Analyse de l’influence des ouvrages hydroélectriques et des facteurs biotiques et abiotiques sur les déplacements de la forme résidente de la truite commune aux stades précoces
Texte intégral
Figure
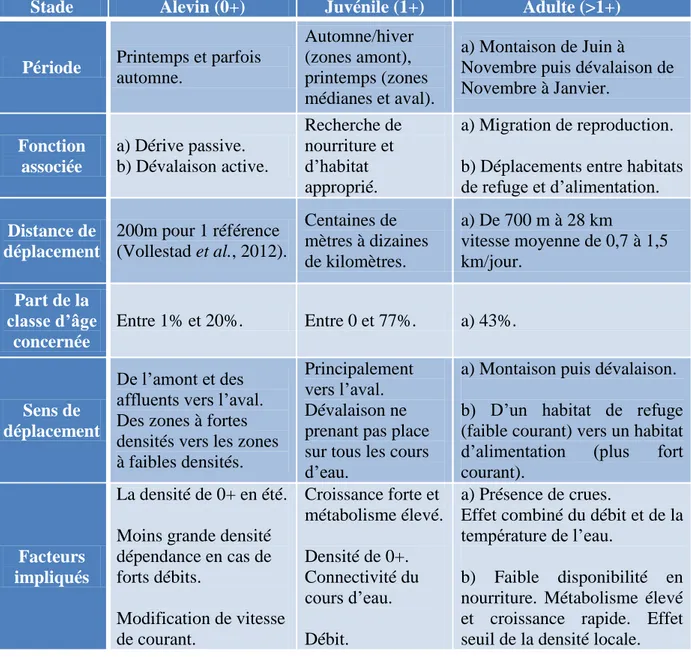
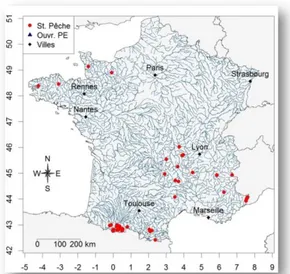
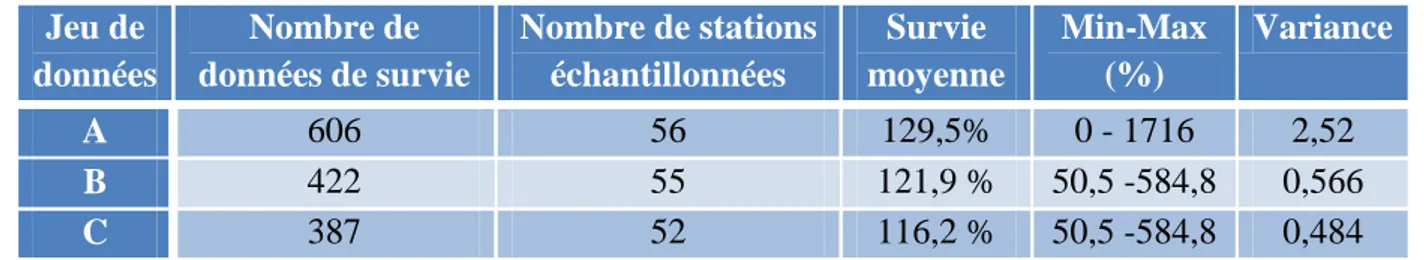
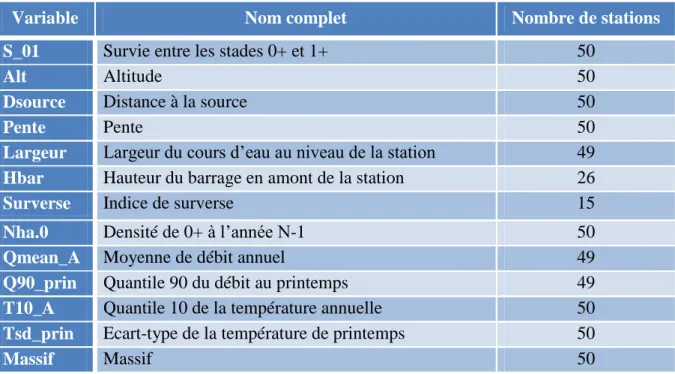
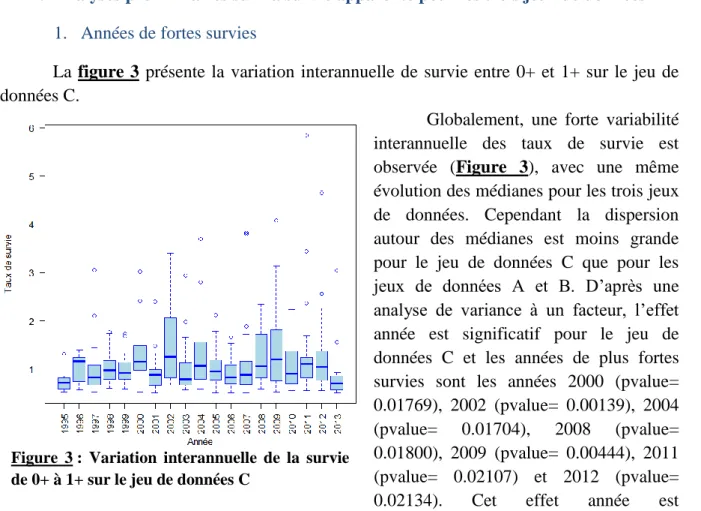
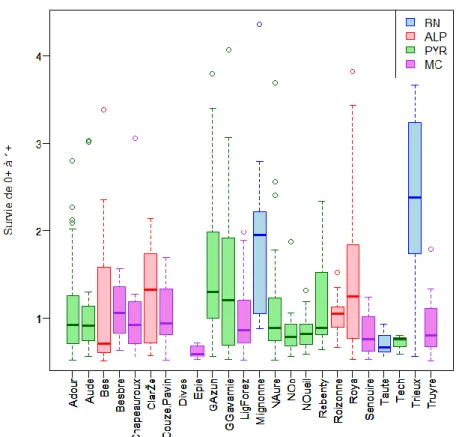
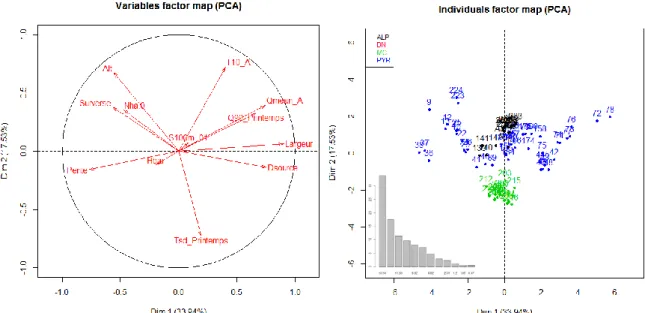

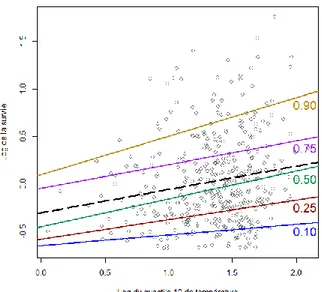
Documents relatifs
Donner la mesure de l’angle marqué en justifiant
[r]
En comparaison avec le lancer de marteau, quel type d’action doit exercer le Soleil sur une planète pour l’empêcher de s’échapper dans l’espace?. (figures 1 et
Les conditions aux limites sont : un flux imposé de 10 4 W/m 2 sur la face inférieure et une température imposée de 20° C sur la face supérieure de la structure.. Les conditions
REPRODUCTION DE FIGURE PAR SYMÉTRIE N° 1.. REPRODUCTION DE FIGURE PAR SYMÉTRIE N°
Une origine (point d’application) Un point où l’on souhaite mesurer la vitesse Sa direction : La droite tangente à la trajectoire au point M. Son sens : Celui
3 -a – Compléter le schéma suivant avec les noms des changements d’état physique convenable ( fusion – vaporisation – solidification – liquéfaction ). 3 –b- Donner le
1 -a – Compléter le schéma suivant avec les noms des changements d’état physique convenable ( fusion – vaporisation – solidification – liquéfaction ). 1 –b- donner le
