royalsocietypublishing.org/journal/rspb
Review
Cite this article: Carvalho G, Forestier C,
Mathias J-D. 2019 Antibiotic resilience: a
necessary concept to complement antibiotic
resistance? Proc. R. Soc. B 286: 20192408.
http://dx.doi.org/10.1098/rspb.2019.2408
Received: 15 October 2019
Accepted: 7 November 2019
Subject Category:
Evolution
Subject Areas:
health and disease and epidemiology,
microbiology
Keywords:
antibiotic resistance, antibiotic resilience,
bacterial persistence, antibiotic tolerance,
phenotypic heterogeneity, heteroresistance
Author for correspondence:
Jean-Denis Mathias
e-mail: [email protected]
Antibiotic resilience: a necessary concept
to complement antibiotic resistance?
Gabriel Carvalho
1, Christiane Forestier
2and Jean-Denis Mathias
11Université Clermont Auvergne, Irstea, UR LISC, Centre de Clermont-Ferrand, 9 Avenue Blaise Pascal CS 20085,
F-63178, Aubière, France
2Université Clermont Auvergne, CNRS, UMR6023 LMGE, Clermont-Ferrand, France
GC, 0000-0001-6864-5815
Resilience is the capacity of systems to recover their initial state or functions after a disturbance. The concepts of resilience and resistance are comple-mentary in ecology and both represent different aspects of the stability of ecosystems. However, antibiotic resilience is not used in clinical bacteriology whereas antibiotic resistance is a recognized major problem. To join the fields of ecology and clinical bacteriology, we first review the resilience concept from ecology, socio-ecological systems and microbiology where it is widely developed. We then review resilience-related concepts in microbiology, including bacterial tolerance and persistence, phenotypic heterogeneity and collective tolerance and resistance. We discuss how antibiotic resilience could be defined and argue that the use of this concept largely relies on its experimental measure and its clinical relevance. We review indicators in microbiology which could be used to reflect antibiotic resilience and used as valuable indicators to anticipate the capacity of bacteria to recover from antibiotic treatments.
1. Introduction
Bacterial systems are complex and adaptive. When faced with disturbances such as antibiotic treatments, they survive, recover and evolve. The capacity of systems to recover their initial state, some of their functions or composition after a disturbance is called resilience. In ecology, the concepts of resilience and resistance are comp-lementary: while resistance refers to the insensitivity to disturbance, resilience focuses on the recovery of the disturbed system [1,2]. Both resistance and resilience are two different aspects of systems’ stability. However, in clinical bacteriology, antibiotic resilience is not used whereas antibiotic resistance is overwhelmingly studied because multidrug-resistance is currently a major societal challenge [3]. Instead of using resilience, in clinical bacteriology, several other terms have been developed to describe bacterial responses to antibiotic treatments such as tolerance, persistence, phenotypic resistance or recalcitrance. The interest of the scientific community in these mechanisms is gaining importance because they may precede the emergence of antibiotic resistance [4–6].
One reason for the lack of interest in the resilience concept in clinical bacteri-ology is the fact that this concept is often ambiguous. The resilience-based framework, initially developed by Holling [7], suggested numerous concepts and ideas to structure the reactions of systems to disturbances. However, the con-cepts used, mainly resistance and resilience, are vague and potentially conflicting if they are not clearly defined [8,9]. Ecology and socio-ecology have assigned var-ious definitions to resistance and resilience. For instance, resistance can be‘staying essentially unchanged despite the presence of disturbances’ and resilience ‘returning to the reference state (or dynamic) after a temporary disturbance’ [10, p. 325]. Elsewhere, Walkeret al. [11, p. 1] define resilience as ‘the capacity of a system to absorb disturbance and reorganize while undergoing change so as to retain essentially the same function, structure, identity and feedback’. Another paradigm of resilience is the viability-based framework of resilience which focuses on the recovery and maintenance of properties of interest instead
© 2019 The Author(s) Published by the Royal Society. All rights reserved.
of pre-disturbance states [12–14]. In microbial ecology, the focus is usually on the capacity of microbial communities to maintain or recover their pre-disturbance functions and species composition [1]. In clinical bacteriology, Meredithet al. [15] recently developed a framework of antibiotic resistance and resilience and applied it to characterize the response of bac-terial populations expressing β-lactamases and exposed to β-lactam treatments. While the framework of resistance and resilience they propose fits well with their case study, its general application remains questionable.
To develop the concept of antibiotic resilience for clinical bacteriology, we review the concept of resilience used in the fields of socio-ecological systems and microbiology. We then review resilience-related concepts in microbiology, including bacterial tolerance and persistence, phenotypic heterogeneity and collective tolerance and resistance. We argue that the appli-cability of the concept of antibiotic resilience largely relies on representative experimental indicators and we discuss how antibiotic resilience could be measured and used to classify the response of bacterial strains to antibiotic treatments. Such an indicator, in addition to the commonly used minimal inhibitory concentration (MIC) to assess antibiotic resistance, could be a valuable tool to anticipate the success or failure of antibiotic treatments.
2. What is resilience?
The resilience concept is widely developed in the field of socio-ecology and has various meanings. In microbiology, the use of this concept is more recent and mostly restrained to the field of microbial ecology. To have a global view of resilience, we review the main concepts from both fields.
