Choix des graminées fourragères sur les fermes
laitières québécoises dans un contexte de changements
climatiques
Mémoire
Camille Payant
Maîtrise en sciences animales - avec mémoire
Maître ès sciences (M. Sc.)
Choix des graminées fourragères sur les
fermes laitières québécoises dans un contexte
de changements climatiques
Mémoire
Camille Payant
Sous la direction de :
Édith Charbonneau, directrice de recherche
Doris Pellerin, codirecteur de recherche
Résumé
L’objectif des études présentées dans ce mémoire était de déterminer si la fléole des prés, la graminée la plus couramment utilisée en association binaire avec la luzerne au Québec, sera encore un choix pertinent dans le futur. Sa réponse aux changements climatiques pour le rendement et la valeur nutritive a donc été comparée à trois autres graminées cultivées en association binaire avec la luzerne, soit la fétuque élevée, la fétuque des prés et le brome des prés. Afin de se projeter à l’échelle de la ferme, l’impact des changements climatiques sur les cultures annuelles les plus fréquemment utilisées sur les fermes laitières québécoises a aussi été évalué. Ensuite, les conséquences qu’auront ces changements sur la rentabilité et le bilan environnemental des entreprises laitières ont été rapportées. Pour ce faire, des fermes virtuelles de deux régions du Québec au climat contrasté et aux systèmes de production différents, l’Est-du-Québec (Bas-Saint-Laurent) et l’Ouest du Québec (Montérégie), ont été créées. Afin de valider la pertinence de conserver la fléole des prés en association binaire avec la luzerne dans le futur, les rendements et valeurs nutritives des quatre associations fourragères et des cultures annuelles les plus cultivées au Québec ont été simulés grâce au modèle Integrated Farm System Model (IFSM). Puis, les résultats ont été utilisés comme entrées dans le modèle Nutrient Cycling : Crops, Livestock, Environnement and Soil (N-CyCLES) afin de déterminer l’impact de chacune des associations fourragères en contexte de changement climatique sur le bénéfice net et le bilan environnemental des fermes laitières québécoises dans le futur. Dans l’Est-du-Québec, on remarquait que c’est l’association luzerne - fétuque élevée qui devrait le plus bénéficier des changements climatiques, bien que l’association luzerne - fléole des prés devrait aussi en profiter, mais avec une augmentation des rendements projetés moins importante. Dans l’Ouest du Québec, les changements climatiques devraient aussi amener de meilleures performances agronomiques, mais à une moindre échelle étant donné la température déjà plus élevée dans cette région. Le bénéfice net des fermes laitières devrait aussi augmenter. Ce résultat est plus flagrant dans l’Est-du-Québec, car il devrait être possible sous les conditions climatiques projetées d’y cultiver et de vendre du maïs grain et du soya. Puisque plus de cultures seront exportées, la différence entre les entrées et sorties d’azote et de phosphore du système de la ferme devrait diminuer dans le futur. Les émissions totales de gaz à effet de serre devraient rester constantes, mais plus de gaz à effet de serre devraient être attribués aux récoltes vendues. Les résultats des
deux études suggèrent que l’association binaire avec la fétuque élevée serait une bonne alternative à la fléole des prés sous les futures conditions climatiques et que l’Est-du-Québec devrait plus bénéficier des changements climatiques que l’Ouest du Québec.
Abstract
The objective of the studies presented in this document was to determine if timothy, the grass most frequently use in binary association with alfalfa in Quebec, will still be a relevant choice in the future. Its response to climate change in terms of yield and nutritive value were then compared to three other grasses grown in a binary association with alfalfa, which were tall fescue, meadow fescue, and meadow bromegrass. To project at the farm scale, the impacts of climate change on the annual crops mostly used on Quebec dairy farms were also evaluated. Then, the consequences of these changes on the profitability and environmental balance of dairy farms were reported. To do so, virtual farms in two areas of the province of Quebec with contrasted climate and different production system, Eastern Quebec (Bas-Saint-Laurent) and South West Quebec (Montérégie), were created. To validate the relevance to keep timothy as the grass in binary association with alfalfa, the yield and nutritive value of the four forage associations and the ones of the major annual crops in Quebec were simulated with the Integrated Farm System Model (IFSM). Then, the results were used as entries in the Nutrient Cycling: Crops, Livestock, Environment and Soil (N-CyCLES) model to determine the impact of every forage association under the climate change conditions on the net income and environmental balance of dairy farms. In Eastern Quebec, the association alfalfa - tall fescue should benefit more from climate change. Although the association alfalfa - timothy should benefit from climate change, it is the association with the least projected increase in yield. In South West Quebec, climate change should also allow greater agronomic performances, but at a least extend because of the already higher temperature in this area. The net income of dairy farms should also increase. This is more obvious in Eastern Quebec, because it should be possible under future projected climate conditions to grow and sell corn grain and soybean. Because more crops should be exported, the N and P balances should decrease in the future. The total green house gas emissions should be maintained, but more should be allocated to sold crops. Results suggest that the alfalfa-tall fescue mixture could be an alternative to the alfalfa-timothy association under future climate conditions and that Eastern Quebec could benefit more from the climate change then South West Quebec.
Table des matières
Résumé ... ii
Abstract ... iv
Table des matières ... v
Liste des tableaux ... viii
Liste des figures ... xii
Liste des abréviations ... xvi
Remerciements ... xviii
Avant-propos ... xix
Introduction ... 1
Chapitre 1 Revue des travaux antérieurs ... 3
1.1 Changements climatiques attendus ... 4
1.2 Différences physiologiques entre les plantes C3 et C4 ... 6
1.3 Effets des changements climatiques sur le rendement des cultures présentes sur les fermes laitières québécoises ... 9
1.3.1 Dioxyde de carbone ... 9
1.3.2 Température ... 12
1.3.3 Précipitations ... 14
1.3.4 Interactions ... 14
1.4 Effets des changements climatiques sur la valeur nutritive des cultures présentes sur les fermes laitières ... 16
1.4.1 Dioxyde de carbone ... 16
1.4.2 Température ... 19
1.4.3 Précipitations ... 21
1.4.4 Interactions ... 24
1.5 Effets des changements climatiques sur maladies et ravageurs des cultures présentes sur les fermes laitières ... 28
1.5.1 Dioxyde de carbone ... 29
1.5.2 Température ... 32
1.5.3 Précipitations ... 33
1.6.1 Luzerne ... 36
1.6.2 Fléole des prés ... 41
1.6.3 Fétuque élevée ... 45
1.7 Changements climatiques et productions des autres cultures présentes sur les fermes laitières ... 47
1.7.1 Blé ... 47
1.7.2 Orge... 52
1.7.3 Maïs ... 55
1.7.4 Soya ... 59
1.8 Modélisation à l’échelle de la ferme... 62
1.8.1 IFSM ... 62
1.8.2 N-CyCLES ... 64
1.9 Impact économique d’une modification aux choix des espèces fourragères... 66
1.10 Impact environnemental d’une modification dans le choix des espèces fourragères ... 67
1.11 Conclusion de la section ... 69
Chapter 2 Projected impact of future climate conditions on the agronomic performance of four binary grass-alfalfa mixtures in Quebec dairy farms ... 70
2.1 Résumé ... 71
2.2 Abstract ... 72
2.3 Introduction ... 73
2.4 Material and Methods ... 75
2.4.1 Climate data and scenarios ... 75
2.4.2 Projected climatic conditions... 76
2.4.3 Integrated Farm System Model ... 76
2.4.4 Virtual dairy farm ... 77
2.4.5 Calibration ... 79
2.4.6 Assumptions ... 79
2.5 Results and discussion ... 81
2.5.2 Forage and major crop performance ... 81
2.6 Conclusion ... 87
2.7 Acknowledgments ... 88
2.8 Bibliography ... 89
Chapitre 3 Projected impact of future climate conditions on profitability and environmental performances of four binary alfalfa-grass mixtures in Quebec dairy farms ... 104
3.1 Résumé ... 105
3.2 Abstract ... 106
3.3 Introduction ... 107
3.4 Material and Methods ... 110
3.4.1 N-CyCLES ... 110
3.4.2 Virtual farms ... 110
3.4.3 Climatic periods and scenarios ... 112
3.4.4 Simulations ... 112
3.4.5 Rotations ... 113
3.4.6 Sensitivity analysis ... 114
3.5 Results and discussion ... 115
3.5.1 Crop rotations ... 115
3.5.2 Net income ... 115
3.5.3 Nutrient balance and GHG emissions... 117
3.5.4 Sensitivity analysis ... 122 3.6 Conclusion ... 123 3.7 Acknowledgments ... 124 3.8 Bibliography ... 125 Conclusions ... 139 Bibliographie ... 141
Liste des tableaux
Tableau 1.1. Différence (%) de l’effet du CO2 sur la composition chimique des plantes entre
