phyllochrone cascade LOESS ModelePhyllochrone2.3
polynome du 1er degré
polynome du 4e degré
Tableau 2
n° simulaton Nombre d’itératon nombre de plantes Densité C LAI critque Date de coupe (°J) Hauteur de coupe (mm)
1 1500 9 1111 0,54 0,8 1000 30 2 1500 9 1111 0,92 0,8 1000 30 3 1500 9 1111 1,04 0,8 1000 30 4 1500 9 1111 1,31 0,8 1000 30 5 1500 9 1111 1,39 0,8 1000 30 6 1500 9 1111 2,05 0,8 1000 30 7 1500 9 1111 2,24 0,8 1000 30 8 1500 9 1111 2,39 0,8 1000 30 9 1500 9 1111 2,44 0,8 1000 30 10 1500 9 1111 2,45 0,8 1000 30
Tableau 2 : Scenarii des simulations d’évaluation des méta-modèles. Les parametres de coupe ne sont pas utilisés pour les simulations sans coupe
La nomenclature des modèles, selon leur type de construction et leur équation, est répertoriée dans le tableau 1.
4 Évaluation des méta-modèles
4.1.
Évaluation par des données indépendantes
La première méthode de validation des méta-modèles consiste à simuler les variables de sorties par L-grass et par les méta-modèles à partir de valeurs de paramètres d’entrée n’ayant pas été utilisées pour la construction des méta-modèles. Ces valeurs de C, de densité et de LAI critique sont tirés aléatoirement dans la gamme de chaque paramètre. Le temps en degrés-jour et le nombre de plantes reste identique aux simulations d’origine, de plus les paramètres d’entrée n’ayant pas d’effet sur la variable étudiée sont fixées à partir des gammes utilisées pour les ajustements. Les scenarii de simulation de validation sont dans le tableau 2.
Une fois les simulations sur ces valeurs effectuées une comparaison est effectuée entre les sorties de L-grass et les sorties des méta-modèles. Par abus de langage, les sorties de L-grass sont également désignées comme étant des valeurs « observées ».
Plusieurs critères statistiques d’évaluation des modèles sont utilisés :
• Le critère AIC est un critère calculé sur les modèles paramétriques et permet d’établir un classement entre les différents modèles, par ordre croissant des valeurs de AIC, la valeur la plus faible est donc la mieux classée.
• La RMSE (Root Mean Square Error) permet d’évaluer la magnitude de l’erreur des modèles (Coucheney et al., 2015). Ce critère à la même unité que la variable étudiée :
RMSE=
√
1n
∑
i=1n
(Si−Oi)
2
Avec O les valeurs observées et S les valeurs simulées par le méta-modèle
• La RMSE relative (rRMSE), correspondant à la RMSE divisée par la moyenne des valeurs observées. Ce critère est sans unité.
RMSE=100RMSE
O
Les seuils permettant de déterminer si les modèles prédisent correctement les variables étudiées sont choisis arbitrairement : un modèle ayant une RMSE relative inférieure à 30 % sont considérés comme ayant une bonne prédiction, car la totalité de la dispersion Couturier Arthur| Développement d’une approche de méta-modélisation du développement et de la
est inférieure à la moitié de la moyenne de la variable étudiée ; un second seuil est fixé à 50 %, les modèles ayant une rRMSE comprise entre 30 % et 50 % sont considérés comme ayant une prédiction correcte et les modèles dont la rRMSE est supérieure à 50 % sont considérés comme des modèles ne prédisant pas correctement la variable étudiée.
• La RMSE peut être divisée en 2 critères permettant d’identifier le principal type d’erreur du modèle (Willmott, 1981) : Le biais (RMSE systématique, ou RMSEs) et la dispersion( RMSE systématique, ou RMSEu). Ces critères ont la même unité que la variable étudiée et sont calculés de la manière suivante :
RMSE=
√
1 n∑
i=1 n ( ¯¯Si−Oi) 2 RMSE =√
1 n∑
i=1 n (Si− ¯¯Si) 2Avec ¯¯S dérivée de la régression linéaire entre les valeurs observées et simulées par le méta-modèle : ¯¯S=a+b×Oi
4.2.
Évaluation pour la réponse à la coupe
À l’instar de la première méthode, le but est de simuler les variables de sortie via L-grass et via les méta-modèles. Les valeurs des paramètres d’entrée choisies sont celles ayant été tirées aléatoirement pour l’évaluation précédente, l’évaluation se faisant par ajout d’un nouveau paramètre d’entrée : la coupe. Pour l’ensemble des scenarii, la coupe sera effectuée à 1000 °J à une hauteur de 30 mm.
La coupe étant un facteur modifiant la taille de la partie aérienne de la plante (gaine et limbe), on s’attend à voir une modification des variables de sortie.
De même que pour les données indépendantes, les modèles sont évalués par les critères statistiques RMSE, rRMSE, RMSEu, RMSEs et AIC.
On s’attend à identifier un biais lié à la coupe pour les méta-modèles direct, qui ne devraient pas être capable d’anticiper la coupe, et une absence de biais pour les méta-modèles en cascade.
Couturier Arthur| Développement d’une approche de méta-modélisation du développement et de la croissance foliaire des graminées fourragère 17
Figure 6 : Courbes d’interaction entre la densité avec le C et entre le LAIc avec le C sur chacune des variables de sorties pour le brin maître des plantes. Les interactions ont été mesurées pour la 5e feuille du brin maître.