(a) Borrowing concepts from socio-ecological systems
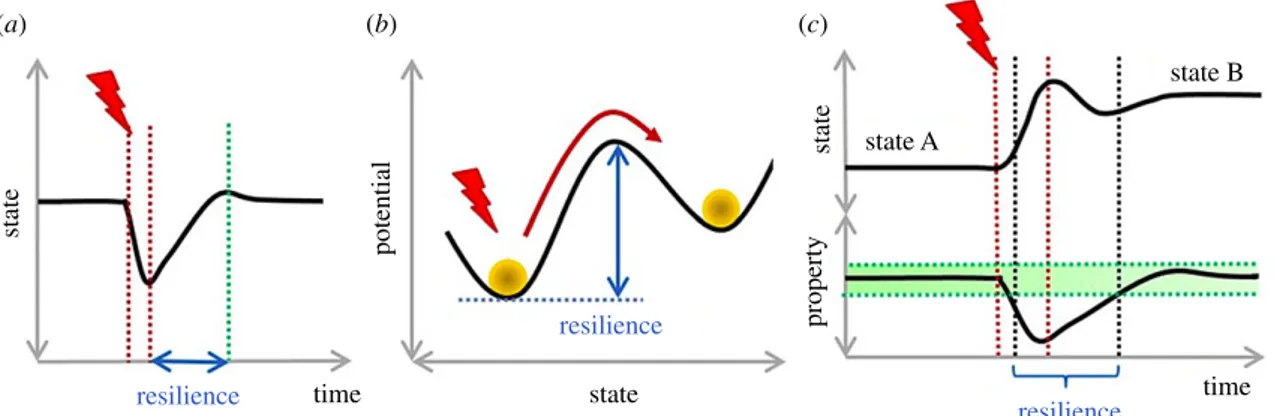
Resilience can take fundamentally different meanings. Holling [16] differentiates two kinds of resilience: engineering resilience and ecological resilience. Engineering resilience is the speed with which a set of given variables recovers after a disturbance. This definition assumes a single equilibrium and does not take into account possible shifts in system behaviours. By contrast, ecological resilience is the amount of disturbance a system can endure before it shifts to an alternative regime with differ-ent properties. It assumes that ecological systems can have
multiple equilibria and shift from one to another under the effect of disturbances [11,17]. In this case, resilience is character-ized by the size of the system’s basin of attraction, i.e. the set of states from which the system tends to a unique equilibrium, also called a stable state or attractor [11,17].
Carpenteret al. [18] add that it is necessary to specify what is resilient to what disturbances:‘resilience of what to what?’ To be operational, it is necessary to state clearly which properties of a system we take into account, and what are the disturbances we consider. Hence, resilience is not the resilience of a whole system but the resilience of a restricted number of properties. This has also been called specific resilience [19]. For Folke [19, p. 260], resilience is more useful as an abstract approach. He differentiates between ecological resilience, focused on the persistence of different equilibria, and socio-ecological resi-lience, focused on adaptation, transformation, learning and innovation. He states that‘the resilience approach is concerned with how to persist through continuous development in the face of change, and how to innovate and transform into new more desirable configurations’. In this view, disturbances are necessary to create opportunities and recombination, themselves necessary for systems to evolve.
The viability-based framework of resilience, which is not specific to socio-ecological systems, is oriented towards a prop-erty of interest instead of attractors [20]. Unlike specific resilience, which is centred on attractors, viability-based resili-ence is defined as the capacity to recover and maintain a property of interest after a disturbance [14], implying that this property has been lost at some point. Thus, systems can adopt various distinct behaviours, stable or not, and still be resilient as regards the chosen property of interest. Considering this defi-nition, the capacity of a system to change its behaviour, i.e. its composition and interactions, is a key factor of resilience.
Different meanings of resilience are represented in figure 1. Two main streams emerge from the socio-ecological literature on resilience. One focuses on the speed of recovery (engineering resilience) or maintenance (ecological resilience) of a reference state, and the other on the resilience of proper-ties of interest that can be lost and recovered and allows changes in the system’s state.
(b) Resilience in microbiology
In microbiology, the resistance/resilience framework is mainly used in the field of microbial ecology. This field
resilience resilience resilience time time state B state A state state property potential state (a) (b) (c)
Figure 1. Different meanings of resilience. Red lightning: disturbance. (a) Engineering resilience: time necessary to recover the initial state after disturbance.
(b) Ecological resilience: amount of disturbance necessary to shift the system to a different state. (c) Viability-based resilience: capacity to recover and maintain
a property of interest (here the green-shaded area) after having lost it. The state of the system may change. Here, the system moves from state A to state B, both
desirable as they maintain the property of interest. (Online version in colour.)
ro
yalsocietypublishing.org/journal/rspb
Pr
oc.
R.
Soc.
B
286
:
20192408
2usually focuses on the resistance and resilience of the compo-sition and the functions of microbial communities faced with particular disturbances [1,21,22]. Resistance is defined as the insensitivity to a disturbance and resilience as the time required for a community to recover its former composition and functions after a disturbance. Both resistance and resili-ence are parts of the concept of robustness, i.e. the capacity of a biological system to maintain particular functions despite disturbances [23]. However, the compositions and functions chosen vary greatly between studies owing to the great diver-sity of microbial communities. For instance, resilience is much used for microbiome studies to refer to the capacity of the gut microbiota to recover its former composition and functions after exposure to antibiotics [24].
Songet al. [25] recently suggested grouping together eco-logical and engineering resilience by defining resilience as the recovery of a function of interest of microbial communities. This point of view is close to the viability-based view of resi-lience which defines resiresi-lience as the capacity to recover and maintain a property of interest [20]. If we elect to study resi-lience relative to a limited function or a property of interest, changes in system state may favour resilience, i.e. the recov-ery of this particular function or property. For example, if the function of interest is a substrate conversion rate, the microbial community can vary its composition to recover this function and be considered resilient.
