Traitement de l’Image et du Signal pour la Biologie
Corentin Spriet, Unité de Glycobiologie Structurale et Fonctionelle, CNRS UMR 8576
Soutenue le 4 juillet 2018 devant le Jury composé de:
Gabriel Paës Directeur de recherche INRA Rapporteur Emanuele Biondi Directeur de recherche CNRS Rapporteur Coralie Bompard Chargée de recherche CNRS Rapporteur Isabelle Fournier Professeur université de Lille Examinateur Emmanuel Margeat Directeur de recherche CNRS Examinateur Jean-François Bodart Professeur université de Lille Garant
Comme vous le verrez tout au long de ce manuscrit, mon parcours professionnel est rythmé par de très nom-breuses rencontres et collaborations interdisciplinaires. Ce sont ces interactions qui renouvellent en permanence le plaisir que je peux avoir à effectuer ce métier, mais qui compliquent la lourde tâche d’écrire des remerciements sur l’ensemble de mon parcours.
Tout d’abord, je tiens à remercier Gabriel Paës, Emanuele Biondi, Isabelle Fournier, Emmanuel Margeat et Cora-lie Bompard de m’avoir fait l’honneur d’accepter de faire partie du jury chargé d’évaluer ce travail malgré vos emplois du temps surchargés! Un grand merci également pour la richesse des discussions lors de la soutenance, j’ai vraiment passé un excellent moment grâce à vous!
Ce manuscrit doit beaucoup à trois personnes:
Jean-François «le Professeur» Bodart, qui me fait l’honneur d’être garant de ce travail et qui m’a clairement poussé et motivé à me lancer dans cette aventure. Merci également pour nos nombreux échanges scientifiques, pour ton soutien professionnel et personnel tout au long de ces années, pour nos stimulantes sessions d’écritures à 4 mains et plus généralement de me suivre dans mes différents délires tout en m’entraînant dans les tiens (c’est pas tous les jours qu’on monte un train brio dans le bureau d’un vice-doyen d’université!).
Dave Trinel, «partner in crime» depuis mes tous débuts. Il s’en est passé des choses depuis notre premier échange téléphonique ou j’étais encore biochimiste. Merci Bro pour ton soutien et l’ensemble de nos échanges tout au long de ces années! Je me réjouis de me lancer dans l’aventure des IAs avec quelqu’un possédant tes qualités scienti-fiques et humaines.
Angelina kasprowicz, étudiante motivée, volontaire, autonome...Typiquement l’étudiante qui donne envie de pas-ser son HDR. Merci pour ces dernières années au laboratoire, c’est un plaisir de travailler avec toi.
Le début de ma carrière scientifique doit énormément à l’ensemble du GDR 2588 et du RTmfm. Merci à vous de m’avoir entraîné du côté obscur de la biophotonique. Mes pensées vont également au regretté professeur Jörg Lan-goswky qui m’a fait l’honneur de me faire découvrir la biophysique et de m’accueillir dans son laboratoire. Merci a tous ses membres, et plus particulièrement à Nicolas Dross d’avoir été mon guide dans les mondes merveilleux de la FCS et des bières allemandes.
Un des virages majeurs de ma carrière a été la création de TISBio avec Dave. Merci à Vincent Villeret puis à Domi-nique Legrand, avec l’aide d’Hafida Kameli, de m’avoir donné cette opportunité. Merci également à Christophe D’Hulst d’avoir «mouillé la chemise», au propre comme au figuré, pour que nous puissions rejoindre le bâtiment
Aymeric, Rédha, Guillaume, Coralie, Lolo, Martine). Merci également aux services techniques et administratifs et à l’ensemble des collègues qui nous ont simplifié la vie au maximum.
Vient alors l’arrivée dans ce nouveau bâtiment particulièrement accueillant, notamment grâce à Fabrice Watte-bled et son équipe, Camille, Coralie, David, Adeline, Xavier, Loub, Sylvain, Christophe qui m’ont accueilli à leur table et m’ont initié au fonctionnement du C9 et de l’université de façon plus générale. À enchaîner les coups de main, les moments de convivialité et de bienveillance, les collaborations ont commencé à se mettre en place. Les quelques images d’amidon isolées du début se transforment en un projet en IA qui devrait nous occuper quelques années, et ce sera un plaisir de les passer en vos compagnies.