les espèces fixatrices d’azote et celles non fixatrices ainsi qu’avec ou sans fertilisation en azote (adapté de Robinson et al., 2012) ... 18 Tableau 1.2. Concentrations moyennes en protéine brute dans les composantes de biomasse
de graminées en réponse au réchauffement et aux précipitations réduites avec ou sans coupe pendant la saison de croissance. Les données proviennent de trois sites de prairies tempérées du nord dans l’ouest du Canada (tiré de White et al., 2014) ... 21 Tableau 1.3. Synthèse de méta-analyse présentant les hypothèses a priori concernant les
réponses des plantes à une teneur élevée de CO2 atmosphérique et à l’interaction entre
le CO2 et d’autres facteurs (tiré de Robinson et al., 2012)a ... 25
Tableau 1.4. Impacts généraux possibles des éléments de changements climatiques sur la prévalence des espèces aliènes invasives (tiré de Dukes et Mooney, 1999)a ... 29 Tableau 1.5. Réponses des insectes herbivores lorsqu'ils sont nourris de feuilles de plantes
hôtes qui ont grandi dans un environnement enrichi vs ambiant en CO2 (tiré de Lincoln
et al., 1993). ... 30 Tableau 1.6. Réponse de la biomasse de plantes C3 et C4 lorsque la concentration en CO2
atmosphérique est doublée (tiré de Patterson et al., 1999). ... 32 Tableau 1.7. Rendements saisonniers moyens de six associations fourragères binaires
(graminée + luzerne) mesurés au cours des trois premières années de production dans une région froide et une région chaude de la province de Québec ainsi que le bénéfice net des fermes types où l’association fourragère est cultivée (adapté de Pomerleau-Lacasse et al., 2018 et Ouellet et al., 2018). ... 36 Tableau 1.8. Effet du CO2, de la température, et de la disponibilité de l'eau sur la matière
sèche totale 30, 45 et 60 jours après l’émergence d’une luzerne nodulée (P < 0,05) (tiré de Aranjuelo et al. 2005). ... 38 Tableau 1.9. Comparaison de quatre traitements de conditions de croissance (trois régimes
de température jour/nuit et deux régimes de photopériode) sur différents paramètres de la fléole des prés (tiré de Bertrand et al., 2008). ... 44 Tableau 1.10. Réponse des variables physiologiques des plantes à un doublement des
Tableau 1.11. Résumé des moyennes (± erreur type) des températures minimums létales (TLmin), maximum létales (TLmax), de base (Tmin), optimales (Topt) et maximales (Tmax) pour différents processus et phases phénologiques du blé (tiré de Porter et Gawitt, 1999). ... 49 Tableau 1.12. Valeurs moyennes de conductance stomatique à la valeur moyenne globale de
l'index Ball-Berrya, telles que déterminées par l'analyse de covariance pour des feuilles de blé et d'orge qui ont poussé à trois concentrations de CO2 atmosphérique (tiré de
Bunce, 2000). ... 54 Tableau 1.13. Réponse de différents paramètres de l’orge cultivée à des concentrations
atmosphériques de CO2 ambiantes ou élevées en chambre de croissance, ou en parcelles
ouvertes au champ (tiré de Sæbø et Mortensen, 1996)... 54 Tableau 1.14. Biomasse des cannes et grains, nombre de grains, masse individuel des grains,
surface de feuilles totales, date à l'anthèse et date à l'apparition des soies du maïs qui a poussé à des concentrations de CO2 ambiante (370 µmol·mol-1) ou élevée (550
µmol·mol-1) (tiré de Leakey et al., 2006a). ... 57 Tableau 1.15. Taux d'assimilation net moyen (TAM), ratio de surface de feuilles (RSF) et
taux de croissance relatif (TCR) pour la période du jour 26 (récolte 1) au jour 37 (récolte 2) du maïs cultivé dans des sols humides ou secs et sous des teneurs faibles ou élevées de CO2 atmosphérique (tiré de Samarakoon et Gifford, 1996). ... 59
Tableau 1.16. Changement dans l'assimilation de la plante complète (µmol CO2·m-2·s-1) et le
taux d’assimilation net moyen (TAM; g·m-2·jour-1) dans le soya en fonction de
l’augmentation de la température (20, 25, 30 et 35°C) et de la concentration en CO2
atmosphérique (350 et 700 µmol·mol-1) (tiré de Ziska et Bunce (1997). ... 60 Table 2.1. Climate scenarios used for the simulation of each scenario1 according to the
number of forage harvests allowed by the accumulation of growing degree-days in Eastern Quebec and South West Quebec. ... 94 Table 2.2. Characteristics of the two virtual farms, one in Eastern Quebec (EQ) and the other
in South West Quebec (SWQ). ... 95 Table 2.3. Growth parameters used to simulate growth of the four mixtures in IFSM1. ... 96
Table 2.4. Length of the growing season (± SD), corn heat unit (CHU) accumulation (± SD), and growing degree-days (GDD) accumulation (± SD) for the reference period, the near future, and distant future in Eastern Quebec and South West Quebec. ... 97 Table 2.5. Day of harvest and growing degree-days accumulation at each harvest, for each
model, and for the near and distant future in Eastern Quebec and South West Quebec. ... 98 Table 2.6. Yields (t·ha-1 ± SD) and two nutritive value attributes (% DM ± SD) of four alfalfa-grass mixtures harvested as silage and hay along with those of the major annual crops cultivated in dairy farms in Eastern Quebec during the reference period, the near future, and the distant future. ... 99 Table 2.7. Alfalfa proportion (%) at each harvest (± SD) for each forage mixture for the
reference period, the near future, and the distant future in Eastern Quebec... 100 Table 2.8. Yields (t·ha-1 ± SD) and two nutritive value attributes (% DM ± SD) of four alfalfa-grass mixtures harvested as silage and hay along with those of the major annual crops cultivated in dairy farms in South West Quebec during the reference period, the near future, and the distant future. ... 101 Table 2.9. Alfalfa proportion (%) at each harvest (± SD) for each forage mixture for the
reference period, the near future, and the distant future in Eastern Quebec... 102 Table 2.10. Number of days under water stress (± SD) for the major crops used on dairy
farms in future in Eastern Quebec and South West Quebec for the reference period, the near future, and the distant future. ... 103 Table 3.1. Climate scenarios used for the simulation of each scenario1 according to the
number of harvests allowed by the growing degree-day (GDD) accumulation in Eastern Quebec and South West Quebec. ... 128 Table 3.2. Forage yield, crude protein (CP) and neutral detergent fibre (NDF) concentrations,
and variable costs per ha (excluding fertilizers) of four binary alfalfa-grass associations and major crops in Eastern Quebec (EQ) and South West Quebec (SWQ) (Payant et al., 2020, Chapter 2). ... 129 Table 3.3. Selected rotations with their associated variable costs per ha and N, P2O5, and K2O
needs for the four binary alfalfa-grass mixtures in Eastern Quebec and South West Quebec for the reference period. ... 131
Table 3.4. Rotation selection (%) of crops produced (t DM·yr-1), used (t DM·yr-1), and
purchased (t DM·yr-1) for the four binary alfalfa-grass mixtures used in the reference
period, the near future, and the distant future in Eastern Quebec. ... 132 Table 3.5. Rotations selection (%), of crops produced (t DM·yr-1), used (t DM·yr-1), and
purchased (t DM·yr-1) for the four binary alfalfa-grass mixtures used in the reference period, the near future, and the distant future in South Western Quebec. ... 133 Table 3.6. Net income ($·kg-1 de FPCM) and different sources of incomes and outcomes in
farms from Eastern Quebec and South West Quebec where four binary alfalfa-grass mixtures are grown during the reference period, the near future, and the distant future. ... 134 Table 3.7. Whole-farm phosphorus (P) and nitrogen (N) balance (kg·yr-1) and green house
gas (GHG) emission (eqCO2·kg-1 FPCM) in farms of Eastern Quebec where four binary
alfalfa-grass mixtures are grown during the reference period, the near future, and distant future. ... 135 Table 3.8. Whole-farm phosphorus (P) and nitrogen (N) balance (kg·yr-1) and green house
gas (GHG) emission (eqCO2·kg-1 FPCM) in farms of South West Quebec where four
binary alfalfa-grass mixtures are grown during the reference period, the near future, and distant future. ... 136 Table 3.9. Effects of variations in concentrate prices, forage production costs, milk
production, and forage DM yield of binary alfalfa-grass mixtures on net income (CAN$·kg-1 of fat and protein-corrected milk; FPCM) in Eastern Quebec (EQ) dairy farms during the reference period, the near future, and distant future. ... 137 Table 3.10. Effect of variation in concentrate prices, forage production costs, milk production
and alfalfa-grass mixture on net income (CAN$·kg-1 of fat and protein-corrected milk; FPCM) in South West Quebec (SWQ) dairy farms. ... 138
Liste des figures
Figure 1.1. Mécanisme d’assimilation du CO2 chez les espèces végétales C3 et C4 (tirée de
Yamori et al., 2014) ... 7 Figure 1.2. Effet du traitement de CO2 sur diverses espèces. a) assimilation maximale du CO2
pour trois espèces en 1999. b) ratio C : N des feuilles des deux espèces C3. c)
changement relatif dans les concentrations de composés phénoliques des racines de Bo.