In a clinical context, resilience sometimes refers to the capacity of a patient to recover from an infection [26], resili-ence is not taken from the perspective of bacteria like antibiotic resistance is. In a recent paper, Meredith and col-leagues applied the ecological concepts of resistance and resilience to study the response of bacterial pathogens to β-lactam treatments [15]. They propose antibiotic resilience as the ability of a bacterial population to recover after an anti-biotic stress, even though the treatment may kill most of the cells. They measure resilience as the time needed for an untreated bacterial population to reach 50% of its maximal biomass compared to an antibiotic treated population, which in their application greatly depends on the population size. In addition, their framework overlaps other concepts in
microbiology including the classical definition of antibiotic resistance (defined for single cells), bacterial persistence and collective resistance. In microbiology, several concepts related to antibiotic resilience already exist and we review the main ones below.
3. Resilience-related concepts in microbiology
Instead of using resilience, a great diversity of concepts have been developed in microbiology such as bacterial tolerance, persistence, recalcitrance and so on (referenced in table 1 and detailed below). Like resilience, these concepts are some-times blurry and depend on the authors and on the context. These concepts tend to reflect the great diversity of mechan-isms used by bacteria to withstand antibiotic treatments and other disturbances.(a) Bacterial tolerance and persistence
Bacterial antibiotic tolerance is the ability of bacteria to survive long exposure to high antibiotic concentrations [43]. Unlike antibiotic resistance, tolerant bacteria cannot grow in the presence of high antibiotic concentrations. However, tolerant bacteria have a small death rate compared to susceptible bacteria, allowing a tolerant bacterial population to survive hours or days longer than a susceptible population.
Bacterial persistence is the presence of a subpopulation of tolerant cells in a susceptible population of bacteria [44,45]. These tolerant cells were first described by Bigger [46] and have been named‘persisters’. The presence of a minority of persisters leads to a characteristic biphasic kill curve under antibiotic exposure. That is, susceptible cells die quickly during the first treatment phase then persisters survive for a long period. Persisters were first described to be non-growing dormant cells in an isogenic population. Because most antibiotics are effective against actively growing cells, persisters survive antibiotic treatments while the majority of the bacterial population dies quickly. Persisters can resume growth spontaneously on standard media. After antibiotic removal, they allow the recovery of the bacterial population,
Table 1. Resilience-related terms used in microbiology.
term
de
finition
bacterial tolerance
the capacity of a bacterial cell or population to survive long exposure to high concentrations of bactericidal antibiotics
while being unable to grow [27,28]
bacterial persistence
the formation of a subpopulation of slow-growing persister cells tolerant to antibiotics. It results in a heterogeneous
response to treatments and a characteristic biphasic killing curve under antibiotic exposure [28,29]
phenotypic
heterogeneity
the heterogeneity of an isogenic bacterial population, possibly leading to its heterogeneous response to antibiotic
exposure [30,31]
phenotypic resistance
(heteroresistance)
non-genetic phenotypic heterogeneity that temporarily enables subpopulations of bacteria to grow in the presence of a
bactericidal antibiotic [32,33]. Also called heteroresistance [34
–36]
adaptive resistance
a transient phenotypic state induced by environmental conditions which increases the survivability of bacteria [37,38]
collective tolerance
the capacity of a suf
ficiently large bacterial population to tolerate the presence of antibiotic although single cells are
susceptible [39]
collective resistance
the capacity of a suf
ficiently large bacterial population to grow in the presence of antibiotic although single cells are
susceptible [40,41]
recalcitrance
the global capacity of a bacterial population to withstand an antibiotic treatment [27,42]
ro
yalsocietypublishing.org/journal/rspb
Pr
oc.
R.
Soc.
B
286
:
20192408
3which is again composed of a majority of susceptible cells and a subpopulation of persisters. Persisters could be formed by stochastic switches in the equilibrium of toxin/antitoxin modules although this hypothesis is still debated [47–49], or induced by stressful conditions such as entry into stationary phase [50]. Stress responses have been reported to be involved in the formation of persisters: stationary phase, starvation or low antibiotic concentrations often increase the level of persisters in bacterial populations [44]. Thus, the persister phenotype is actively maintained by bacteria and not merely a non-growing state [51].
(b) Phenotypic heterogeneity
Although the term‘persister’ was first used to refer to dormant cells, its signification evolved with our knowledge of persist-ence mechanisms. Recent reviews emphasize that persisters do not comprise one single phenotype but, rather, are hetero-geneous [44]. That is, persisters surviving one type of stress may express a different phenotype than persisters surviving another type of stress, although being genetically identical. It is sometimes unclear if bacterial persistence refers to the for-mation of a subpopulation of dormant cells or more generally to the phenotypic heterogeneity of a bacterial populations lead-ing to their partial survival durlead-ing treatments. The phenotypic heterogeneity of an isogenic bacterial population can emerge from‘persisters’ but also from various mechanisms. Hetero-geneity in enzyme production or in efflux pump activity have been reported to induce heterogeneous stress tolerance in bacterial populations [34,37]. InEscherichia coli, cells could escape the action of antibiotics by switching their phenotype from walled state to L-form [52]. Cells could also escape the action ofβ-lactam antibiotics by being protected by the debris of lysed cells [53]. Fluctuations in transcription [54], gene copy number, location of genes, promoter regions and mutation all affect gene expression and can randomly vary from cell to cell. Sometimes, a minority of cells in isogenic populations is able to grow during antibiotic treatments. Because these cells are normally susceptible (unlike resistant cells) and are actively growing (unlike persisters), this phenomenon has been called ‘phenotypic resistance’, ‘adaptive resistance’ or ‘heteroresistance’ [35–37,55]. Phenotypic heterogeneity allows bacterial populations to adopt a‘bet-hedging’ strategy in unpredictable environments, increasing the chances that a part of the population will survive disturbances such as antibiotic treatments [34,37,40]. While this heterogeneity can be actively maintained by bacteria such as the formation of dormant cells, it can also emerge from the inherent randomness at the molecular level [56].