Résultats
1 Résultats des simulations pour le plan de construction
des méta-modèles
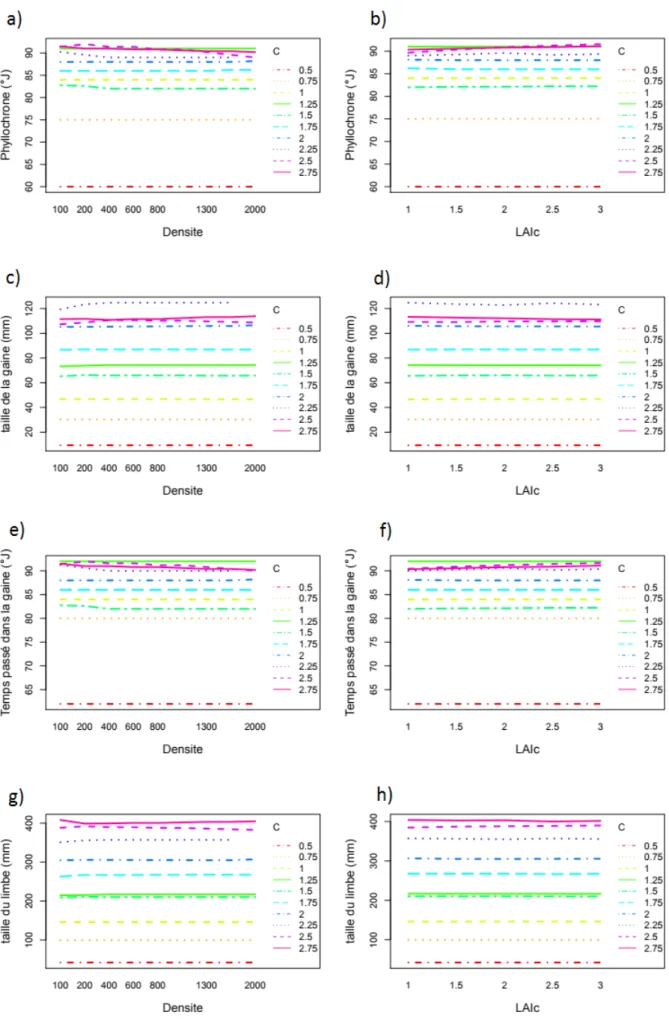
La figure 6 montre les courbes d’interaction entre les paramètres d’entrée par rapport aux variables de sortie. Le phyllochrone, la taille de la gaine, le temps passé dans la gaine et la taille du limbe de la première talle sont très fortement affectés par le paramètre C mais très peu par la densité et le LAI critique.
Pour la 5e feuille du brin maître, le paramètre C fait varier le phyllochrone de 60°J à plus de 90°J (fig.6a,b) ; la taille de la gaine de 9 mm à plus de 120 mm (fig.6c,d) ; le temps passé dans la gaine varie entre 62°J et 92°J (fig.6e,f) et la taille du limbe varie entre 42 mm et 417 mm (fig.6g,h).
La taille des gaines augmente en fonction du rang (fig.7a). Une augmentation du paramètre C provoque une augmentation de la taille de la gaine, mais cette augmentation est moins importante pour des valeurs de C supérieures à 2. Les courbes pour des valeurs de C de 2.5 et 2.75 sont superposées.
Le temps passé dans la gaine augmente au coup du temps phyllochronique ainsi qu’en fonction de C (fig.7b). L’importance de l’augmentation du temps passé dans la gaine lié au paramètre C diminue à partir d’une valeur de C égale à 1.
La taille du limbe en fonction du rang décrit une courbe en cloche (fig.7c) et augmente en fonction de C. Plus la valeur de C est importante, plus la diminution de la taille du limbe se fait précocement.
Comme pour l’évolution du temps passé dans la gaine, le phyllochrone augmente en fonction de C et en fonction du rang phyllochronique (fig.7d), avec une diminution de l’augmentation lié au C à partir de C=1.
Les temps passés dans la gaine en fonction de la taille de la gaine ont un profil logarithmique (fig.7e), avec un plateau proche de 130 °J. Quelques points ont un temps passé dans la gaine élevé pour de faibles tailles de gaine lorsque C est supérieur ou égal à 2,5.
Comme en fonction du rang, les longueurs de limbe en fonction du temps passé dans la gaine suivent une courbe en cloche (fig.7f).
Quels que soient les valeurs de C, le phyllochrone en fonction du temps passé dans la gaine suis une courbe linéaire (fig.7g).
Couturier Arthur| Développement d’une approche de méta-modélisation du développement et de la croissance foliaire des graminées fourragère 18
Figure 7 : Profil des différentes variables d’intérêt par rapport au potentiel morphogénétique C. Les figures a à d sont par rapport au temps phyllochronique, la figure e par rapport à la taille de la gaine et les figures f à g par rapport au temps passé dans la gaine. La couleur des courbes indique leur de potentiel morphogénétique.
2 Méta-modèles des variables intermédiaires
2.1.