L’arrivée au C9 a ainsi été l’occasion de renforcer des collaborations préexistantes et de développer de premiers logiciels pour François Foulquier et son équipe (Sven en super bêta-testeur, Doro et Shush, vraiment ravis qu’on se retrouve dans ce bâtiment) ou de continuer à développer des axes de recherches avec Jean-François et son équipe (Matthieu, Sylvain, Katia, Alain et Arlette) que se soit pour les approches en senseur, de l’automatisation de détection phénotypique ou pour trouver comment on va bien pouvoir maintenir ce foutu embryon sous la Bino. Merci à vous pour tous ces échanges et ces bons moments passés ensemble.
Une fois les premiers résultats obtenus, les collaborations s’enchaînent et se développent. Et très rapidement dans le projet des équipes de Christophe Biot et de Simon Hawkins et le magnifique BLISS que le monde nous envie grâce à Clémence Simon et Cédric Lion. C’est un plaisir de replonger avec vous dans cette interface physique-chimie-biologie.
En parallèle de ces nouveaux développements, j’ai pu revenir à mes premiers amours grâce à Gabriel Paës et «SLiM»-Christine Terryn. Merci pour ces projets passionnants et pour votre accueil exceptionnel à chacun de mes passages à Reims.
Différentes aventures hors sciences m’attendaient également en rejoignant l’UGSF. Tout d’abord, les aspects de communications qui ont pris une place importante depuis mon intégration. Un grand merci à Yann Guerardel de m’avoir confié la fonction de correspondant de communication CNRS et à Virginie Cogez, mon binôme de choc du côté universitaire. La découverte de ce nouveau domaine doit beaucoup à Stéphanie Barbez, puis à Florence Ienna et Ameline Courtois qui m’ont beaucoup aidé et soutenu dans mes projets et dont la bonne humeur et le professionnalisme rendent cette fonction aussi agréable. L’événement annuel le plus marquant de cette activité Com’ est le village des sciences et je remercie chaleureusement toute la team du stand de l’UGSF que j’ai le plaisir d’animer depuis 2017.
Une nouvelle aventure interdisciplinaire m’attendait également, à l’interface entre l’art contemporain et les sciences. Ce qui devait être une simple présentation s’est transformé en une collaboration longue est fructueuse à l’initiative de Nathalie Stefanov et Laura Mené. Merci à vous deux de m’avoir entraîné et fait découvrir ce nou-vel univers passionnant. Merci également aux artistes des projets «Cell fiction» et «Collision» avec qui j’ai eu la chance de collaborer ainsi qu’à Pierre Pauze pour toutes ces manips à la frontière de l’étrange qui offrent de nou-velles perspectives sur notre travail de recherche. Merci enfin aux collègues scientifiques qui ont rejoint l’aventure: Jean-François Bodart, Patricia Le Meillour et Simon Hawkins.
Pour conclure, merci à tous mes coauteurs, collaborateurs et utilisateurs de la plateforme pour la diversité et la richesse de mon quotidien, et à ma famille et à mes amis de sciences et d’ailleurs pour tout le reste.