(c) Collective tolerance and resistance
Antibiotic resistance is usually defined for single cells. However, the effectiveness of antibiotics can be reduced in large bacterial populations. This phenomenon is know as the ‘inoculum effect’, ‘collective resistance’ or ‘collective tolerance’ [39,40]. This phenomenon can occur when bacteria deactivate the antibiotic, e.g. by secreting an enzyme such asβ-lactamase, which becomes effective when bacteria are sufficiently numer-ous [15,41]. In a microbial community and under specific conditions, susceptible cells can also be protected by their resistant neighbours [41,57].
A common form of bacterial collective behaviour leading to treatment failure is the formation of biofilms. A biofilm is com-posed of bacteria agglomerated in a matrix of extra-polymeric
substances. This lifestyle allows bacteria to be grouped in a par-ticular place and behave collectively in a more complex way than their planktonic counterpart. Being organized in biofilms allows bacteria to overcome harsh environments that single cells could not [58]. Biofilms enhance the survivability of bac-terial populations by a variety of mechanisms including limited diffusion of solutes and slow growth rates [40,59,60]. In addition, the spatial heterogeneity of mature biofilms leads to various phenotypes depending on the position of a bacterium in the biofilm. Even when the environment is favourable at the surface of the biofilm, some bacteria are stressed by substrate limitation, metabolite accumulation or pH inside the biofilm and express a tolerant phenotype. Conse-quently, the response of a biofilm to antibiotic treatments is fundamentally heterogeneous.
4. Which definition of antibiotic resilience for
clinical bacteriology?
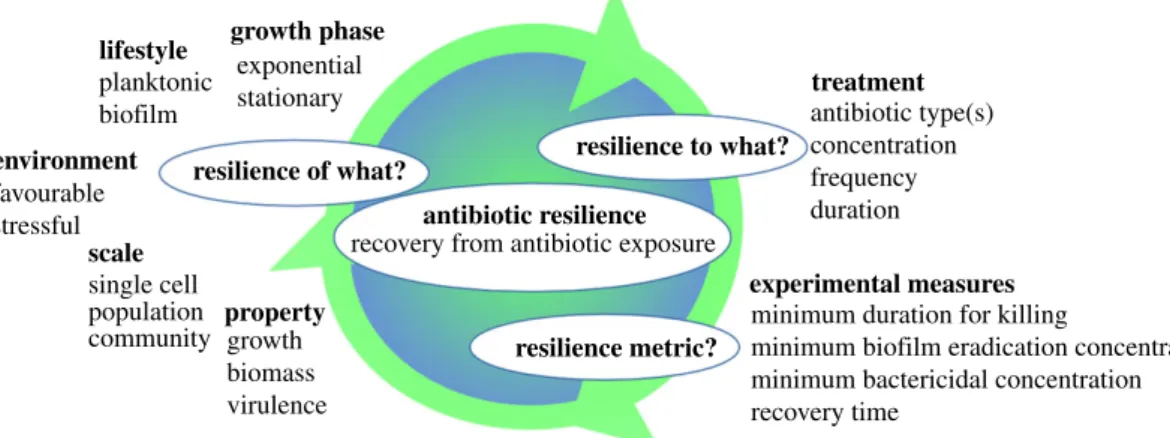
The diversity of survival mechanisms found in bacteria led to a diversity of terms referring to different survival strategies (table 1). This profusion of concepts is difficult to handle and would benefit to be united under a single definition of resilience. On the other hand, the variety of resilience defi-nitions in the literature highlights that resilience depends on what is considered resilient, the disturbances and the context considered (figure 2). Meredithet al. [15, p. 1] propose resili-ence as ‘the ability to recover after being perturbed by an antibiotic’. Although this general definition is easily appre-hended, in practice, they apply the resistance-resilience framework to the particular case of a bacterial population producing β-lactamases and treated with β-lactams, and their framework overlaps with the existing concepts of bacterial persistence and collective tolerance and resistance.
To make operational the resilience concepts, Martinet al. [20] from socio-ecological science and Song et al. [25] from microbial ecology both suggest to define resilience as the capacity to recover a property/function of interest after it has been completely or partially lost owing to a disturbance. In a complementary way, resistance is the capacity to maintain this property/function despite disturbances. Because anti-biotic resistance refers to the capacity of a bacteria to grow in the presence of high concentrations of bactericidal antibiotic, it is tempting to define antibiotic resilience as the capacity of bacteria to resume growth after an antibiotic treatment. However, with this definition, antibiotic resilience would still depend heavily on experimental conditions such as cell den-sity, substrates available and the pre-treatment state of the bacterial population such as its lifestyle (e.g. planktonic or biofilm) and growth phase. While the general definition of anti-biotic resilience seems straightforward—‘the ability to recover after being perturbed by an antibiotic’ [15]—its application readily faces the same problematics as found in socio-ecology, that is a very context-dependent definition.
Antibiotic resistance is mainly based on the measure of the MIC which is then compared to the resistance threshold MIC provided by health institutions. This simple measure makes antibiotic resistance easily understandable, independently of the resistance mechanism at stake. Perhaps antibiotic resilience should also be based on a simple measure or set of measures to be readily applicable by microbiologists, regardless of the various resilience mechanisms and dynamics involved.
ro
yalsocietypublishing.org/journal/rspb
Pr
oc.
R.
Soc.