Évaluation des méta-modèles de la taille des gaines sur des
données indépendantes
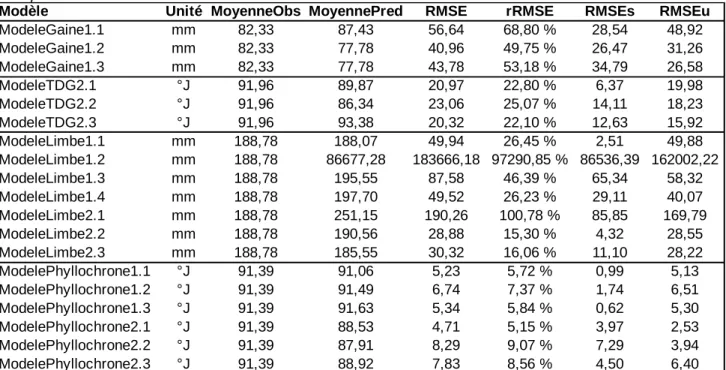
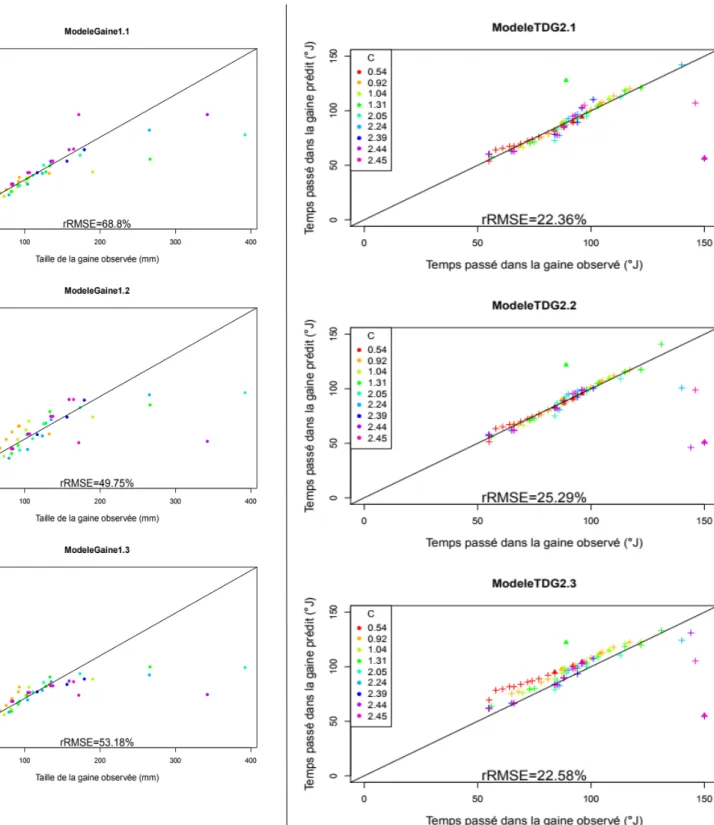
Entre les 2 modèles paramétriques (modèles 1.1 et 1.2), le modèle polynomial 1.1 possède un AIC inférieur au AIC du modèle de surface de réponse 1.2 (27 974 contre 40 843) (Tableau 3). La comparaison des tailles de gaine prédite par rapport aux tailles de gaine observées sur les données de simulation indépendantes (fig.9) montre des profils semblable pour les 3 méta-modèles, avec des valeurs prédites proche des valeurs observées pour des tailles de gaines inférieures à 180 mm et des tailles de gaines sous-estimées par les modèles pour des tailles observées supérieures à 180 mm. De plus, pour les 3 méta-modèles, certaines tailles de gaines sont sur-estimés pour des tailles observées proche de 20 mm.
La RMSE relative du modèle 1.1 est la plus élevée des 3 méta-modèles de la taille de la gaine (Tableau 4) avec une valeur de 68,8 %, les modèles 1.2 et 1.3 ont des rRMSE proches de 50 % (respectivement 49,75 % et 53,13 %). La dispersion du modèle 1.1 est 1,7 fois plus importante que son biais ; la dispersion et le biais du modèle 1.2 sont proches, avec une différence de 15 % entre les 2 indices ; le biais du modèle 1.3 est plus grand de 30 % que sa dispersion.
Les 3 modèles ont des courbes de rRMSE partielle superposées pour les rangs 2 à 8 (fig.8). La RMSE relative des rangs 2 et 3 est autour de 35 % puis chute autour de 15 % pour les rangs 4 à 8. La RMSE relative augmente à partir du 9e rang et atteint une valeur maximale au rang 10 pour les modèles 1.2 et 1.3 et au rang 11 pour le modèle 1.1 avant de retourner à des valeurs inférieures à 20 % au rang 13. La rRMSE maximale est de 244 % pour le méta-modèle 1.1, de 76 % pour le méta-modèle 1.2 et de 73 % pour le méta-modèle 1.3.
La construction des méta-modèles de la taille de la gaine est uniquement de type « direct », la comparaison des valeurs prédites par les modèles et les tailles de gaines observées pour des simulations incluant une coupe ne peut pas être réalisée.