ischaemum (tirée de Gill et al., 2002). ... 10
Figure 1.3. Un examen théorique des possibles mécanismes de régulation puits/source présentant le lien entre la concentration en sucrose (S), la concentration en triose phosphate (TP c.à.-d le produit immédiat de la fixation photosynthétique du CO2), la
concentration en sucrose phosphate (SP) et la voie des pentoses phosphates (RPPP) (tirée de Herold, 1980)... 11 Figure 1.4. Les changements des indices de surface de feuilles des cultures de blé, maïs et
coton sous humidité des sols élevée (symboles pleins) ou faible (symboles vides) et sous des concentrations de CO2 élevées (700 µL·L-1) ou faibles (350 µL·L-1). Les valeurs
sont des moyennes ± É.-T. des cinq plantes pour chaque traitement (tirée de Kang et al., 2002). ... 12 Figure 1.5. Accumulation moyenne (± É.-T.) de la biomasse sur un site en Alberta sous des
conditions de température témoin et de température plus élevée pour chacun des trois traitements de précipitation: réduite, ambiante et ajoutée. La valeur P indique la significativité de chaque paire d’interactions pour chaque valeur de précipitation (tirée de White et al.,2014)... 14 Figure 1.6. Teneur en N par unité de surface des feuilles (Nsurface, g N·m-2) de 13 espèces de
prairies cultivées et mesuré sous des conditions de CO2 ambiantes (barres vides; 368
µmol·mol-1) ou élevées (barres hachurées; 560 µmol·mol-1) et sous des concentrations d'N du sol faibles (sans fertilisation ajoutée) ou élevées (ajout de 4 g N·m-2·année-1). (a) 1998 (11 espèces) et (b) 1999 (12 espèces). Les moyennes de moindre carré (± É.-T.) des interactions espèces × CO2 × N de l’ANOVA sont représentées. Les espèces sont
arrangées par groupe fonctionnel. Chaque année a été analysée séparément (tirée de Lee et al., 2001) ... 17
Figure 1.7. Effet de l’augmentation de la teneur en CO2 seulement, des changements
climatiques seuls, et d’une combinaison des changements climatiques et de l’augmentation en CO2 sur (a) la production, (b) le carbone du sol, (c) la minéralisation
du N, et (d) le facteur de décomposition abiotique dans 7 écorégions. Les résultats sont pour les 25 premières années après la transition vers un nouveau climat (tirée de Parton et al., 1995). Parmi ces choix, le climat canadien ressemble plutôt à un climat tempéré humide. ... 20 Figure 1.8. Concentration en azote de fourrages non pâturés à long terme (U) et non défoliés,
faiblement pâturés (L) ou intensivement pâturés (H), ces deux derniers recevant des défoliations annuelles, et ce, sans (témoin) ou avec une supplémentation en eau (addition). Les intervalles de confiance marqués « 3 » permettent d’évaluer la significativité des différences entre les traitements de pâturage pour un même traitement d’arrosage et les intervalles de confiance marqués « 2 » pour les différences entre les traitements d’arrosage pour un même traitement de pâturage (tirée de Milchunas et al., 1995). ... 22 Figure 1.9. Effet de supplémentation en eau (W) et en azote (N) et leur interaction (Moyennes
des moindres carrés de l’effet d’interaction entre W et N ± É.-T.) sur (A) la concentration en protéine brute (CP) des espèces et (B) le rendement en protéine brute 33 jours après la repousse. Les barres d’erreur avec la même lettre ne sont pas significativement différentes (P>0,05) (tirée de Fanselow et al., 2011). ... 23 Figure 1.10 Nombre de lésions cumulatives (a) et sévérité des maladies exprimées en
pourcentage de la surface des feuilles affectées (b) causées par Colletotrichum
gloeosporiodes sur des plants de Stylosanthes scabra susceptibles cultivés sous 350 ou
700 ppm de CO2 atmosphérique dans des chambres à environnement contrôlé avant
d’être exposé à des inocula se produisant naturellement dans trois parcelles en champ pour 48h à cinq occasions séparées. Les données sont des moyennes et des écarts-types de cinq expositions avec trois plants pour chaque parcelle au champ (tirée de Coakley et al., 1999). ... 31 Figure 1.11. Effet du CO2 atmosphérique (Amb CO2, CO2 ambiant ≈ 400 µmol mol-1 CO2 ;
ou élevé, 700 µmol mol-1 CO2), de la température (Tamb, température ambiante ≈ 19°C
0,4 cm3 cm-3; Drought, sécheresse ≈ 0,2 cm3 cm-3) sur l’efficacité de l’utilisation de l’eau
pour la photosynthèse (WUEph), l’efficacité d’utilisation de l’eau pour la production
(WUEp), la production de matière sèche (DM production) et la consommation hydrique
d’une luzerne nodulée (water consumption). Chaque valeur représente la moyenne ± É.-T. (n=12, WUEph; n=8, WUEp). La WUEph a été déterminée lorsque les plants avaient
56 jours de croissance, alors que la WUEp a été déterminée pour la période incluant la
deuxième (45 jours de croissance) et la dernière (60 jours de croissance) récolte. Chaque valeur représente la moyenne ± É.-T. (n=8). Les lettres différentes indiquent des différences significatives (P < 0,05) entre les traitements (tirée de Aranjuelo et al., 2006). ... 39 Figure 1.12. Effet du CO2 (ambiant soit autour de 400, ou élevée soit à 700 µmol·mol-1),
température (ambiante ou ambiante + 4ºC) et de la souche de bactéries nodulaires sur les teneurs en NDF (A) (% MS), en ADF (B) (% MS), et sur la IVDMD (C) (% MS). Les barres représentent la moyenne ± É.-T.; n=4. Dans chaque groupe, les barres avec la même lettre ne sont pas significativement différentes (P ≤ 0,05) selon le test LSD (tirée de Sanz-Sáez et al., 2012). ... 41 Figure 1.13. Évolution des concentrations en certains éléments dans les feuilles vertes (à
l’anthèse) et les grains (à maturité) de blé de printemps suite à un enrichissement de CO2 atmosphérique. Les données sont exprimées en pourcentage de la concentration
observée à une teneur ambiante de CO2 atmosphérique. L’expérience a été réalisée à
deux sites en Europe. Les symboles plus grands réfèrent au premier site alors que les plus petits réfèrent au deuxième site. Les données des deux traitements azotés ont été regroupées étant donné que les interactions impliquant les traitements de CO2 et d'azote
n’étaient pas significatives pour aucun élément dans aucun des tissus échantillonnés. Les lignes de régression de chaque élément ont été calculées pour les ensembles de données des deux sites. La significativité des analyses de régression est indiquée dans le graphique (ns: non significatif, *P<0,05, **P<0,01, ***P<0,001) (tirée de Fangmeier et al., 1999). ... 51 Figure 1.14 . Évolution du poids et de la teneur en amidon de 1000 grains de blé de printemps
suite à un enrichissement de CO2 atmosphérique (les données proviennent de cinq sites
concentrations observées sous concentration ambiante de CO2. Les données sont
rapportées pour les trois traitements azotés (faible, <100 kg N·ha-1 normal, 100-160 kg
N·ha-1; élevé, >160 kg N·ha-1). Aucune des régressions n’était significative à l’exception de la réponse de l’amidon à l’enrichissement en CO2 à la fertilisation élevée
en azote, qui suivait l’équation : y = 85,6 + 0,0393x (P < 0,01) (tirée de Fangmeier et al., 1999). ... 52 Figure 1.15. Rendement en matière sèche totale des plants (a et b), utilisation cumulative
d’eau (c et d) et efficacité d’utilisation de l’eau (WUE; e et f) à la récolte du maïs cultivé dans des sols humides ou secs et sous des teneurs faibles (barres hachurées) ou élevées (barres pleines) de CO2 atmosphérique. **=P<0,01; *** = P<0,001; NS = non
Liste des abréviations
Amb, ambiant
Asat, point auquel la photosynthèse est saturée par le flux de photon C:N, ratio carbone:azote
CO2, dioxyde de carbone
DACA, différence alimentaire cations anions
GES, gaz à effet de serre GNS, glucides non structuraux gs, conductance stomatique H2O, eau
HDP, haut degré de polymérisation MS, Matière sèche
PB, protéine brute
RSF, ratio de surface de feuilles T, température
TAM, taux d’assimilation net TCR, taux de croissance relatif TH, tétanie d’herbage
TLmax, température létale maximale TLmin, température létale minimale Tmax, température maximale
Tmin, température minimale Topt, température optimale UTM, unité thermique maïs
ADF, acid detergent fiber ADL, acid detergent lignin B, Barley
C, Canola
C:N, ratio Carbon: Nitrogen Cg, Corn grain
CHO, carbohydrate CHU, corn heat unit CO2, Carbon dioxyde CP, Crude protein Cs, Corn silage
DBC, date beginning cumulation DEC, date end cumulation EQ, Eastern Quebec
GDD, Growing degree-days GHG, Green house gas Gs, grass silage
H, hay
i, implantation
IFSM, Integrated Farm System Model IVDMD, in Vitro dry matter digestibility IVTD, in vitro true digestibility
AMB, alfalfa + meadow bromegrass AMF, alfalfa + meadow fescue DM, dry matter
N-CYCLES, Nutrient Cycling: Crops, Livestock, Environment and soils
NDF, neutral detergent fiber NDFd, digestibility of NDF
RCP, representative concentration patways
S, straw So, soybean
SWQ, South West Quebec ATF, alfalfa + tall fescue AT, alfalfa + timothy W, wheat
WUEp, water use efficiency for production
WUEph, water use efficiency for photosyntesis
Vcmax, maximum rate of Rubisco carboxylase activity
Remerciements
La réalisation de ce projet avec tout ce qu’il implique a été très enrichissante, que ce soit personnellement ou professionnellement. Les deux dernières années n’auraient certainement pas été aussi faciles et plaisantes sans l’équipe qui m’a entourée et c’est pourquoi je désire prendre quelques instants pour remercier tous les gens qui m’ont appuyée.