B
286
:
20192408
45. Which indicator for antibiotic resilience?
Owing to the dynamical nature of resilience, measuring it remains a challenge in ecology [61]. However, having simple, reliable and general indicators of antibiotic resilience is necessary to make this concept applicable. As an indicator of antibiotic resistance, the MIC is the most commonly used in laboratories, that is the capacity of a strain to grow in the presence of a given concentration of antibiotic. However, when the inoculum effect is strong, Artemovaet al. suggest to use the single-cell MIC instead of the classical MIC [62]. Concerning antibiotic resilience, because temporal evolution, phenotypic and spatial heterogeneity are important factors, its measure is not straightforward. The MIC does not give any information on the capacity of a strain to partially survive and recover from antibiotic treatments. To complement the MIC, other measures have been developed. The minimal bac-tericidal concentration is the lowest antibiotic concentration that kills greater than 99.9% of the cells in a planktonic culture overnight [63]. For biofilms, the minimum biofilms eradication concentration is the lowest antibiotic concen-tration necessary to eradicate a biofilm [27], which is often orders of magnitude higher than the MIC [64]. With this method, biofilms are grown on pegs and then dislodged by sonication before the addition of the antibiotic.Brauneret al. [28] proposed an experimental workflow to differentiate between susceptible, resistant, tolerant and per-sistent strains. This method is based on the measure of the MIC and the minimal duration for killing (MDK). The MDK99is the exposure time to an antibiotic required to
elim-inate greater than 99% or more of the bacteria. The MDK is relevant to identify strains which are tolerant or form sub-populations of dormant cells able to survive long period of antibiotic exposure. In their application of the resilience con-cept to antibiotic treatments, Meredith et al. [15] measure resilience as the time needed for a population to reach 50% of its maximal biomass in an unstressed environment com-pared to an environment disturbed by a given antibiotic. However, this measure implies that the recovery occurs while the antibiotic is still present and that the trajectory of the recovery over time is known.
These measures are interesting efforts to quantify the capacity of bacterial populations to overcome antibiotic treat-ments without resistance mechanisms. However, to account
for the dynamical nature of antibiotic resilience and for the bac-terial heterogeneity involved is a difficult task. For instance, biofilms can grow on many different surfaces and have differ-ent stages of maturity. In addition, the proportion of persister cells varies greatly depending on the growth phase of bacterial cultures [65]. Regarding the diversity of biological mechanisms bacterial populations can use to survive antibiotic treatments, the MIC used to determine the resistance or susceptibility of a bacterial strain is clearly limited. The MDK may be the most relevant indicator of antibiotic resilience because the abil-ity to recover from antibiotic treatment depends on the abilabil-ity of bacteria to withstand high concentrations for an extended period. The MDK of bacteria in exponential and stationary phase should provide a first valuable insight into the antibiotic resilience of a bacterial strain, in a similar way that the MIC pro-vides a first valuable insight into its antibiotic resistance, regardless of the specific mechanisms at stake. However, the MDK threshold value that should be considered worrying in a clinical context remains to be evaluated.
6. Concluding remarks
Resilience and resistance are complementary concepts in ecol-ogy. Similarly, the concept of antibiotic resilience could be complementary to antibiotic resistance. While antibiotic resistance focuses on the ability of a strain to grow under high concentrations of antibiotic, antibiotic resilience focuses on the ability of bacteria to recover from antibiotic exposure. This capacity largely relies on the phenotypic heterogeneity of bacterial populations. It joins other ecological concepts such as response diversity [66,67], i.e. the diversity of responses to disturbances, and bet-hedging strategies [31], i.e. random variations increasing the chance to respond adequately to environmental change; strategies which may play a major role in the evolutionary success of bacteria [68,69].
The involvement of biological mechanisms related to anti-biotic resilience in the emergence of antianti-biotic resistance is gaining concern in the scientific community [6,38,70], and the development of strategies to oppose these mechanisms with specific molecules is in progress [43,58,71]. Antibiotic resilience could complement antibiotic resistance in our understanding of bacterial response to treatments. However, the use of the con-cept of antibiotic resilience in clinical bacteriology is critically
growth phase exponential stationary planktonic biofilm lifestyle treatment antibiotic resilience resilience to what? resilience of what? resilience metric?
recovery from antibiotic exposure
antibiotic type(s) concentration frequency duration
experimental measures
minimum duration for killing
minimum biofilm eradication concentration minimum bactericidal concentration recovery time environment favourable stressful scale single cell population
community propertygrowth biomass virulence
Figure 2. The context of antibiotic resilience. While the general definition of antibiotic resilience is easily apprehensible
—the ability to recover from antibiotic
exposure (regardless of the mechanism)
—in practice, antibiotic resilience widely depends on the bacterial system under consideration, on the disturbance applied
and on the metric used to quantify resilience.
ro
yalsocietypublishing.org/journal/rspb
Pr
oc.
R.
Soc.
B
286
:
20192408
5related to its measurement. We urge microbiologists to use new indicators complementary to the MIC such as the MDK and to evaluate their relevance in clinical settings [28,72]. The measure of antibiotic resilience should be a useful indicator to identify and classify strains able to overcome antibiotic treatments despite lacking antibiotic resistance mechanisms.
Data accessibility.This article does not contain any additional data.
Authors’ contributions.C.F. and J.-D.M. obtained the funding. All authors conceived the study. G.C. wrote the manuscript. All authors gave fruitful comments during the research process and revised the manuscript.
Competing interests.We declare we have no competing interests. Funding.This work was supported by a Contrat Plan Etot Région from the Région Auvergne, France.
Acknowledgements.We thank Damien Balestrino and two anonymous reviewers for their helpful advice on the manuscript.