Couturier Arthur| Développement d’une approche de méta-modélisation du développement et de la croissance foliaire des graminées fourragère 19
Tableau 3: Classement des méta-modèles paramétriques sur la base du critère AIC
Modèle AIC Classement
ModeleGaine1.1 27974,52 1 ModeleGaine1.2 40843,81 2 ModeleLimbe1.1 42497,12 3 ModeleLimbe1.3 50309,16 4 ModeleLimbe2.1 32636,67 1 ModeleLimbe2.2 40205,53 2 ModelePhyllochrone1.1 26378,40 1 ModelePhyllochrone1.2 29033,85 2 ModelePhyllochrone2.1 36750,62 4 ModelePhyllochrone2.2 34803,64 3 ModeleTDG2.1 21343,77 2 ModeleTDG2.2 17891,13 1
Tableau 4: Principaux critères d’évaluation statistique des méta-modèles sur des données indépendantes.
Modèle Unité MoyenneObs MoyennePred RMSE rRMSE RMSEs RMSEu
ModeleGaine1.1 mm 82,33 87,43 56,64 68,80 % 28,54 48,92 ModeleGaine1.2 mm 82,33 77,78 40,96 49,75 % 26,47 31,26 ModeleGaine1.3 mm 82,33 77,78 43,78 53,18 % 34,79 26,58 ModeleTDG2.1 °J 91,96 89,87 20,97 22,80 % 6,37 19,98 ModeleTDG2.2 °J 91,96 86,34 23,06 25,07 % 14,11 18,23 ModeleTDG2.3 °J 91,96 93,38 20,32 22,10 % 12,63 15,92 ModeleLimbe1.1 mm 188,78 188,07 49,94 26,45 % 2,51 49,88 ModeleLimbe1.2 mm 188,78 86677,28 183666,18 97290,85 % 86536,39 162002,22 ModeleLimbe1.3 mm 188,78 195,55 87,58 46,39 % 65,34 58,32 ModeleLimbe1.4 mm 188,78 197,70 49,52 26,23 % 29,11 40,07 ModeleLimbe2.1 mm 188,78 251,15 190,26 100,78 % 85,85 169,79 ModeleLimbe2.2 mm 188,78 190,56 28,88 15,30 % 4,32 28,55 ModeleLimbe2.3 mm 188,78 185,55 30,32 16,06 % 11,10 28,22 ModelePhyllochrone1.1 °J 91,39 91,06 5,23 5,72 % 0,99 5,13 ModelePhyllochrone1.2 °J 91,39 91,49 6,74 7,37 % 1,74 6,51 ModelePhyllochrone1.3 °J 91,39 91,63 5,34 5,84 % 0,62 5,30 ModelePhyllochrone2.1 °J 91,39 88,53 4,71 5,15 % 3,97 2,53 ModelePhyllochrone2.2 °J 91,39 87,91 8,29 9,07 % 7,29 3,94 ModelePhyllochrone2.3 °J 91,39 88,92 7,83 8,56 % 4,50 6,40
2.2.
Évaluation des méta-modèles du temps passé dans la gaine
sur des données indépendantes
Les méta-modèles du temps passé dans la gaine sont des méta-modèles intermédiaire dont les résultats sont utilisés pour prédire la taille des limbes et le phyllochrone en tenant compte des coupes. Les méta-modèles sont donc uniquement évalués sur des observations de simulations ayant un scénario de coupe.
Entre les 2 modèles paramétriques (modèles 2.1 et 2.2), le modèle 2.1 possède un AIC supérieur au AIC du modèle 2.2 (21 344 contre 17 891) (Tableau 3).
La comparaison des temps passés dans la gaine par les limbes prédits par les modèles avec les temps observés sur les données de simulation indépendantes avec coupe (fig.10) montrent, pour les 3 méta-modèles, la majorité des temps passé dans la gaine prédit sont proches des temps observés, notamment pour des temps observés compris entre 60 °J et 120 °J. Pour des temps passés dans la gaine observés supérieurs à 130 °J, les temps prédits par les 3 méta-modèles en cascade sont sous-estimés par rapport aux temps observés.
Pour les 3 méta-modèles, un temps passé dans la gaine prédit est surestimé par rapport au temps prédit pour un C de 1,31. Ce point correspond au dernier rang du brin, à 1006°J soit 6 itérations après la coupe.
Le modèle 2.1 est le modèle ayant la plus faible RMSE relative (22,36 %) et le modèle 2.2 a la plus élevée (25,29 %).Pour les rangs 2 à 9, les rRMSE partielles des 3 modèles sont inférieures à 20 % (fig.8) puis subissent une augmentation avec une valeur maximum au 11e rang, avec une rRMSE de 50 %. La rRMSE diminue au 12e rang pour les 3 modèles, pour atteindre des valeurs inférieures à 10 %.
3 Méta-modèles du phyllochrone
3.1.
