Master and servant: Regulation of auxin transporters by FKBPs and cyclophilins
10
0
0
Texte intégral
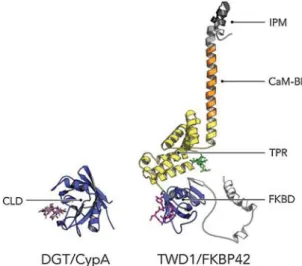
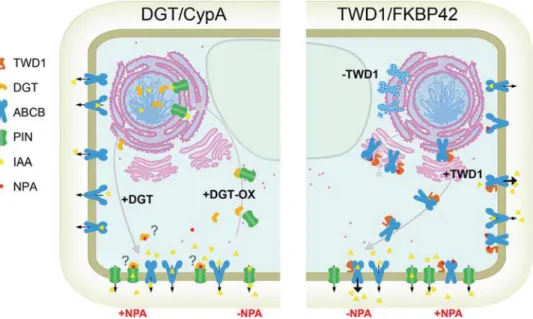
Figure
Documents relatifs