References
1. Shade A et al. 2012 Fundamentals of microbial community resistance and resilience. Front. Microbiol. 3, 417. (doi:10.3389/fmicb.2012.00417) 2. Griffiths BS, Philippot L. 2013 Insights into the
resistance and resilience of the soil microbial community. FEMS Microbiol. Rev. 37, 112–129. (doi:10.1111/j.1574-6976.2012.00343.x) 3. Levy SB, Marshall B. 2004 Antibacterial resistance
worldwide: causes, challenges and responses. Nat. Med. 10, S122–S129. (doi:10.1038/nm1145) 4. Levin-reisman I, Ronin I, Gefen O, Braniss I, Shoresh
N, Balaban NQ. 2017 Antibiotic tolerance facilitates the evolution of resistance. Science 355, 826–830. (doi:10.1126/science.aaj2191)
5. Lewis K, Shan Y. 2017 Why tolerance invites resistance. Science 355, 796. (doi:10.1126/science. aam7926)
6. Bakkeren E et al. 2019 Salmonella persisters promote the spread of antibiotic resistance plasmids in the gut. Nature 573, 276–280. (doi:10.1038/ s41586-019-1521-8)
7. Holling CS. 1973 Resilience and stability of ecological systems. Annu. Rev. Ecol. Syst. 4, 1–23. (doi:10.1146/annurev.es.04.110173.000245) 8. Brand FS, Jax K. 2007 Focusing the meaning(s) of
resilience: resilience as a descriptive concept and a boundary object. Ecol. Soc. 12, 23. (doi:10.5751/ES-02029-120123)
9. Flood S, Schechtman J. 2014 The rise of resilience: evolution of a new concept in coastal planning in Ireland and the US. Ocean Coast. Manage. 102, 19–31. (doi:10.1016/j.ocecoaman.2014.08.015) 10. Grimm V, Wissel C. 1997 Babel, or the ecological
stability discussions: an inventory and analysis of terminology and a guide for avoiding confusion. Oecologia 109, 323–334. (doi:10.1007/ s004420050090)
11. Walker B, Holling CS, Carpenter SR, Kinzig A. 2004 Resilience, adaptability and transformability in social-ecological systems. Ecol. Soc. 9, 5. (doi:10. 5751/ES-00650-090205)
12. Martin S. 2004 The cost of restoration as a way of defining resilience: a viability approach applied to a model of lake eutrophication. Ecol. Soc. 9, 8. (doi:10.5751/ES-00659-090208)
13. Deffuant G, Gilbert N. 2011 Viability and resilience of complex systems. Berlin, Germany: Springer. 14. Rougé C, Mathias J-D, Deffuant G. 2013 Extending
the viability theory framework of resilience to
uncertain dynamics, and application to lake eutrophication. Ecol. Indic. 29, 420–433. (doi:10. 1016/j.ecolind.2012.12.032)
15. Meredith HR, Andreani V, Ma HR, Lopatkin AJ, Lee AJ, Anderson DJ, Batt G, You L. 2018 Applying ecological resistance and resilience to dissect bacterial antibiotic responses. Sci. Adv. 4, eaau1873. (doi:10.1126/sciadv.aau1873)
16. Holling CS. 1996 Engineering resilience versus ecological resilience. In Engineering within ecological constraints (ed. PC Schulze), pp. 31–44. Washington, DC: National Academy Press. 17. Gallopín GC. 2006 Linkages between vulnerability,
resilience, and adaptive capacity. Glob. Environ. Chang. 16, 293–303. (doi:10.1016/j.gloenvcha. 2006.02.004)
18. Carpenter S, Walker B, Anderies JM, Abel N. 2001 From metaphor to measurement: resilience of what to what ? Ecosystems 4, 765–781. (doi:10.1007/ si0021-001-0045-9)
19. Folke C. 2006 Resilience: the emergence of a perspective for social-ecological systems analyses. Glob. Environ. Chang. 16, 253–267. (doi:10.1016/j. gloenvcha.2006.04.002)
20. Martin S, Deffuant G, Calabrese JM. 2011 Defining resilience mathematically: from attractors to viability. In Viability and resilience of complex systems (eds G Deffuant, N Gilbert), pp. 15–36. Berlin, Germany: Springer.