Sommaire
Introduction/parcours :
9
• Liste de publications depuis la thèse: 12
Articles répertoriés (31) 12 Articles NR (6) 15 Chapitre d’ouvrage (2) 15 Brevets et licences (6) 15 • Productions artistiques 17 Expositions 17 Conférence invitée 17 • Enseignement 18
• Communication, responsabilités collectives et valorisation 22
Conférences 22
• Rayonnement scientifique et impact socioculturel 24
Fête de la science 24
• TISBio : Traitement de l’image et du signal pour la biologie 25
Prestation de service 25
Développements méthodologiques 26
Description synthétique des systèmes de microscopie disponibles 27
Dynamique et interactions moléculaires
31
• Introduction 32 • FLIM/SLiM 35
Description générale du système d’acquisition 35
Caractérisation du système 39
Étude de l’hétérodimère GABAB dans les neurones de souris 40
• FCS 45
Introduction 45
Topologie des temps de diffusion en cellule vivante 47
Corrélation entre la densité chromatinienne et la diffusion d’oligomères de eGFP 49
• Conclusion et perspectives 50
Couplage SLiM/FCS 50
Nouvelle procédure d’analyse des données de SLiM 50
Étude dans un contexte d’autofluorescence systématique 53
Les biosenseurs FRET encodés génétiquement :
57
• Introduction 58Les biosenseurs rapporteurs d’activité kinasiques 59
Définition de l’intervalle dynamique 61
• Méthodologie 63
Stratégie d’acquisition 63
Stratégie d’analyse des données ratiométriques 65
• Application : suivi dynamique de l’activité PKA 67
Mesures d’activité PKA au cours du cycle cellulaire 67
L’activité de PKA est sous localisée pendant les différentes étapes de la mitose 69
• Perspectives : les biosenseurs pour étudier les interconnections entre réseaux de régulation. 72
Contexte 72
Perspectives 76
Intelligence artificielle et images biologiques
79
• Introduction 80
La segmentation basée sur l’intensité de fluorescence 80
Le « machine Learning » 82
• Problématique biologique : étude in situ de grains d’amidon 85
Introduction et limites des méthodes actuelles 85
Mesures en fluorescence négative 86
• Machine Learning pour la Segmentation automatique de grains d’amidon 88
Première approche: le « Trainable WEKA segmentation » ou TWS 89
Perspectives: Deep learning 93
Bibliographie 98
Annexes: 105
• FLIM/SLIM/FCS 106 • Biosenseurs 106 • Traitement d’images 106 • Autres projets 107Issu d’une maîtrise de biochimie, j’ai eu l’occasion de découvrir les réponses que pouvait apporter la microscopie photonique à l’étude des fonctions du vivant. J’ai donc entrepris un mas-ter puis une thèse en physique (instrumentation et analyse avancée) au sein de l’IRI (Institut de Recherche Interdisciplinaire). J’ai alors pu appliquer ces connaissances à l’étude des interactions moléculaires dans la cellule vivante. J’ai ensuite effectué un postdoctorat en biophysique dans le laboratoire de biophysique des macromolécules, ce qui m’a apporté de nouvelles compétences pour l’étude de la dynamique moléculaire, mais également une approche plus biophysique de ces problématiques.
Mon recrutement au CNRS en tant qu’ingénieur de recherche se situe dans cette continuité et m’a permis de développer de nouvelles stratégies d’instrumentation et d’analyse pour l’étude de la dynamique et des interactions moléculaires en cellules vivantes. J’ai enfin eu, en 2012, la possibilité de créer le plateau TISBio de traitement de l’image et du signal pour la biologie dans le but de pouvoir apporter un point de vue interdisciplinaire aux problématiques de nos collabo-rateurs, tout en développant de nouvelles stratégies en adéquation avec leurs besoins.
Mon parcours m’a donc permis de m’intéresser à un grand nombre de problématiques à la fois biologiques et technologiques. J’ai donc choisi d’illustrer mon activité par différents exemples représentatifs en instrumentation, en biologie, et en analyse d’images. Une première partie présentera les développements en instrumentation et en analyse appliqués à l’étude de la dynamique et des interactions moléculaires dans le vivant. Je développerai ensuite la mise en place de stratégies de mesure d’activité kinasique dans la cellule vivante développées en étroite collaboration avec le Professeur Jean-François Bodart. Enfin, je présenterai notre stra-tégie d’intelligence artificielle pour la biologie. En parallèle de ces travaux, vous trouverez dif-férents «focus» qui sont des introductions à difdif-férents concepts utiles à la compréhension de ce manuscrit.