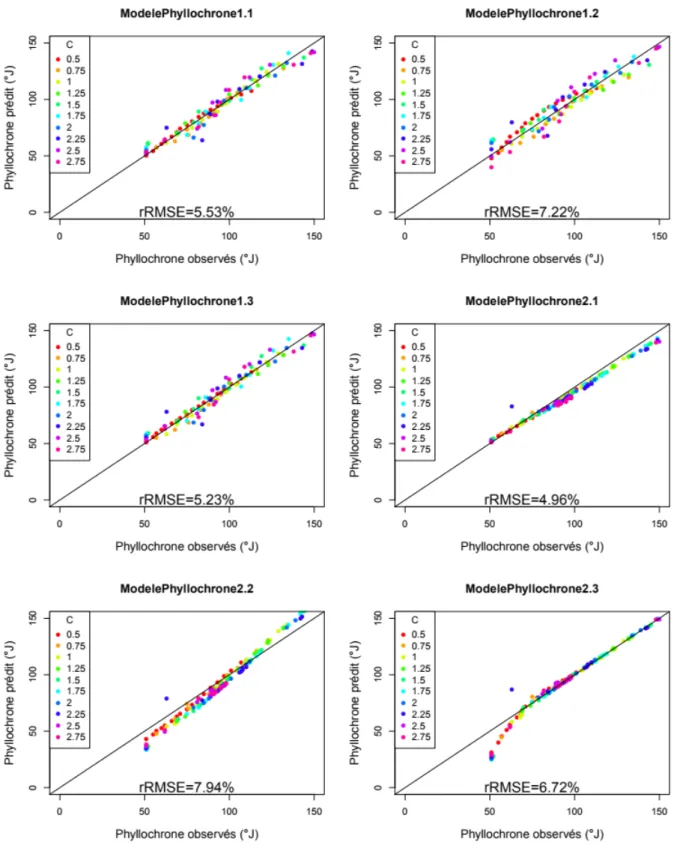
Qualité de l’ajustement des méta-modèles
La comparaison entre les valeurs de phyllochrone prédites par les méta-modèles et les données utilisées pour l’ajustement (fig.11) montrent que les méta-modèles 1.1, 1.2 et 1.3 ont une distribution ayant une distribution homogène. Le modèle 2.1 sous-estime de plus en plus les phyllochrones lorsque la durée des phyllochrones observés augmentent. Le modèle 2.2 prédit des phyllochrone inférieurs à ceux observés sur les données d’ajustement lorsque la valeur Couturier Arthur| Développement d’une approche de méta-modélisation du développement et de la croissance foliaire des graminées fourragère 20
Figure 9: Comparaison entre les tailles de gaines observées sur les données indépendantes et les tailles de gaines prédites par les méta-modèles.
Figure 10: Comparaison entre les temps passés dans la gaine observés sur les données indépendantes avec coupe et ceux prédits par les méta-modèles. La coupe à lieu à 1000 °J à une hauteur de 30 mm.
Les « + » les feuilles dont le limbe a émergé avant la coupe et les « ▲ » indiquent les feuilles dont le limbe a émergé après la coupe.
observée est inférieure à 100 °J puis surestime le phyllochrone pour des valeurs observées supérieures à 120°J. Enfin le modèle 2.3 prédit des phyllochrones très proches de ceux observé lorsque les phyllochrones observés sont supérieurs à 70 °J et sous-estime avant ce seuil.
La RMSE relative des 3 modèles directs est inférieure à 10 % ; parmi ces 3 modèles le modèle 1.3 a la plus faible RMSE (5,62 %) et le modèle 1.2 a la plus élevée (7,52 %). Pour les méta-modèles du phyllochrone de type cascade, le modèle 2.2 à la RMSE relative la plus élevée (7,94 %) et le modèle 2.1 a la rRMSE la plus faible (4,96 %).
Le tableau 3 montre que le modèle 1.1 à la valeur de AIC la plus faible (26 378) parmi les méta-modèles paramétriques du phyllochrone (Tableau 3), suivi du modèle 1.2 et du modèle 2.2 ; le modèle 1.2 possède la valeur de AIC la plus élevée (36 750).
3.2.
Évaluation des méta-modèles sur des données
indépendantes
3.2.1. Données indépendantes sans coupe
Comme pour les comparaisons aux données des simulations, la comparaison des phyllochrones prédits par les méta-modèles par rapport aux données indépendantes (fig.12) montre une distribution homogène des points pour les modèles directs (1.1, 1.2 et 1.3). Pour les méta-modèles en cascade, les phyllochrones prédits par le modèle 2.1 sont sous-estimés pour des phyllochrones observés supérieurs à 80°J ; les phyllochrones prédits par le modèle 2.2 sont sous-estimés pour des valeurs observées inférieures à 100°J et sont sur-estimés pour des phyllochrones observés supérieurs à 100°J. Le modèle 2.3 sous-estime les valeurs de phyllochrone pour des valeurs observées inférieures à 70°J mais possède prédit des phyllochrone très proche de ce qui est observé pour des valeurs supérieures.
Pour les 6 méta-modèles de phyllochrone, la RMSE relative est inférieure à 10 % (Tableau 4). Les 3 méta-modèles direct ont un biais (RMSEs) de 3 à 8 fois inférieure à leur dispersion (RMSEu), les modèles en cascade 2.1 et 2.2 ont des biais supérieurs à la dispersion (respectivement 1,5 et 1,8 fois supérieur) et le modèle 2.3 à une dispersion plus grande que le biais.
Les modèles 2.2 et 2.3 ont des rRMSE partielle maximale sur le deuxième rang (fig.8), respectivement 25 % et 38 %, puis diminuent pour atteindre des rRMSE inférieures à 8 % pour Couturier Arthur| Développement d’une approche de méta-modélisation du développement et de la croissance foliaire des graminées fourragère 21
Figure 11 : Comparaison entre les phyllochrones observées sur les données d’ajustement et celles prédites par les méta-modèles.
le modèle 2.2 et des rRMSE inférieures à 3 % pour le modèle 2.3. Les méta-modèles 1.1, 1.2, 1.3 et 2.1 possèdent une rRMSE inférieure à 10 % pour tous les rangs.