Premièrement, merci à ma directrice de maîtrise, Édith Charbonneau, qui m’a offert un soutien extraordinaire à toutes les étapes. Merci de m’avoir acceptée dans ton équipe et de m’avoir permis de vivre toutes ces belles expériences. Surtout, merci de toujours avoir laissé ta porte ouverte pour répondre à mes millions de questions et pour m’aider à faire face à mes petites insécurités.
Merci à Doris Pellerin, mon codirecteur, d’avoir pris la place d’Édith pour me prêter main-forte lorsqu’elle était absente et de m’avoir partagé un peu de sa sagesse.
Un merci tout particulier à Guillaume Jégo de chez Agriculture et Agroalimentaire Canada qui m’a toujours accueillie à bras ouverts afin de m’aider avec le casse-tête que peut être la modélisation. Tes connaissances très approfondies sur le sujet et ton grand calme m’ont souvent été d’un grand secours.
Merci beaucoup aux organismes de financement, Ouranos et le gouvernement du Québec avec son initiative du Fonds vert/ PACC 2013-2020, sans qui ces recherches n’auraient pas été possibles.
Finalement, le plus grand des mercis à toutes les personnes qui m’ont soutenue dans mon quotidien; mon conjoint, ma famille et mes merveilleuses amies. Merci d’avoir partagé mes joies et de m’avoir aidée à surmonter les moments difficiles. Je vous aime.
Avant-propos
Ce mémoire comprend, au Chapitre 1, une revue de littérature portant sur les changements climatiques attendus, les différences physiologiques entre les plantes C3 et C4 et les impacts
des changements climatiques sur le rendement et la valeur nutritive des cultures ainsi que sur les ennemis des cultures. Puis, plus spécifiquement, l’impact des changements climatiques sur les cultures pérennes et annelles les plus couramment cultivées au Québec est abordé. Finalement, les dernières sections du chapitre portent sur la modélisation à l’échelle de la ferme et sur l’impact économique et environnemental découlant d’une modification de choix d’espèce de graminée fourragère.
Au chapitre 2, un premier article scientifique est présenté en anglais. Il traite de l’impact projeté des futures conditions climatiques sur les performances agronomiques de quatre associations binaires graminée-luzerne sur les fermes laitières du Québec. Cet article sera soumis au Agronomy Journal pour publication.
Le chapitre 3 présente le deuxième article scientifique portant sur l’impact projeté des futures conditions climatiques sur la rentabilité et les performances environnementales de quatre associations binaires graminée-luzerne sur les fermes laitières québécoises. Ce chapitre est aussi présenté en anglais. Cet article sera soumis pour publication dans Agricultural System
Journal.
Je suis l’auteure principale du présent mémoire et j’ai réalisé les projections de rendement, valeur nutritive, rentabilité et bilan environnemental, ainsi que l’analyse des résultats. Au chapitre 2, les coauteurs sont G. Jégo, V. Ouellet, P. Grenier, G. Tremblay, G. Bélanger et É. Charbonneau. Au chapitre 3, les coauteurs sont G. Jégo, V. Ouellet, P. Grenier, G. Tremblay, G. Bélanger, D. Pellerin, A. Vanasse, et É. Charbonneau.
Introduction
Les changements climatiques sont maintenant considérés comme une réalité et de nombreux impacts sont attendus au Québec, ce qui affectera l’agriculture telle qu’on la connaît aujourd’hui. Selon Ouranos (2015), la température dans le sud du Québec devrait augmenter de 2 à 4 °C entre 2041 et 2070, et de 4 à 7 °C entre 2070 et 2100. Pour le Nord québécois, on pourrait même s’attendre à des augmentations de 5 à 10 °C d’ici la fin du siècle. Ces changements se traduiraient par un raccourcissement de la saison d’enneigement et une augmentation de la durée des vagues de chaleur. Par contre, cela permettrait l’allongement de la saison de croissance. De plus, il pourrait y avoir une augmentation des précipitations totales, mais une diminution des précipitations sous forme de neige dans le sud du Québec et certains phénomènes météorologiques extrêmes pourraient être plus fréquents et/ou plus marqués. En effet, on pourrait s’attendre à une augmentation de la fréquence et de l’intensité des orages. Par contre, Ouranos (2015) indique que le nombre de jours consécutifs sans précipitations devrait être plus élevé pendant la période estivale avec les changements climatiques. Les experts rapportent aussi un risque d’apparition de nouveaux ravageurs ainsi que de nouvelles mauvaises herbes et maladies des cultures.
La fléole des prés, l’espèce de graminée la plus couramment cultivée dans les prairies québécoises, ne sera pas épargnée par ces changements. En effet, l’augmentation des températures devrait se traduire par une augmentation de l’évapotranspiration chez les plantes qui subiront alors plus d’épisodes de sécheresse. Or, bien que la fléole des prés soit bien adaptée aux hivers québécois, cette dernière répond très mal à la sécheresse (Bertrand et al., 2008). Ainsi, connaissant les bénéfices d’un mélange entre graminées et légumineuses (Sturludóttir et al., 2014), il serait judicieux d’identifier une graminée alternative à la fléole des prés, lorsque cultivée en partenariat avec la luzerne. Dans le futur, Thivierge et al. (2016) mentionnent que cette interaction bénéfique devrait persister puisque dans un mélange graminée et légumineuse, la légumineuse donne de l’azote à la graminée ce qui permettrait à cette dernière de répondre à l’augmentation des besoins en azote associé à l’augmentation de l’assimilation de carbone sous une concentration en CO2 élevée. Sous le climat actuel, six
démontré que la fétuque élevée, la fétuque de près et le brome des prés représentaient des alternatives intéressantes à la fléole des prés (Pomerleau-Lacasse et al., 2018).
Ainsi, ces résultats et les connaissances actuelles sur les changements climatiques ont mené à se questionner sur l’impact des changements climatiques sur les associations fourragères actuellement recommandées, qui sera traité dans cette étude. Dans un premier temps, l’état des connaissances actuelles sur les effets des changements climatiques sur le rendement et la valeur nutritive des cultures les plus fréquemment utilisées au Québec ainsi que l’intérêt de la modélisation des fermes laitières sont présentés dans une revue de littérature. Ensuite, deux études ont été réalisées dans le cadre de ce mémoire. La première visait à utiliser le modèle
Integrated Farm System Model (IFSM) afin de prédire les effets des changements climatiques
(radiations solaires, températures, précipitations, humidité et concentration atmosphérique de CO2) sur les rendements et la valeur nutritive de quatre graminées (fléole des prés, fétuque
élevée, fétuque des prés et brome des prés) cultivées en association avec la luzerne. Les rendements et valeurs nutritives des grandes cultures annuelles les plus répandues au Québec ont aussi été évalués. Dans la seconde étude, les modifications de rendements et valeurs nutritives des cultures amenées par les changements climatiques ont été utilisées comme entrées dans le modèle Nutrient Cycling : Crops, Livestock, environment and soil (N-CyCLES) afin d’évaluer l’effet sur le bénéfice net et le bilan environnemental (bilan N et P et émissions de GES) des entreprises laitières québécoises.