Introduction/parcours :
INFORMATION PERSONNELLE:
Nom: Spriet Corentin D.d.N: 10 juillet 1979
Web: http://tisbio.wix.com/tisbio Tel: 33 (0) 3 62 53 17 12
Mail: [email protected]
FORMATION INITIALE:
Doctorat Instrumentation et analyse
avancée (SCL) 2003-2006
Master Physiques/SPI (Major) 2002-2003
LM Biochimie 1998-2002
EXPERIENCE: STATUT ACTUEL : IR2 EN BIOPHOTONIqUE CNRS
TISBio@UGSF et FRABio
2013-Villeneuve-d’Ascq
Responsable Plateau TISBio
À l’interface entre la biologie, l’informatique et la physique, le plateau TISBio accompagne les projets des biologistes, de la conception des expériences et des contrôles jusqu’à l’analyse et l’interprétation des don-nées et les développements logiciels associés.
BICeL-Lille1-HB@FRABio 2012-2016
Villeneuve-d’Ascq
Responsable plateforme
La plateforme BICeL Lille1-IRI est composée de systèmes de proximité, mais également de systèmes originaux permettant : i) les mesures d’acti-vité de protéines et d’interactions moléculaires dans la cellule vivante. ii) l’imagerie 3D dans les tissus, iii) le tracking de cellules très longue durée. BCF@IRI
2010-2012
Villeneuve-d’Ascq
Ingénieur en instrumentation
Projet: développement de nouvelles stratégies de microscopie multi-modale pour l’étude de la régulation transcriptionnelle dans la cellule vivante
BoM@DKFZ 2007-2009 Heidelberg
Post-doctorant en Biophysiques
Projet: étude biophysique de la diffusion dans le noyau de la cellule vi-vante par FCS/FCCS
Profil:
Bonjour!Je suis tou-jours à la recherche de nouvelles collabora-tions, donc n’hésitez pas à me contacter ou à suivre mon actua-lité:
En bref:
Spécialités: Analyse d’images Biophotonique
Dynamiques et interactions moléculaires
Production : 48 publications 5 licences logiciels 2 brevets
1 deutsches patentamt Organisation, Didactique et activités parascientifiques:
Enseignement universitaire, encadrement, reviewing, vulgarisation Organisation de formations et écoles nationales
Porteur ou associé à des demandes de financement (ANR, BqR...) Responsable national «axe FLIM» du RTmfm (2008-2013)
CV détaillé
table des matières
Liste de publications depuis la thèse:
10
• Articles répertoriés (31) 10
• Articles NR (6) 12
• Chapitre d’ouvrage (2) 13
• Brevets et licences (6) 13
Liste non exhaustive d’encadrement
14
Productions artistiques
15
• Expositions 15
• Conférences invitées 15
Enseignement 16
Participation régulière à la formation universitaire 18
Participation régulière à des formations CNRS/INSERM 18
Participation régulière à des actions de vulgarisation scientifique 18
Communication, responsabilités collectives et valorisation
19
Participation aux réseaux nationaux 19
• Conférences 19
Conférences invitées 19
Conférence internationales 20
Poster 20
Rayonnement scientifique et impact socioculturel
21
• Fête de la science 21
TISBio : Traitement de l’image et du signal pour la biologie
22
• Prestation de service 22
Mise en place de stratégies de biophotonique 22
• Développements méthodologiques 22
• Description synthétique des systèmes de microscopie disponibles 23
Loupe Binoculaire, LEICA 23
Vidéomicroscope, LEICA AF6000X 23
Microscope Confocal Droit, Nikon A1R 24
Biostation, Nikon 24
Liste de pubLications depuis La thèse:
• Articles répertoriés (31)
Visualizing Lignification Dynamics in Plants with Click Chemistry: Dual Labeling is BLISS! C. Simon, C. Spriet, S. Hawkins, C. Lion.
Jove (2018)
The Chlamydomonas mex1 mutant shows impaired starch mobilization without maltose accumu-lation.
J. Findinier , H. Tunçay , M. Schulz-Raffelt , P. Deschamps ,C. Spriet , JM. Lacroix, T. Duchêne , N. Szydlowski ,Y. Li-Beisson , G. Peltier , C. D’Hulst , F. Wattebled , D. Dauvillée.