3.2.2. Données indépendantes avec coupe
Les comparaisons entre les phyllochrones prédits par les méta-modèles et les phyllochrones observées sur des données indépendantes ayant subis une coupe (fig.13) montre une surestimation des phyllochrone des feuilles apparues après la coupe pour les 3 modèles directs. Les prédictions des phyllochrones des feuilles apparues avant la coupe sont identiques à celles observées pour les simulations indépendantes sans coupe.
Les comparaisons des méta-modèles en cascade 2.1, 2.2 et 2.3 ont des profils identiques à ceux observés pour les données indépendantes sans coupe (fig.12), les phyllochrones des feuilles apparues après la coupe sont confondues avec les phyllochrones des feuilles apparues avant la coupe.
Les rRMSE des méta-modèles directs ont une augmentation allant de plus de 100 % par rapport aux rRMSE calculée pour les données indépendantes sans coupe (tableau 4) pour le « ModelePhyllochrone1.1 » et « ModelePhyllochrone1.3 » et une augmentation de 47 % pour le « ModelePhyllochrone1.2 ».
La rRMSE des méta-modèles en cascade varie de moins de 5 % pour les 3 modèles par rapport aux rRMSE calculée pour les données sans coupe (tableau 4).
4 Méta-modèles de la taille du limbe
4.1.
Qualité de l’ajustement des méta-modèles de la taille du
limbe
La figure 14 montre les différences entre les tailles des limbes utilisées pour la construction des méta-modèles et les tailles de limbes prédites par les différents modèles. Pour le modèle 1.1, la dispersion des points augmente lorsque la taille des limbes observées augmente, mais la distribution des points est homogène. Le modèle 1.2 présente une dispersion très importante car, contrairement aux autres méta-modèles de la taille du limbe, une grande partie des points n’est pas visible sur la figure 14.
Le modèle 1.3 a une baisse des tailles de limbes prédites par rapport aux valeurs observées pour des tailles de limbes observées supérieure à 250 mm, ainsi que des tailles prédites plus importantes que les tailles observées pour des tailles observées proche de 100 mm.
Couturier Arthur| Développement d’une approche de méta-modélisation du développement et de la croissance foliaire des graminées fourragère 22
Figure 12: Comparaison entre les phyllochrones observés sur les données indépendantes et ceux prédits par les méta-modèles.
Les modèles 2.1, 2.2 et 2.3 ont des tailles de limbe prédites proches des tailles observées. Cependant des points sont fortement surestimés pour des tailles observées de proches de 80 mm, ces points correspondent aux derniers rangs du brin maîtres de plantes ayant une valeur de C supérieures à 2.
Les méta-modèles de taille de limbe 1.1 et 1.4 ont des RMSE relative inférieures au seuil de 30 % et le méta-modèle 1.3 à une RMSE relative inférieur au seuil de 50 %, le modèle 1.2 a une RMSE relative de 73 352 %.
Les 3 méta-modèles en cascade 2.1, 2.2 et 2.3 ont des RMSE relatives comprises entre 150 % et 200 % avec des tailles de limbes prédites nulles pour certaines tailles de limbes observées supérieures à 250 mm.
Les valeurs de AIC pour les modèles paramétriques de la taille des limbes (Tableau 3) montrent que le modèle 2.1 a la valeur la plus faible de 32 363, les modèles 1.1 et 2.2 ont des valeurs supérieures à 40 000 et le modèle 1.3 à la valeur la plus élevée et supérieure à 50 000.
4.2.
Évaluation des méta-modèles de la taille du limbe sur des
données indépendantes
4.2.1. Données indépendantes sans coupe
Le modèle 1.3 prédit des tailles de limbes supérieures aux observées pour des tailles de limbes observées inférieures à 200 mm et prédit des tailles inférieures aux observées pour des tailles observées supérieures à 200 mm (fig.15). Le modèle 1.4 sous-estime les tailles de limbe pour des tailles observées supérieures à 350 mm et sur-estime pour des tailles observées proche de 80 mm. La dispersion des points pour les comparaisons entre les tailles prédites et les tailles observées pour des données indépendantes est homogène pour les modèles 1.1, 2.1, 2.2 et 2.3. Les rRMSE des méta-modèles de limbe 1.1, 1.4, 2.2 et 2.3 sont inférieures au seuil de 30 % et le méta-modèle 1.3 a une rRMSE inférieure au seuil de 50 % (Tableau 4). La RMSE relative du méta-modèle 2.1 est d’environ 100 % et celle du méta-modèle 1.3 est supérieures à 97 000 %. À l’exception du méta-modèle 1.3, les méta-modèles de la taille du limbe ont une dispersion (RMSEu) supérieure au biais (RMSEs). La RMSE relative du modèle 2.1 est la plus élevée des 3 méta-modèles en cascade, mais seulement quelques points ont des estimations très éloignées des valeurs observées, ces points correspondent aux rangs 9 à 12 (fig.8) à des valeurs de C supérieures à 2 (fig.15). Ces points sont également observables chez tous les autres
méta-Couturier Arthur| Développement d’une approche de méta-modélisation du développement et de la croissance foliaire des graminées fourragère 23
Figure 13: Comparaison entre les phyllochrones observés sur les données indépendantes avec coupe et ceux prédits par les méta-modèles. La coupe à lieu à 1000 °J à une hauteur de 30 mm.