21. Girvan MS, Campbell CD, Killham K, Prosser JI, Glover LA. 2005 Bacterial diversity promotes community stability and functional resilience after perturbation. Environ. Microbiol. 7, 301–313. (doi:10.1111/j.1462-2920.2004.00695.x) 22. Meola M, Lazzaro A, Zeyer J. 2014 Diversity,
resistance and resilience of the bacterial communities at two alpine glacier forefields after a reciprocal soil transplantation. Environ. Microbiol. 16, 1918–1934. (doi:10.1111/1462-2920.12435) 23. Kitano H. 2007 Towards a theory of biological
robustness. Mol. Syst. Biol. 3, 137. (doi:10.1038/ msb4100179)
24. Panda S, El Khader I, Casellas F, López Vivancos J, García Cors M, Santiago A, Cuenca S, Guarner F, Manichanh C. 2014 Short-term effect of antibiotics on human gut microbiota. PLoS ONE 9, e0095476. (doi:10.1371/journal.pone.0095476)
25. Song HS, Renslow RS, Fredrickson JK, Lindemann SR. 2015 Integrating ecological and engineering
concepts of resilience in microbial communities. Front. Microbiol. 6, 1–7. (doi:10.3389/fmicb. 2015.01298)
26. Torres BY, Oliveira JHM, Thomas Tate A, Rath P, Cumnock K, Schneider DS. 2016 Tracking resilience to infections by mapping disease space. PLoS Biol. 14, 1–19. (doi:10.1371/journal.pbio.1002436) 27. Hall CW, Mah T-F. 2017 Molecular mechanisms of
biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol. Rev. 41, 276–301. (doi:10.1093/femsre/fux010) 28. Brauner A, Fridman O, Gefen O, Balaban NQ. 2016
Distinguishing between resistance, tolerance and persistence to antibiotic treatment. Nat. Rev. Microbiol. 14, 320–330. (doi:10.1038/nrmicro.2016.34) 29. Harms A, Maisonneuve E, Gerdes K. 2016
Mechanisms of bacterial persistence during stress and antibiotic exposure. Science 354, aaf4268. (doi:10.1126/science.aaf4268)
30. Bódi Z et al. 2017 Phenotypic heterogeneity promotes adaptive evolution. PLoS Biol. 15, 1–26. (doi:10.1371/journal.pbio.2000644)
31. Grimbergen AJ, Siebring J, Solopova A, Kuipers OP. 2015 Microbial bet-hedging: the power of being different. Curr. Opin. Microbiol. 25, 67–72. (doi:10. 1016/j.mib.2015.04.008)
32. Szomolay B, Cogan NG. 2015 Modelling
mechanical and chemical treatment of biofilms with two phenotypic resistance mechanisms. Environ. Microbiol. 17, 1870–1883. (doi:10.1111/1462-2920. 12710)
33. Javid B, Sorrentino F, Toosky M, Zheng W, Pinkham JT, Jain N, Pan M, Deighan P, Rubin EJ. 2014 Mycobacterial mistranslation is necessary and sufficient for rifampicin phenotypic resistance. Proc. Natl Acad. Sci. USA 111, 1132–1137. (doi:10.1073/ pnas.1317580111)
34. Wang X et al. 2014 Heteroresistance at the single-cell level: adapting to antibiotic stress through a population-based strategy and growth-controlled interphenotypic coordination. MBio 5, e00942-13. (doi:10.1128/mBio.00942-13)
35. Dewachter L, Fauvart M, Michiels J. 2019 Bacterial heterogeneity and antibiotic survival: understanding and combatting persistence and heteroresistance. Mol. Cell 76, 255–267. (doi:10.1016/j.molcel.2019. 09.028)
36. Andersson DI, Nicoloff H, Hjort K. 2019 Mechanisms and clinical relevance of bacterial heteroresistance.
ro
yalsocietypublishing.org/journal/rspb
Pr
oc.
R.
Soc.
B
286
:
20192408
6Nat. Rev. Microbiol. 17, 479–496. (doi:10.1038/ s41579-019-0218-1)
37. Sánchez-Romero MA, Casadesús J. 2014 Contribution of phenotypic heterogeneity to adaptive antibiotic resistance. Proc. Natl Acad. Sci. USA 111, 355–360. (doi:10.1073/pnas. 1316084111)
38. Sandoval-Motta S, Aldana M. 2016 Adaptive resistance to antibiotics in bacteria: a systems biology perspective. Wiley Interdiscip. Rev. Syst. Biol. Med. 8, 253–267. (doi:10.1002/wsbm.1335) 39. Meredith HR, Srimani JK, Lee AJ, Lopatkin AJ, You
L. 2015 Collective antibiotic tolerance: mechanisms, dynamics and intervention. Nat. Chem. Biol. 11, 182. (doi:10.1038/nchembio.1754)
40. Vega NM, Gore J. 2014 Collective antibiotic resistance: mechanisms and implications. Curr. Opin. Microbiol. 21, 28–34. (doi:10.1016/j.mib. 2014.09.003)
41. Sorg RA, Lin L, van Doorn GS, Sorg M, Olson J, Nizet V, Veening JW. 2016 Collective resistance in microbial communities by intracellular antibiotic deactivation. PLoS Biol. 14, 1–19. (doi:10.1371/ journal.pbio.2000631)
42. Michiels JE, Van den Bergh B, Verstraeten N, Michiels J. 2016 Molecular mechanisms and clinical implications of bacterial persistence. Drug Resist. Updat. 29, 76–89. (doi:10.1016/j.drup. 2016.10.002)
43. Meylan S, Andrews IW, Collins JJ. 2018 Targeting antibiotic tolerance, pathogen by pathogen. Cell 172, 1228–1238. (doi:10.1016/j.cell.2018.01.037) 44. Kint CI, Verstraeten N, Fauvart M, Michiels J. 2012
New-found fundamentals of bacterial persistence. Trends Microbiol. 20, 577–585. (doi:10.1016/j.tim. 2012.08.009)
45. Balaban NQ, Gerdes K, Lewis K, McKinney JD. 2013 A problem of persistence: still more questions than answers? Nat. Rev. Microbiol. 11, 587–591. (doi:10. 1038/nrmicro3076)
46. Bigger J. 1944 Treatment of Staphylococcal infections with penicillin by intermittent sterilisation. Lancet 244, 497–500. (doi:10.1016/ S0140-6736(00)74210-3)