Chapitre 1
1.1 Changements climatiques attendus
Ouranos (2015) s’est penché sur les changements climatiques attendus dans la province de Québec et l’impact de ces changements sur, entre autres, l’agriculture. Dans la province, on s’attend à avoir une augmentation de la température de 2 à 4°C entre 2041 et 2070, et de 4 à 7°C pour la période 2071-2100. De plus, la saison d’enneigement devrait être plus courte alors que les vagues de chaleur devraient être plus longues au cours de l’été. Les auteurs prévoient aussi une possible augmentation de la quantité annuelle des précipitations, bien que ces dernières seront distribuées différemment. De plus, les phénomènes météorologiques extrêmes, tels les orages, pourraient être plus fréquents et/ou plus importants. La hausse des températures devrait favoriser l’évapotranspiration chez les plantes, qui pourraient alors expérimenter plus fréquemment des périodes de stress hydrique (Thivierge et al., 2017). Il est aussi possible que la température optimale de croissance de certaines cultures soit dépassée (Cassman et al., 2010). Cependant, les changements attendus sous les futures conditions climatiques permettraient aussi d’allonger la saison de croissance (Ouranos, 2015). De plus, la concentration plus importante en CO2 atmosphérique devrait permettre
l’augmentation de l’effet de fertilisation du CO2 (McGrath et Lobell, 2013). L’effet de
fertilisation de CO2 fait référence à l’augmentation du taux de photosynthèse chez les plantes
suite à l’augmentation de la concentration en CO2 atmosphérique. En effet, Ouranos (2015)
a formulé plusieurs hypothèses quant à la concentration atmosphérique en CO2 dans le futur
selon différents scénarios. Selon le scénario le plus optimiste, où les émissions de GES sont réduites, la concentration atmosphérique en équivalent CO2 atteindrait un pic à 490 ppm en
2050 avant de redescendre. Le scénario le plus pessimiste prévoit plutôt une concentration atmosphérique en équivalent CO2 de 1370 ppm en 2100. Finalement, les scénarios
intermédiaires prévoient des concentrations entre 650 et 850 ppm équivalent CO2 en 2100.
Comme les plantes utilisent le CO2 comme source de carbone, les rendements pourraient
ainsi être augmentés. On pourrait aussi s’attendre à ce qu’il y ait plus de mauvaises herbes, d’insectes et/ou de maladies et que de nouveaux ennemis des cultures entrent dans la province (Gagnon et al., 2013). Finalement, les changements climatiques attendus pourraient permettre de cultiver de nouvelles cultures ou encore de donner la possibilité de cultiver des
cultures dans des régions plus fraîches qu’il est actuellement seulement possible de cultiver dans les régions plus chaudes (Bryant et al., 2000).
1.2 Différences physiologiques entre les plantes C
3et C
4Il est important de se remémorer quelques notions de physiologie végétale avant d’aller plus loin dans la relation entre le climat et la productivité des plantes. On s’attend à des augmentations de CO2 atmosphérique dans le futur (Ouranos, 2015) et toutes les plantes ne
réagiront pas de manière aussi efficace à cette augmentation. Selon Long et al. (2004), les plantes répondent à une hausse de la concentration en CO2 en augmentant la photosynthèse
et en diminuant la conductance stomatique. Selon ces auteurs, tous les autres effets d’une teneur élevée en CO2 sur les plantes et les écosystèmes dérivent de ces deux réponses
fondamentales.
Afin de comprendre comment les changements climatiques et l’augmentation de la concentration atmosphérique de CO2 affectent la photosynthèse et par le fait même la
croissance des plantes, il est important de comprendre le mécanisme d’assimilation du CO2
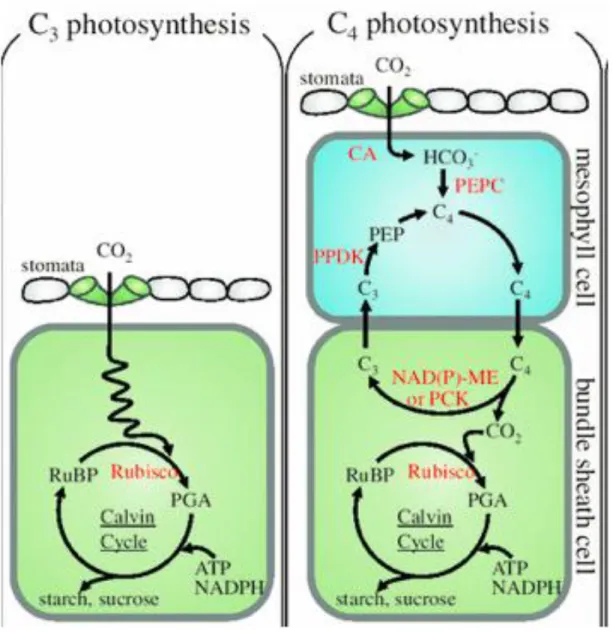
des différentes espèces végétales (figure 1.1). En effet, il existe trois types de mécanismes de captation du CO2 : les C3, tels la fléole des prés, la luzerne, le blé, l’orge et le soya, les C4, tel
Figure 1.1. Mécanisme d’assimilation du CO2 chez les espèces végétales C3 et C4 (tirée de
Le mécanisme de concentration du CO2 dans les plantes C4 augmente la concentration en
CO2 au site actif de 12 à 20 fois en comparaison aux plantes C3 (Kiirats et al., 2002; von
Caemmerer et Furbank, 2003). Ainsi, avec les concentrations actuelles de CO2
atmosphérique, ce site est déjà saturé. De plus, ce type de plante évite la photorespiration.
Pour les C3, le taux maximal de photosynthèse possible est plutôt limité par le taux maximal
de carboxylation possible par la rubisco (Anderson et al., 2001). Dans le futur, si la quantité de CO2 atmosphérique disponible est plus élevée suite à l’augmentation de sa concentration,
le taux de carboxylation pourra augmenter en conséquence, tout en évitant le gaspillage d’énergie (Ainsworth et Long, 2005). Cependant, Lee et al. (2001) mentionnent que ce mécanisme ne sera possible que si l’azote apporté à la plante est suffisant. De plus, l’augmentation de la teneur en CO2 atmosphérique déclencherait la fermeture des stomates,
ce qui diminuerait la transpiration, qui à son tour augmenterait l’assimilation nette de carbone, ce qui finalement, se traduirait par une augmentation de la croissance (Ruget et al., 2012). De plus, la fermeture des stomates permettrait de diminuer les pertes en eau. Avec l’augmentation du CO2 atmosphérique, l’avantage compétitif associé au métabolisme des C4
pourrait donc être réduit dans le futur (Sage, 2004), même si celles-ci pourraient tout de même aussi bénéficier d’une augmentation de l’efficacité d’utilisation de l’eau (Patterson, 1995).
En conclusion, les groupes fonctionnels de plantes dont la photosynthèse est limitée par la rubisco à une teneur élevée de CO2, c’est-à-dire les C3, ont plus de potentiel d’augmenter
leur point auquel la photosynthèse est saturée par le flux de photon (Asat) si la fertilisation
azotée est adéquate, que ceux dont la photosynthèse est limitée par la régénération du RubP à un niveau élevé de CO2, c’est-à-dire les C4.
1.3 Effets des changements climatiques sur le rendement des
cultures présentes sur les fermes laitières québécoises
Il est possible que les changements climatiques attendus aient un impact sur le rendement des cultures présentes sur les fermes laitières québécoises. Parmi les facteurs pouvant avoir un effet sur le rendement, la concentration de CO2 atmosphérique, la température et les
précipitations sont analysées dans cette revue. Comme ces facteurs risquent très fortement d’entrer en interaction, les connaissances actuelles sur leur effet conjugué sur le rendement seront décrites afin de déterminer les effets synergiques ou antagonistes.
1.3.1 Dioxyde de carbone
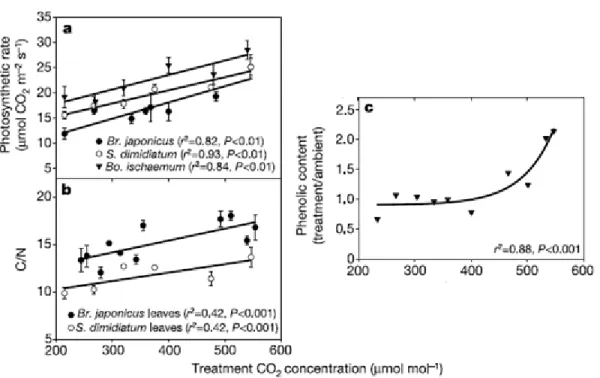
Gill et al. (2002) ont remarqué qu’une élévation de la concentration en CO2 sur un gradient
continu de 200 à 500 µmol·mol-1 permet des taux de photosynthèse supérieurs, pour les raisons mentionnées dans la section précédente. Cette augmentation de la photosynthèse se traduit par une augmentation de la production primaire nette (g·m-2·an-1) d’une prairie d’environ 50 %, mais aussi par une modification dans la chimie des tissus. En effet, on s’attend à une augmentation linéaire du ratio C :N et une augmentation exponentielle de la concentration en composés phénoliques jusqu’à l’atteinte d’un plateau (figure 1.2). La figure démontre une relation positive significative entre l’assimilation maximale du CO2 et la
concentration en CO2 et ce pour toutes les espèces (P<0,01) en plus d’une augmentation
positive et linéaire du ratio C :N pour les deux espèces avec une augmentation du CO2. La
figure montre aussi une augmentation forte et exponentielle dans le contenu en composés phénoliques des racines (P<0,001).