Les « + » les feuilles dont le limbe a émergé avant la coupe et les « ▲ » indiquent les feuilles dont le limbe a émergé après la coupe.
modèles de limbe, à l’exception du méta-modèle 1.2, mais leur taille prédite reste plus proche des tailles observées.
Parmi tous les méta-modèles de taille de limbe, les méta-modèles en cascade 2.2 et 2.3 sont ceux ayant la plus faible RMSE relative (proche de 15 %).
La RMSE partielle des méta-modèles de la taille du limbe (fig.8) est, pour tous les modèles à l’exception du 1.2, inférieure à 150 mm sur les 8 premiers rangs du brin maître. Sur ces premiers rangs la RMSE du modèle 1.3 est supérieure à celles des modèles 1.1, 1.4, 2.1, 2.2 et 2.3. Pour l’ensemble les méta-modèles de la taille du limbe, une forte augmentation de la RMSE peut être observée entre les rangs 8 et 10 ; cette augmentation varie de 200 % pour le modèle 2.3 à 10 000 % pour le modèle 2.1. Une diminution de la RMSE à partir du rang 12 est ensuite observable pour l’ensemble des modèles.
4.2.2. Données indépendantes avec coupe
La comparaison des tailles de limbes prédites par les méta-modèles avec les tailles observées sur les données indépendantes avec coupe (fig.16) ont des profils similaires que les comparaisons avec les données indépendantes sans coupe.
Les méta-modèles directs présentent un biais de prédiction des tailles de limbe : les feuilles subissant une coupe ont une taille de limbe surestimée par rapport aux tailles observées. Les méta-modèles 1.3 et 1.4 ont tendance à sous-estimer les tailles de limbes lorsque les limbes observés font plus de 200 mm (fig.15), la sur-estimation des tailles de limbes ayant subis une coupe compense cette sous-estimation et améliore la RMSE relative de ces méta-modèles.
La RMSE relative de tout les méta-modèles de taille de limbe par rapport à des simulations ayant une coupe est du même ordre de grandeur que pour les comparaisons sans coupe, cependant le méta-modèle 1.1 dépasse le seuil de 30 % dans le cas d’une coupe.
À l’inverse des méta-modèles directs, les méta-modèles en cascade ne présentent pas de variation des estimations par rapport aux simulations sans coupe. Les erreurs des méta-modèles sont identiques à celles identifiées sur les précédentes comparaisons.
Les méta-modèles de taille de limbe en cascade 2.2 et 2.3 sont, comme pour les comparaisons sur les simulations indépendantes sans coupe, les méta-modèles ayant les rRMSE les plus faible.
Couturier Arthur| Développement d’une approche de méta-modélisation du développement et de la croissance foliaire des graminées fourragère 24
Figure 14 : Comparaison entre les tailles de limbe observées sur les données d’ajustement et celles prédites par les méta-modèles.
Discussion
1 Résultats des simulations
Les résultats des interactions (fig.6) sont cohérents avec les résultats attendus. L’effet de la lumière sur la croissance des graminées dans L-grass n’a pas d’impact sur le profil des feuilles (taille des limbes et taille des gaines) mais sur le tallage, une talle donnée est donc censé avoir le même profil si le potentiel morphogénétique est identique.
Les profils de longueur de la gaine (fig.7) ne sont pas cohérent avec ce que l’on trouve dans la littérature : lors de la croissance d’une talle, les gaines de chaque feuille doivent être plus grandes que les gaines précédentes, car la ligule doit émerger au-dessus du tube des gaines adultes (Lafarge and Durand, 2011), or les longueurs de gaine pour des C supérieurs ou égal à 2,5 montrent une forte diminution de la taille de la gaine sur leur dernier rang. Cette observation a des conséquences sur les modèles construits, car leur construction tienne compte de la diminution observée et leurs coefficients en sont donc impactés.
Le phyllochrone des graminées est décrit comme régulier dans la littérature (Lafarge and Durand, 2011) et L-grass est construit pour prendre en simuler cette régularité (Verdenal, 2009). Le profil des phyllochrones de L-grass, pour toutes les valeurs de C, augmente au cours du temps, l’accélération de la croissance des limbes pour compenser l’augmentation de la taille des gaines n’est donc pas fonctionnelle sur la version de L-grass utilisée. Cependant les profils observés conservent un profil linéaire, les méta-modèles ayant des bonnes prédictions peuvent donc être facilement réajustés avec de nouvelles données si la raison de cette anomalie est identifiée.
L’objectif de ce stage est de construire des méta-modèles capable de prévoir les sorties de L - grass, si les sorties de celui-ci ne sont pas cohérentes avec les résultats attendus les méta-modèles peuvent alors permettre de mieux comprendre d’où viennent ces anomalies. Ces observations ne sont donc pas gênantes pour répondre à l’objectif.
2 Performance des modèles
2.1.
Méta-modèles des variables intermédiaires
2.1.1. Méta-modèles de la taille de la gaine
Aucun des méta-modèles de la taille de gaine ne passe sous le seuil de RMSE relative de 30 %, et seul le méta-modèle 1.2 est légèrement sous le seuil de 50 %. Les méta-modèles de taille de gaine ne sont donc pas concluant.