47. Fasani R, Savageau M. 2013 Molecular mechanisms of multiple toxin-antitoxin systems are coordinated to govern the persister phenotype. Proc. Natl Acad. Sci. USA 110, E2528–E2537. (doi:10.1073/pnas. 1301023110)
48. Gelens L, Hill L, Vandervelde A, Danckaert J, Loris R. 2013 A general model for toxin-antitoxin module dynamics can explain persister cell formation in
E. coli. PLoS Comput. Biol. 9, e1003190. (doi:10. 1371/journal.pcbi.1003190)
49. Goormaghtigh F et al. 2018 Reassessing the role of type II toxin-antitoxin systems in formation of Escherichia coli type II persister cells. MBio 9, e00640-18. (doi:10.1128/mBio.00640-18) 50. Balaban NQ, Merrin J, Chait R, Kowalik L,
Leibler S. 2004 Bacterial persistence as a phenotypic switch. Science 305, 1622–1625. (doi:10.1126/ science.1099390)
51. Orman MA, Brynildsen MP. 2013 Dormancy is not necessary or sufficient for bacterial persistence. Antimicrob. Agents Chemother. 57, 3230–3239. (doi:10.1128/AAC.00243-13)
52. Mickiewicz KM et al. 2019 Possible role of L-form switching in recurrent urinary tract infection. Nat. Commun. 10, 4379. (doi:10.1038/s41467-019-12359-3)
53. Podlesek Z, Butala M,Šakanović A, Žgur-Bertok D. 2016 Antibiotic induced bacterial lysis provides a reservoir of persisters. Antonie Van Leeuwenhoek 109, 523–528. (doi:10.1007/s10482-016-0657-x) 54. Lv C, Li X, Li F, Li T. 2014 Constructing the energy
landscape for genetic switching system driven by intrinsic noise. PLoS ONE 9, e88167. (doi:10.1371/ journal.pone.0088167)
55. Gollan B, Grabe G, Michaux C, Helaine S. 2019 Bacterial persisters and infection: past, present, and progressing. Annu. Rev. Microbiol. 73, 359–385. (doi:10.1146/annurev-micro-020518-115650) 56. Richard M, Yvert G. 2014 How does evolution tune
biological noise? Front. Genet. 5, 374. (doi:10.3389/ fgene.2014.00374)
57. Medaney F, Dimitriu T, Ellis RJ, Raymond B. 2016 Live to cheat another day: bacterial dormancy facilitates the social exploitation of β-lactamases. ISME J. 10, 778–787. (doi:10.1038/ ismej.2015.154)
58. Römling U, Balsalobre C. 2012 Biofilm infections, their resilience to therapy and innovative treatment strategies. J. Intern. Med. 272, 541–561. (doi:10. 1111/joim.12004)
59. Jolivet-Gougeon A, Bonnaure-Mallet M. 2014 Biofilms as a mechanism of bacterial resistance. Drug Discov. Today Technol. 11, 49–56. (doi:10. 1016/j.ddtec.2014.02.003)
60. Olsen I. 2015 Biofilm-specific antibiotic tolerance and resistance. Eur. J. Clin. Microbiol. Infect. Dis. 34, 877–886. (doi:10.1007/s10096-015-2323-z) 61. Todman LC, Fraser FC, Corstanje R, Deeks LK, Harris
JA, Pawlett M, Ritz K, Whitmore AP. 2016 Defining and quantifying the resilience of responses to
disturbance: a conceptual and modelling approach from soil science. Nat. Publ. Gr. 6, 1–12. (doi:10. 1038/srep28426)
62. Artemova T, Gerardin Y, Dudley C, Vega NM, Gore J. 2015 Isolated cell behavior drives the evolution of antibiotic resistance. Mol. Syst. Biol. 11, 822. (doi:10.15252/msb.20145888)
63. Mah T-F. 2014 Establishing the minimal bactericidal concentration of an antimicrobial agent for planktonic cells (MBC-P) and biofilm cells (MBC-B). J. Vis. Exp. 83, e50854. (doi:10.3791/50854) 64. Marquès C, Tasse J, Pracros A, Collin V, Franceschi C,
Laurent F, Chatellier S, Forestier C. 2015 Effects of antibiotics on biofilm and unattached cells of a clinical Staphylococcus aureus isolate from bone and joint infection. J. Med. Microbiol. 64, 1021–1026. (doi:10.1099/jmm.0.000125)
65. Carvalho G, Guilhen C, Balestrino D, Forestier C, Mathias J-D. 2017 Relating switching rates between normal and persister cells to substrate and antibiotic concentrations: a mathematical modeling approach supported by experiments. Microb. Biotechnol. 10, 1616–1627. (doi:10.1111/1751-7915.12739) 66. Leslie P, McCabe JT. 2013 Response diversity and
resilience in social-ecological systems. Curr. Anthropol. 54, 114–143. (doi:10.1086/669563) 67. Kahiluoto H, Kaseva J, Hakala K, Himanen SJ, Jauhiainen L, Rötter RP, Salo T, Trnka M. 2014 Cultivating resilience by empirically revealing response diversity. Glob. Environ. Chang. 25, 186–193. (doi:10.1016/j.gloenvcha.2014.02.002) 68. Ryall B, Eydallin G, Ferenci T. 2012 Culture history
and population heterogeneity as determinants of bacterial adaptation: the adaptomics of a single environmental transition. Microbiol. Mol. Biol. Rev. 76, 597–625. (doi:10.1128/MMBR.05028-11) 69. Ferenci T, Maharjan R. 2015 Mutational
heterogeneity: a key ingredient of bet-hedging and evolutionary divergence? Bioessays 37, 123–130. (doi:10.1002/bies.201400153)
70. Cohen NR, Lobritz MA, Collins JJ. 2013 Microbial persistence and the road to drug resistance. Cell Host Microbe 13, 632–642. (doi:10.1016/j.chom. 2013.05.009)
71. Defraine V, Fauvart M, Michiels J. 2018 Fighting bacterial persistence: current and emerging anti-persister strategies and therapeutics. Drug Resist. Updat. 38, 12–26. (doi:10.1016/j.drup.2018.03.002) 72. Brauner A, Shoresh N, Fridman O, Balaban NQ. 2017
An experimental framework for quantifying bacterial tolerance. Biophys. J. 112, 2664–2671. (doi:10.1016/j.bpj.2017.05.014)