J Exp Bot. (2017)
BLISS: Shining a light on lignification in plants.
C. Simon, C. Lion, B. Huss, AS. Blervacq, C. Spriet, Y. Guerardel, C. Biot, S. Hawkins. Plant Signal Behav. (2017)
Manganese-induced turnover of TMEM165.
Potelle S, Dulary E, Climer L, Duvet S, Morelle W, Vicogne D, Lebredonchel E, Houdou M, Spriet C, Krzewinski-Recchi MA, Peanne R, Klein A, DE Bettignies G, Morsomme P, Matthijs G, Mar-quardt T, Lupashin V, Foulquier F.
Biochem J. (2017)
BLISS: A Bioorthogonal Dual-Labeling Strategy to Unravel Lignification Dynamics in Plants C. Lion, C. Simon, B. Huss, AS. Blervacq, L. Tirot, D. Toybou, C. Spriet, C. Slomianny, Y. Guerar-del, S. Hawkins, C. Biot
Cell ChemicalBiology (2017)
Glycosylation abnormalities in Gdt1p/TMEM165 deficient cells result from a defect in Golgi manganese homeostasis.
Potelle S, Morelle W, Dulary E, Duvet S, Vicogne D, Spriet C, Krzewinski-Recchi MA, Morsomme P, Jaeken J, Matthijs G, De Bettignies G, Foulquier F.
Hum Mol Genet. (2016)
Shadow Technique Algorithm (STA) Sheds a New Light on Differential Interference Contrast (DIC) Microscopy
Trinel D ,Vandame P, Hervieu M, Floquet E, Aumercier M, Biondi EG , Bodart JF and Spriet C* Analytical & Bioanalytical Techniques (2015)
Genetically-encoded Biosensor for Kinases: To See is to Believe? Vandame P, Trinel D, Bodart JF and Spriet C*
Biosensors Journal (2015)
Lipid nanocapsules as a new delivery system in copepods: Toxicity studies and optical imaging Stancheva S, Souissi A, Ibrahim A, Barras A, Spriet C, Souissi S and Boukherroub R
Novel Reporter for Faithful Monitoring of ERK2 Dynamics in Living Cells and Model Organisms Sipieter F, Cappe B, Gonzalez Pisfil M, Spriet C, Bodart JF, Cailliau-Maggio K, Vandenabeele P, Héliot L, Riquet F
PLoS One (2015)
The spatio-temporal dynamics of PKA activity profile during mitosis and its correlation to chro-mosome segregation
Vandame P, Spriet C, Trinel D, Gelaude A, Caillau K, Bompard C, Biondi E, Bodart JF. Cell Cycle (2014)
Optimization of ERK activity biosensors for both ratiometric and lifetime FRET measurements. Vandame P, Spriet C, Riquet F, Trinel D, Cailliau-Maggio K, Bodart JF.
Sensors (2014)
The level of Ets-1 protein is regulated by poly(ADP-ribose) polymerase-1 (PARP-1) in cancer cells to prevent DNA damage.
Legrand AJ, Choul-Li S, Spriet C, Idziorek T, Vicogne D, Drobecq H, Dantzer F, Villeret V, Au-mercier M. PLoS One 8, (2013).
Impairment of GABAB receptor dimer by endogenous 14-3-3zeta in chronic pain conditions. Laffray S, Bouali-Benazzouz R, Papon MA, Favereaux A, Jiang Y, Holm T, Spriet C, Desbarats P, Fossat P, Le Feuvre Y, Decossas M, Héliot L, Langel U, Nagy F, Landry M.
EMBO J (2012).
Generalization of the polar representation in time domain fluorescence lifetime imaging micros-copy for biological applications: practical implementation.
Leray A, Spriet C, Trinel D, Usson Y & Heliot L. J Microsc 248, 66-76, (2012).
Interleukin-7 regulates adipose tissue mass and insulin sensitivity in high-fat diet-fed mice through lymphocyte-dependent and independent mechanisms.