Figure 1.2. Effet du traitement de CO2 sur diverses espèces. a) assimilation maximale du CO2
pour trois espèces en 1999. b) ratio C : N des feuilles des deux espèces C3. c) changement
relatif dans les concentrations de composés phénoliques des racines de Bo. ischaemum (tirée de Gill et al., 2002).
Certains auteurs ajoutent, par contre, que la stimulation de la photosynthèse pourrait ne pas continuer à long terme à cause d’une saturation possible en CO2 des plantes C3 combiné à
une rétro-inhibition de la photosynthèse induite par l’accumulation de glucides (Herold, 1980; figure 1.3). En effet, si le taux d’utilisation du sucrose par les organes puits est inférieur au taux de synthèse par les organes sources, le sucrose s’accumule dans ces derniers. Il en résulte une régulation négative des enzymes impliquées dans la synthèse du sucrose. Certaines plantes seraient, par contre, meilleures pour faire face à un excès de glucides que d’autres (Ainsworth et Rogers, 2007).
Figure 1.3. Un examen théorique des possibles mécanismes de régulation puits/source présentant le lien entre la concentration en sucrose (S), la concentration en triose phosphate (TP c.à.-d le produit immédiat de la fixation photosynthétique du CO2), la concentration en
sucrose phosphate (SP) et la voie des pentoses phosphates (RPPP)(tirée de Herold, 1980). L’effet positif de l’élévation de la teneur en CO2 sur le rendement serait aussi lié à
l’augmentation de la capacité d’extraire l’eau et les nutriments par les racines et ainsi contrebalancer l'humidité faible des sols (Samarkoon et Gifford, 1996; Wall, 2001). Dans la même lignée, chez les plantes C4, la baisse de la disponibilité en eau dans le sol dans le futur
ferait fermer les stomates, ce qui pourrait diminuer la concentration interne de CO2 à des
niveaux auxquels la photosynthèse serait sensible au CO2 atmosphérique (Polley, 2002). Par
contre, dans le cas des plantes C3, l’augmentation de l’efficacité de l’utilisation de l’eau au
niveau des feuilles pourrait ne pas diminuer la consommation d’eau de façon importante, car il y a une augmentation de la surface foliaire totale sous une élévation du CO2 (Kang et al.,
Figure 1.4. Les changements des indices de surface de feuilles des cultures de blé, maïs et coton sous humidité des sols élevée (symboles pleins) ou faible (symboles vides) et sous des concentrations de CO2 élevées (700 µL·L-1) ou faibles (350 µL·L-1). Les valeurs sont des
moyennes ± É.-T. des cinq plantes pour chaque traitement (tirée de Kang et al., 2002).
1.3.2 Température
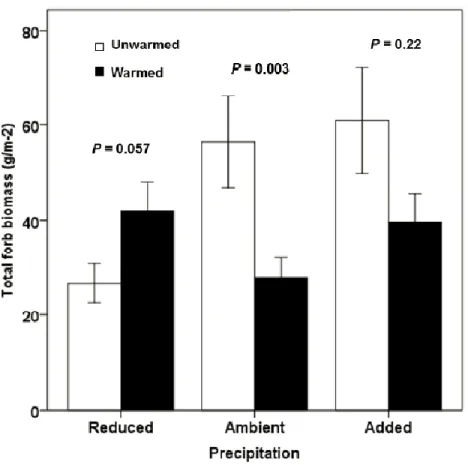
Les processus physiologiques liés à la croissance, par exemple la photosynthèse et la respiration, ont une température optimale jusqu’à laquelle ils augmentent avant de redescendre. Au contraire, le taux de développement et la progression dans le cycle de vie des cultures ont souvent une réponse strictement linéaire à la température (Porter et Semenov, 2005). Ainsi, un changement de température peut avoir comme effet de diminuer les rendements de certaines grandes cultures. White et al. (2014) ont étudié l’effet d’une
augmentation de température de 2 à 4°C sur trois prairies canadiennes à différentes proportions de graminées et arbustes, la plupart des espèces étant indigènes. La température ambiante variait entre 13,5 et 16,5 °C selon le site pendant l’expérience et l’augmentation de température était obtenue dans des chambres à ciel ouvert. Dans cette étude, l’augmentation de la température s’est traduite par une diminution de l’accumulation de biomasse totale (figure 1.5), expliquée majoritairement par une baisse de la proportion de graminées. Ces effets négatifs se manifesteraient puisqu’une augmentation de la température diminue l’humidité du sol, ce qui limite la croissance de la plante (Kardol et al., 2010). En effet, il a été estimé que l’évaporation potentielle d’une prairie augmentera de 2 à 3 % pour chaque augmentation de 1°C de température (Lockwood, 1999). Par contre, il est important de mentionner qu’il est judicieux de prendre en compte la combinaison de l’amplitude et de la durée de l’épisode de stress thermique ainsi que le stade de développement de la plante lorsque celle-ci subit le stress, et non simplement la température absolue (Wheeler et al., 2000).
La baisse de rendement suite à l’augmentation de la température pourrait aussi s’expliquer par le fait que l’affinité de la rubisco pour le CO2 et la solubilité du CO2 relativement à
Figure 1.5. Accumulation moyenne (± É.-T.) de la biomasse sur un site en Alberta sous des conditions de température témoin et de température plus élevée pour chacun des trois traitements de précipitation: réduite, ambiante et ajoutée. La valeur P indique la significativité de chaque paire d’interactions pour chaque valeur de précipitation (tirée de White et al.,2014)
1.3.3 Précipitations
Stout et al. (1988) ont noté qu’au cours des années où les précipitations étaient majoritairement sous forme de gros orages ou faibles ou mal distribués, comme ce qui est attendu dans le futur, c’était la capacité de rétention en eau du sol qui devenait le facteur principal contrôlant le rendement et l’efficacité d’utilisation de l’eau.
1.3.4 Interactions
Il semble clair que les facteurs présentés précédemment entreront en interaction dans le futur, ce qui modifiera les réponses attendues.
Par exemple, l’augmentation de la température combinée à une diminution des précipitations pourrait diminuer le rendement en biomasse des graminées et cela permet aux autres plantes herbacées de prendre l’avantage compétitif (White et al., 2014). Par contre, les effets négatifs de l’augmentation de température pourraient être contrebalancés par les effets du CO2 sur la
tolérance au stress hydrique (Fuhrer, 2003). En effet, sous une concentration élevée en CO2,
la conductance stomatique serait réduite. Combinée à une meilleure densité racinaire attendue, cela devrait se traduira en une meilleure efficacité d’utilisation de l’eau. De plus, les sécheresses attendues pourraient faire augmenter le temps d’activité photosynthétique. Ces deux paramètres mis ensemble se traduiraient par une augmentation des rendements (Clifford et al., 2000).
1.4 Effets des changements climatiques sur la valeur nutritive des
cultures présentes sur les fermes laitières
En plus de l’effet des changements climatiques sur le rendement, il se peut que ces derniers aient des effets directs sur la valeur nutritive des plantes, via les mécanismes de défense ou d’adaptation, ou encore par des effets indirects suite aux changements de rendements. De cette façon, les mêmes facteurs étudiés pour l’impact des changements climatiques sur le rendement seront analysés pour la valeur nutritive.
