Nuclear Mechanotransduction in Skeletal Muscle
Texte intégral
Figure
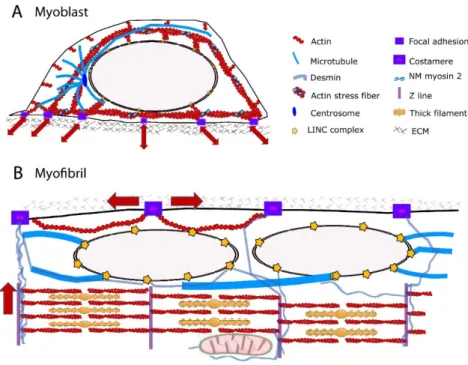
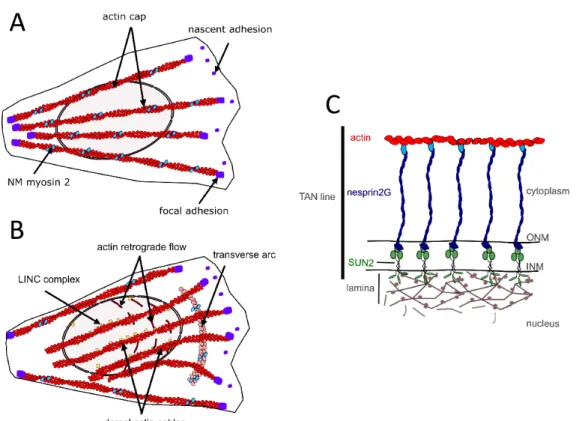
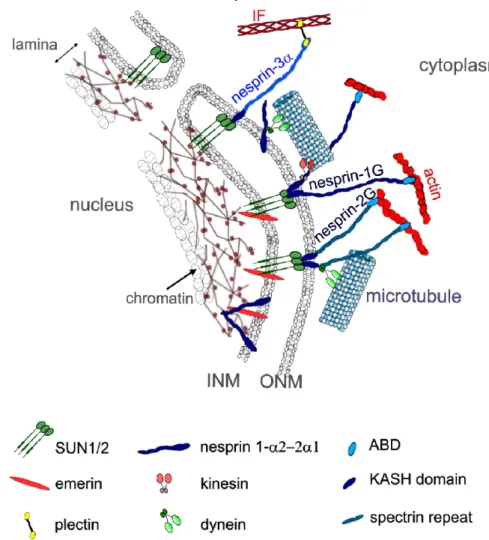
Documents relatifs
mortaLité d'un niveau trophique, elle doit être également assez forte pour. permeÈtre une accumulation
L’absence de répartition des domaines de compétences de principe entre l’Etat et les collectivités territoriales et l’inexistence consécutive d’une procédure spéciale
• Chapter 4: in this chapter, we will present the different possible applications of the proposed models and exhibit preliminar study using the first deformable model that is able
La décade suivante entre pour Jeff Barda dans une phase « performative », l Anne P gal, Oli ie Cadi e Pierre Alféri rencontrent Hocquard à la fin des années
The potential impact of a biogenic alkalinity source on the sulfur budget in the MBL could be better tested with sulfate D 17 O measurements in a region of high oceanic productivity
Argyrophilic grain disease (AgD), first described by Braak and Braak [2, 3] is a sporadic late onset dementia that ac- counts for approximately 5% of all cases of dementia [5, 13,
tive chromatin fraction from round spermatids after 1.5 rain of digestion, lane C same as lane B but after 3-min digestion, lane D MNase-sensitive
- Le premier concerne l’adaptation des acteurs de l’économie maritime, portuaire et logistique aux aléas de la mondialisation : stratégies des armements de