Lucas S, Taront S, Magnan C, Fauconnier L, Delacre M, Macia L, Delanoye A, Verwaerde C, Spriet C, Saule P, Goormachtigh G, Héliot L, Ktorza A, Movassat J, Polakowska R, Auriault C, Poulain-Godefroy O, Di Santo J, Froguel P, Wolowczuk I.
PLoS One (2012).
quantitative comparison of polar approach versus fitting method in time domain FLIM image analysis.
Leray A, Spriet C, Trinel D, Blossey R, Usson Y, Héliot L. Cytometry A 79, 149-158, (2011).
Reporting Kinase Activities: Paradigms, Tools and Perspectives
Riquet F, Vandame P, Sipieter F, Cailliau-Maggio K, Spriet C, Heliot L,Bodart JF. JBM (2011)
Short exposure to the DNA intercalator DRAq5 dislocates the transcription machinery and induces cell death.
Richard E, Causse S, Spriet C, Fourré N, Trinel D, Darzacq X, Vandenbunder B, Heliot L. Photochem Photobiol 87, 256-261, (2011).
Upgrading time domain FLIM using an adaptive Monte Carlo data inflation algorithm. Trinel D, Leray A, Spriet C, Usson Y & Heliot L.
Cytometry A 79, 528-537, (2011)
Control of pulse-to-pulse fluctuations in visible supercontinuum. Kudlinski A, Barviau B, Leray A, Spriet C, Héliot L, Mussot A. Opt Express 18, (2010).
Cell adhesion properties on chemically micropatterned boron-doped diamond surfaces. Marcon L, Spriet C, Coffinier Y, Galopin E, Rosnoblet C, Szunerits S, Héliot L, Angrand PO, Boukherroub R.
Langmuir 26, (2010).
Mapping eGFP oligomer mobility in living cell nuclei.
Spriet C & Dross N, Zwerger M, Müller G, Waldeck W, Langowski J. PLoS One 4, (2009).
Optimized protocol of a frequency domain fluorescence lifetime imaging microscope for FRET measurements.
Leray A, Riquet FB, Richard E, Spriet C, Trinel D, Héliot L. Microsc Res Tech (2009).
Three-dimensional polar representation for multispectral fluorescence lifetime imaging micros-copy.
Leray A, Spriet C, Trinel D & Heliot L. Cytometry A 75, 1007-1014 (2009).
Synthesis and application of FRET nanoparticles in the profiling of a protease. Spriet C* & Marcon L, Meehan TD, Battersby BJ, Lawrie GA, Héliot L, Trau M. Small 5, 2053-2056, (2009).
Functional cooperation between Stat-1 and ets-1 to optimize icam-1 gene transcription. Yockell-Lelièvre J, Spriet C, Cantin P, Malenfant P, Heliot L, de Launoit Y, Audette M. Biochem Cell Biol 87, 905-918, (2009).
Enhanced FRET contrast in lifetime imaging.
Spriet C, Trinel D, Riquet F, Vandenbunder B, Usson Y, Heliot L. Cytometry A 73, 745-753 (2008).
Concurrent fast and slow cycling of a transcriptional activator at an endogenous promoter. Karpova TS, Kim MJ, Spriet C, Nalley K, Stasevich TJ, Kherrouche Z, Heliot L, McNally JG.. Science 319, 466-469, (2008)
Correlated fluorescence lifetime and spectral measurements in living cells. Spriet C, Trinel D, Waharte F, Deslee D, Vandenbunder B, Barbillat J, Heliot L. Microscopy Research and Technique (2007). Egalement en couverture.
Epithelial Na+ channel stimulation by n-3 fatty acids requires proximity to a membrane-bound A-kinase anchoring protein complexed with PKA and Phosphodiesterase.