1.4.1 Dioxyde de carbone
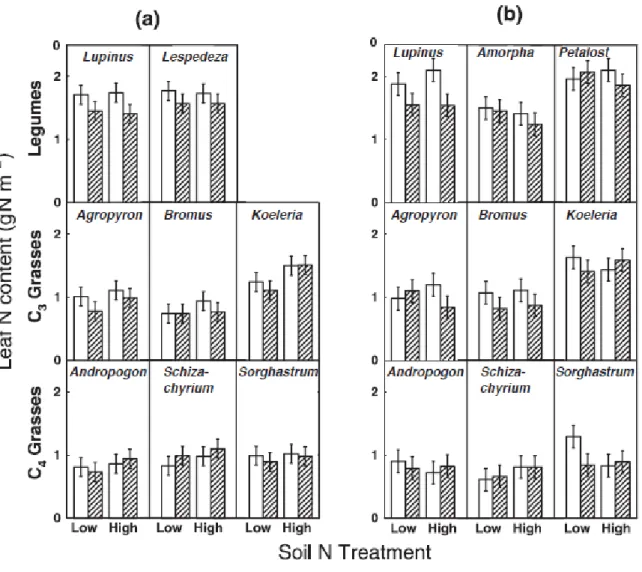
La diminution de la teneur en azote des feuilles (figure 1.6), induite par l’élévation de la teneur en CO2 atmosphérique, est associée à la diminution du potentiel photosynthétique,
probablement via des changements dans les enzymes photosynthétiques riches en azote, comme la rubisco (Lee et al., 2001), ou encore à cause de l’effet de dilution causé par la croissance rapide des plantes. Ainsi, la teneur en CO2 de l’air est positivement corrélée avec
le ratio C :N des tissus. Roumet et al. (2000) notent une diminution de la concentration en azote des feuilles de 19 % par unité de matière sèche chez les graminées et aucun changement significatif chez les légumineuses lors d’une augmentation de 350 à 700 ppm de CO2
atmosphérique. On aperçoit aussi une augmentation exponentielle de la concentration en composés phénoliques avec une augmentation de la teneur en CO2 (Gill et al., 2002; figure
Figure 1.6. Teneur en N par unité de surface des feuilles (Nsurface, g N·m-2) de 13 espèces de
prairies cultivées et mesuré sous des conditions de CO2 ambiantes (barres vides; 368
µmol·mol-1) ou élevées (barres hachurées; 560 µmol·mol-1) et sous des concentrations d'N du sol faibles (sans fertilisation ajoutée) ou élevées (ajout de 4 g N·m-2·année-1). (a) 1998 (11 espèces) et (b) 1999 (12 espèces). Les moyennes de moindre carré (± É.-T.) des interactions espèces × CO2 × N de l’ANOVA sont représentées. Les espèces sont arrangées par groupe
fonctionnel. Chaque année a été analysée séparément (tirée de Lee et al., 2001)
De plus, suite à l’augmentation de la teneur en CO2 de l’air, on note une augmentation de
24 % en moyenne des glucides non structuraux totaux (GNS) totaux chez les 13 espèces de graminées lorsque la concentration en CO2 atmosphérique passait de 368 à 560 µmol·mol-1
analysées dans l’étude de Lee et al. (2001), ce qui est en lien avec ce qui a été mentionné dans les sections précédentes. Par contre, tout comme pour la concentration en azote des
feuilles, les changements dans les teneurs en GNS des légumineuses sont non significatifs. En effet, chez ces dernières, le processus de fixation du N2 représente un puit additionnel ce
qui permet d’éviter le déséquilibre (Roumet et al., 2000). Dans leur méta-analyse, Robinson et al. (2012) notent, quant à eux, une augmentation des tannins de 30 %, du ratio C :N de 27 %, des glucides totaux de 23 %, de l’amidon de 50 %, des sucres solubles de 8 % et des GNS de 39 %, mais une diminution de la concentration en azote de 16 % et des glucides structuraux de 13 % suite à une élévation de la concentration de CO2 atmosphérique. Les
auteurs ont aussi fait la différence entre l’effet du CO2 sur la composition chimique des
plantes des espèces fixatrices d’azote et celles non fixatrices ainsi qu’avec ou sans fertilisation en azote. Les résultats sont présentés au tableau 1.1.
Tableau 1.1. Différence (%) de l’effet du CO2 sur la composition chimique des plantes entre
les espèces fixatrices d’azote et celles non fixatrices ainsi qu’avec ou sans fertilisation en azote (adapté de Robinson et al., 2012)
Effet de l’augmentation de la teneur en CO2 ambiante (368 µmol·mol-1) à élevée (560
µmol·mol-1) chez plusieurs espèces regroupées dans 170 études (herbacées, graminées, arbustes, arbres)
Espèces fixatrices de N. Espèces non fixatrices de N.
N total - 10 -17
Glucides totaux + 30 +20
Protéines -16 -8
Composés phénoliques totaux +35 +16
Composés flavonoïdes totaux +60 +13
Effet de passer des concentrations d'N du sol faibles (sans fertilisation ajoutée) à élevées (ajout de 4 g N·m-2·année-1). Avec fertilisation (systèmes agriculturals) Sans fertilisation (systèmes naturels) N total -13 -17 Protéines solubles -11 -19 Acides aminées -10 -25 Ratio C :N +16 +20
Métabolites secondaires à base
1.4.2 Température
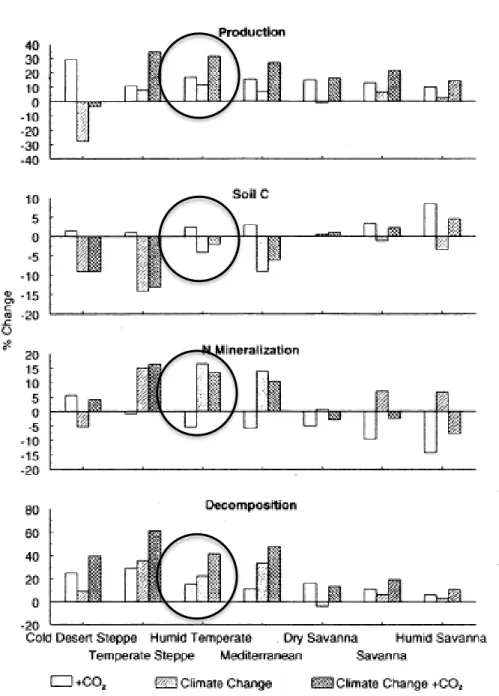
L’augmentation de la température pourrait avoir un effet positif sur la disponibilité des nutriments puisqu’elle crée une augmentation du taux de minéralisation, ce qui augmenterait la disponibilité de l’azote du sol et qui entraînerait une augmentation de la productivité (Parton et al., 1995; figure 1.7). Par contre, Parton et al. (1995) mentionnent qu’il y a une augmentation de la perte d’azote si la plante n’est pas en synchronisme avec la disponibilité en azote.
Figure 1.7. Effet de l’augmentation de la teneur en CO2 seulement, des changements
climatiques seuls, et d’une combinaison des changements climatiques et de l’augmentation en CO2 sur (a) la production, (b) le carbone du sol, (c) la minéralisation du N, et (d) le facteur
de décomposition abiotique dans 7 écorégions. Les résultats sont pour les 25 premières années après la transition vers un nouveau climat (tirée de Parton et al., 1995). Parmi ces choix, le climat canadien ressemble plutôt à un climat tempéré humide.
D’un côté plus négatif, les conditions qui accélèrent la maturité des plantes, telle une augmentation de la température, auront des effets négatifs sur la teneur en protéines brutes
de la plante (tableau 1.2), probablement expliquée par une maturation accélérée et à une diminution du ratio feuilles : stomates (Buxton, 1996).
Tableau 1.2. Concentrations moyennes en protéine brute dans les composantes de biomasse de graminées en réponse au réchauffement et aux précipitations réduites avec ou sans coupe pendant la saison de croissance. Les données proviennent de trois sites de prairies tempérées du nord dans l’ouest du Canada (tiré de White et al., 2014)
Protéine brute (g·kg-1 MS)
Traitement Niveau Sans coupe Avec
coupe Précipitation ambiante 87 a 110a réduite 81b 98b Réchauffement non 86 a 106a oui 82a 102b
Coupe faible intensité 99
a
haute intensité 109b
a,bDans une même colonne et un même traitement, les moyennes suivies de la même lettre ne
diffèrent pas significativement à P > 0,05.
1.4.3 Précipitations
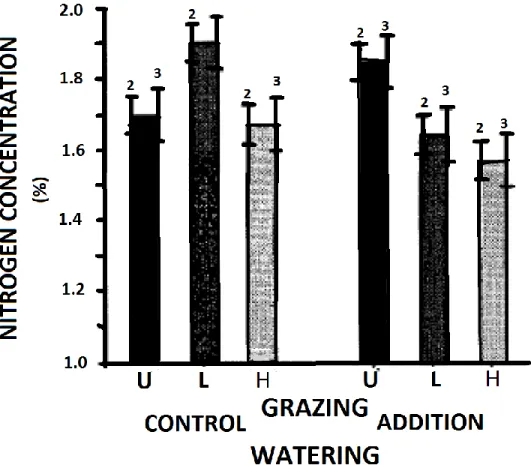
Tout comme sous une augmentation de la température, la teneur en protéines brutes de la plante peut diminuer sous une diminution sévère des précipitations, potentiellement à cause d’une translocation des nutriments de la partie aérienne vers les racines (Buxton, 1996) ou encore à cause de la sénescence forcée amenée par le manque d’eau. Au contraire, une augmentation des précipitations augmenterait la biomasse, ce qui diluerait l’azote (figure 1.8) et diminuerait aussi, par conséquent, la concentration en protéine brute (Milchunas et al., 1995). Toutefois, dû à l’augmentation simultanée de la biomasse, le rendement en protéines net augmente (figure 1.9). Cependant, dans des écosystèmes semi-arides, la diminution de la concentration en azote peut être compensée par une augmentation simultanée de l’assimilation de l’azote (Fanselow et al., 2011). Toutefois, il est encore inconnu à savoir si ce mécanisme est expliqué par une amélioration du système racinaire ou une amélioration du transport de l’azote dans le sol par une plus grande quantité d’eau dans les pores du sol.