Couturier Arthur| Développement d’une approche de méta-modélisation du développement et de la croissance foliaire des graminées fourragère 25
Figure 15: Comparaison entre les tailles de limbe observées sur les données indépendantes et celles prédites par les méta-modèles.
Les rRMSE partielle en fonction du rang permettent d’identifier une mauvaise prédiction des rangs 9 à 11, cette mauvaise prédiction est due aux valeurs de C élevées (supérieures à 2) ayant pour effet de diminuer les tailles des gaines des derniers rangs ; cette diminution entraîne une mauvaise estimation des paramètres du modèle et diminue la précision de la prédiction de la taille de la gaine pour les 3 méta-modèles. Ces mauvaises prédictions sont issues des anomalies identifiées précédemment sur la taille des gaines.
Un ajustement des méta-modèles en excluant les valeurs de C supérieures à 2 pourraient aboutir à une meilleure prédiction de la part de l’ensemble des méta-modèles de la taille de la gaine.
2.1.2. Méta-modèles du temps passé dans la gaine
Les 3 méta-modèles construits pour la prédiction du temps passé dans la gaine sont considérés comme de bons modèles car leur RMSE relative est inférieure au seuil de 30 %.
Ces méta-modèles ont été évalués sur des données indépendantes ayant subis une coupe, les résultats montrent que ces méta-modèles en cascade sont capables de prédire correctement la coupe. Mais le temps passé dans la gaine pour les feuilles qui émergent juste après la coupe est mal estimé.
Malgré la plus faible rRMSE, le « ModeleTDG2.3 » possède un critère de biais plus important que son critère de dispersion, ce résultat permet d’identifier un problème lié à la construction du modèle ayant pour effet de sur-estimer les temps. Parmi les 2 modèles paramétriques, le « ModeleTDG2.2 » est le mieux classé. Ce méta-modèle semble donc être le plus approprié pour prédire le temps passé dans la gaine.
Cependant la fiabilité des méta-modèles du temps passé dans la gaine diminue fortement pour les rangs 9 à 11 des valeurs de C supérieures à 2.
Comme pour les tailles de gaine, un ajustement des méta-modèles des temps passé dans la gaine sur des données dont les valeurs de C sont inférieures à 2 permettrait d’améliorer la précision de ces modèles.
2.2.
Méta-modèles du phyllochrone
L’ensemble des modèles de prédiction du phyllochrone possède des RMSE relatives très inférieurs au seuil de validation, étant tous inférieurs à 10 % pour la comparaison à des données indépendantes sans coupe.
Couturier Arthur| Développement d’une approche de méta-modélisation du développement et de la croissance foliaire des graminées fourragère 26
Figure 16: Comparaison entre les tailles de limbe observées sur les données indépendantes avec coupe et celles prédites par les méta-modèles. La coupe à lieu à 1000 °J à une hauteur de 30 mm.
Les « + » les feuilles dont le limbe a émergé avant la coupe et les « ▲ » indiquent les feuilles dont le limbe a émergé après la coupe.
Pour la réponse à la coupe, les modèles en cascade présentent des résultats très proches de ceux prédits dans coupe tandis que les modèles directs voient leur RMSE relative augmenter. Les modèles en cascades sont donc capables d’anticiper la coupe contrairement aux méta-modèles directs. Les phyllochrones prédits par les méta-modèles directs pour les feuilles apparues après la coupe est sur-estimée.
Le méta-modèle « ModelePhyllochrone2.1 » à la RMSE relative la plus faible des méta-modèles, que ce soit pour la prédiction sur des données sans coupe et sur des données avec coupe. Il est donc le meilleur modèle pour prédire le phyllochrone, quels que soient les paramètres de simulation.
2.3.
Méta-modèles de la taille du limbe
Le « ModeleLimbe1.2 » construit à partir de l’équation de Brain-Cousens à des valeurs de critères d’évaluation très supérieurs aux seuils fixés. La mauvaise prédiction de ce modèle peut s’expliquer par le paramètre « d » de l’équation, qui représente l’asymptote horizontale supérieure. Les variations de ce paramètre pour la paramétrisation du modèle sont très importantes par rapport aux autres paramètres de l’équation ; une mauvaise prédiction de ce paramètre a donc pour effet de modifier significativement l’équation du modèle et provoque une forte variation des prédictions. Cette équation n’est donc pas adaptée pour la méta-modélisation de la taille des limbes.
Les méta-modèles direct « ModeleLimbe1.1 » et « ModeleLimbe1.4 » ont des rRMSE inférieurs au seuil de 30 % et sont donc considérés comme ayant une bonne prédiction. Le « ModeleLimbe1.3 » possède un biais supérieur à sa dispersion, les erreurs de prédiction proviennent donc de la construction du modèle.
Pour les modèles en cascade, le « ModeleLimbe2.1 » possède une rRMSE supérieure à 100 % mais cette forte valeur est principalement liée par une mauvaise estimation des tailles de limbes pour les rangs 9 à 11 (fig.8), ce qui permet de montrer les limites du critère rRMSE, car le modèle est considéré mauvais alors que la grande majorité des points est correctement estimée.
Couturier Arthur| Développement d’une approche de méta-modélisation du développement et de la croissance foliaire des graminées fourragère 27