Figure 1.8. Concentration en azote de fourrages non pâturés à long terme (U) et non défoliés, faiblement pâturés (L) ou intensivement pâturés (H), ces deux derniers recevant des défoliations annuelles, et ce, sans (témoin) ou avec une supplémentation en eau (addition). Les intervalles de confiance marqués « 3 » permettent d’évaluer la significativité des différences entre les traitements de pâturage pour un même traitement d’arrosage et les intervalles de confiance marqués « 2 » pour les différences entre les traitements d’arrosage pour un même traitement de pâturage (tirée de Milchunas et al., 1995).
Figure 1.9. Effet de supplémentation en eau (W) et en azote (N) et leur interaction (Moyennes des moindres carrés de l’effet d’interaction entre W et N ± É.-T.) sur (A) la concentration en protéine brute (CP) des espèces et (B) le rendement en protéine brute 33 jours après la repousse. Les barres d’erreur avec la même lettre ne sont pas significativement différentes (P>0,05) (tirée de Fanselow et al., 2011).
1.4.4 Interactions
Tout comme pour le rendement, on peut s’attendre à ce que les paramètres des changements climatiques entrent en interaction pour venir modifier la valeur nutritive des cultures. Il est connu, par exemple, qu’une augmentation de la température combinée à une diminution des précipitations pourrait avoir des effets négatifs sur la teneur en protéine brute de la plante (White et al., 2014) et que chez les graminées de grande qualité, l’augmentation de la teneur en CO2 couplé à l’élévation de la température semblerait n’avoir que des impacts mineurs
sur la digestibilité et la composition de la fibre (Porter et Semenov, 2005). Les nombreuses interactions entre différents paramètres et leurs effets sont présentés au tableau 1.3.
Tableau 1.3. Synthèse de méta-analyse présentant les hypothèses a priori concernant les réponses des plantes à une teneur élevée de CO2 atmosphérique et à l’interaction entre
le CO2 et d’autres facteurs (tiré de Robinson et al., 2012)a
Réponse de la plante CO2 CO2 × N. CO2 × H2O CO2 × T CO2 × lumière
Concentration en carbone
Référencesb ↓ attendue à haut niveau de
CO21 Pas d'hypothèse Pas d'hypothèse Pas d'hypothèse Pas d'hypothèse
Auteursc Non supportées Pas d'évidence d'interaction Pas d'évidence d'interaction Pas d'évidence d'interaction Données insuffisantes Concentration en azote
Références ↓ attendues à haut niveau de CO22
↓ supérieures à faible N. Pas d'hypothèses Pas d'hypothèses Pas d'hypothèses Auteurs Confirmées *** Confirmées * Pas d'évidence d'interaction Pas d'évidence d'interaction Pas d'évidence d'interaction
Concentration en amidon
Références ↑ attendue à haut niveau de
CO23 ↑ supérieures à faible N Pas d'hypothèses ↑ inférieures à haute T Pas d'hypothèses Auteurs Confirmées *** Pas d'évidence d'interaction Pas d'évidence d'interaction Pas d'évidence d'interaction Données insuffisantes
Concentration en CHO solubles
Références ↑ attendue à haut niveau de
CO23 Pas d'hypothèses Pas d'hypothèses ↑ supérieures à haute T
Auteurs Confirmées ** Pas d'évidence d'interaction Pas d'évidence d'interaction Pas d'évidence d'interaction Pas d'évidence d'interaction Concentration en GNS
Références ↑ attendue à haut niveau de
CO23 ↑ supérieures à faible N. Pas d'hypothèses ↑ inférieures à haute T Pas d'hypothèses Auteurs Confirmées *** Pas d'évidence d'interaction Données insuffisantes Données insuffisantes Données insuffisantes
Concentration en acides aminés
Références ↓ attendue à haut niveau de
CO2 ↑ supérieure à faible N. Pas d'hypothèses Pas d'hypothèses Pas d'hypothèses Auteurs Confirmées *** Pas d'évidence d'interaction Données insuffisantes Pas d'évidence d'interaction Données insuffisantes
Concentration en protéines
Références ↓ attendue à haut niveau de
CO2 ↓ supérieure à faible N. Pas d'hypothèses Pas d'hypothèses Pas d'hypothèses Auteurs Confirmées *** Pas d'évidence d'interaction Pas d'évidence d'interaction Pas d'évidence d'interaction Données insuffisantes
Réponse de la plante CO2 CO2 × N. CO2 × H2O CO2 × T CO2 × lumière Solidité des feuilles
Références ↑ attendue à haut niveau de
CO2 Pas d'hypothèses Pas d'hypothèses Pas d'hypothèses Pas d'hypothèses
Auteurs Confirmées *** Pas d'évidence d'interaction Données insuffisantes Pas d'évidence d'interaction Données insuffisantes Biomasse
Références ↑ attendue à haut niveau de
CO24-7 ↑ supérieures à haut N. ↑ inférieure à faible H2O ↑ supérieure à haute T
↑ supérieures à haute lumière ou sans différence
Auteurs Confirmées *** Confirmées *** Pas d'évidence d'interaction
Ratio C :N
Références ↑ attendue à haut niveau de
CO2 ↑ supérieure à faible N. Pas d'hypothèses Pas d'évidence d'interaction Pas d'évidence d'interaction Auteurs Confirmées *** Pas d'évidence d'interaction Données insuffisantes Pas d'hypothèses Pas d'hypothèses
Eau
Références ↑ attendue e EUE à haut
niveau de CO2 Pas d'hypothèses
↑ inférieures de EUE à faible
H2O Pas d'évidence d'interaction Pas d'évidence d'interaction
Auteurs ↓ contenu en eau; EUE non
évaluée Pas d'évidence d'interaction Pas d'évidence d'interaction Pas d'hypothèses Pas d'hypothèses Ratio racine :pousse
Références ↑ attendue à haut niveau de
CO2 ↑ supérieure à haut N. ↑ supérieure à faible H2O Pas d'évidence d'interaction Pas d'évidence d'interaction Auteurs Non supportées Pas d'évidence d'interaction Pas d'évidence d'interaction Pas d'hypothèses Pas d'hypothèses Métabolites secondaires basés sur
N
Références ↓ attendue à haut niveau de
CO2 ↓ supérieure à faible N. Pas d'hypothèses Données insuffisantes Données insuffisantes Auteurs Confirmées *** Pas d'évidence d'interaction Pas d'évidence d'interaction Pas d'hypothèses Pas d'hypothèses Métabolites secondaires basés sur
C
Références ↑ attendue à haut niveau de
CO23 Pas d'hypothèse Pas d'hypothèse
Glucosides totaux ↑ sous T
Réponse de la plante CO2 CO2 × N. CO2 × H2O CO2 × T CO2 × lumière
Auteurs Phénoliques ↑; Terpènes ↓
Pas d'évidence d'interaction pour composés phénoliques ou terpènes totaux, mais sous
CO2 élevé, les taux d'émission de terpènes ↑ sous
N élevé et ↓ sous faible N
Pas d'évidence d'interaction pour composés phénoliques ou terpènes totaux, mais sous
CO2 élevé, les taux d'émission de terpènes ↓ plus
si contenu en eau élevé
↑ inférieure à haute T Pas d'hypothèses
a Les textes en bleu dénotent où l’évidence conforme apparemment l’hypothèse (* P<0,05; **P<0,01; ***P<0,001); les textes en rouge dénotent où l’évidence contredit apparemment l’hypothèse; et les textes
en vert dénotent de nouvelles hypothèses émergentes de cette analyse.
b Résultats attendus à priori basés sur les travaux expérimentaux de la littérature c Ce que les auteurs de la méta-analyse ont trouvé
1Augmentation [C] plus grande pour plantes C
3 que C4. Les auteurs ont trouvé : confirmées* pour glucides totaux et amidon, mais pas d’évidence d’interaction pour GNS ou CHO solubles 2Diminution [N] plus grande pour plantes C
3 que C4. Les auteurs ont trouvé : confirmées*
3Amidon, CHO soluble et GNS augmentent plus pour les plantes qui croissent lentement que les plantes qui croissent rapidement. Les auteurs ont trouvé : aucune évidence pour une interaction pour CHO soluble,
CHO totaux ou GNS. Ils ont trouvé une interaction pour l’amidon, mais dans la direction opposée.
4Augmentation biomasse plus importance pour les plantes qui croissent lentement que les plantes qui croissent rapidement. Les auteurs ont trouvé : pas d’évidence d’interaction 5Augmentation biomasse plus importante pour les plantes fixatrices d’azote. Les auteurs ont trouvé : pas d’évidence d’interaction
6Augmentation biomasse plus importante pour les plantes herbacées que pour les planes ligneuses. Les auteurs ont trouvé : interactions significatives, mais dans la direction opposée
7Augmentation biomasse plus importante pour les angiospermes que les gymnospermes. Les auteurs ont trouvé : interactions significatives, mais dans la direction opposée T, température; EUE, efficacité