Mies F, Spriet C, Heliot L and Sariban-Sohraby S. JBC (2007)
• Articles NR (6)
Biometric data assessment on Xenopus laevis tadpoles
S. Slaby, J. Hanotel,J.-F. Bodart, S. Lemiere, D. Trinel, Leprêtre, M. Marin & C. Spriet Journal of Xenobiotics (2016)
Biological based biosensor’s inception Spriet C, Kasprowicz A, Bodart JF. J of Biosens Biomark Diagn. (2016)
Being Analytical: À Sharp End in Integrative Approaches Spriet C, Trinel D, Bodart JF
Journal of Research Analytica (2015) Embryogenesis: How to get the Picture? Spriet C, Trinel D, Bodart JF
Current Synthetic and Systems Biology (2015)
Getting ab ovo Developmental Processes Intelligible Using Metaphors, Microscopy and Para-digms.
Spriet C, Maggio K, Bodart JF
Current Synthetic and Systems Biology (2015)
FRET-Based Sensors for Kinase Activity: An increasing Attractiveness. Spriet C* and Bodart JF
Austin J Biosens & Bioelectron (2015)
• Chapitre d’ouvrage (2)
From FRET imaging to practical methodology for kinase activity sensing in living cells. Vandame P, Sipieter F, Spriet C, Leray A, Vincent P, Trinel D, Bodart JF, Riquet FB, Héliot L Prog Mol Biol Transl Sci 113, 145-216 (2013).
FRET-Based Enzyme Activity Reporter: Practical Hints for Kinases as Indicators of Virulence Spriet C, Kasprowicz A, Trinel D, Bodart JF. Biosensing Technologies for the Detection of Pa-thogens (2018)
• Brevets et licences (6)
• Sub-diffraction Imaging (SdI) (2015) • Easyquick Super Resolution (EqSR) (2015)
• Procédé et système d’analyse moléculaire à commutation de mesures d’imagerie et de spectroscopie. Brevet d’invention. Spriet C*, Laurent H, Barbillat J, 2010.
• Deutsches Patentamt n°20 2008 010 895.0, heidelberg, 2009. • TITAN n°2354-1, 2008,
Liste non exhaustive d’encadrement
Septembre 2017 - juin 2018: Angélina Kasprowicz M2. 60%Diversité des interactions des voies MAPK et OGT/OGA dans les cellules cancéreuses mam-maires, utérines et colorectales : approche quantitative en cellules uniques.
février - juin 2017: Angélina Kasprowicz M1, stage rémunéré. 60% La O-GlcNacylation module-t-elle l’activité MAPK ?
2012-2017: Encadrement de Dave Trinel 100%
IE CNRS expert en développement logiciel et déploiement d’application. 2016: Angélina Kasprowicz M1, 50%
La O-GlcNacylation module-t-elle l’activité MAPK ?
2012-2014: Encadrement technologique de Pauline Vandame, doctorante en «Biologie Santé»: 40%
Utilisation et développement de biosenseurs FRET pour la mesure d’actvités kinases in vivo au cours du cycle cellulaire (tuteurs déclarés : C. Bompard, J.F. Bodart, encadrement effectif par Corentin Spriet : 40%, attesté par six publications)
2013: Encadrement de Philippe Saudemont, étudiant en Master I« Sciences, Technologies, Santé »: 70%
Régulation de l’activité PKA au cours de la mitose
2012: Encadrement de Marie Boyer, étudiante en Master I « Biologie et Biotechnologies»: 80% Caractérisation d’un prototype permettant une mesure corrélée de la dynamique et des interac-tions moléculaires dans la cellule vivante.
2012: Encadrement de Ludovic Depraeter, étudiant en Master I «physiques médicales»: 80% Mise en place d’un système de RICS (Raster Image Correlation Spectroscopy) pour l’étude de la dynamique moléculaire en cellule vivante.
2011: Encadrement de Julie Gallet, étudiante en Master I« Biologie et Biotechnologies »: 80% Travail préliminaire à l’étude de l’interaction ARN Polymerase II / Cycline T1
2011: Encadrement technologique de Pauline Vandame, étudiante en Master II « Biologie-Santé »: 50%
Rôle de PKA sur l’activité et la localisation de MAPK/ERK au cours du cycle cellulaire dans les cellules HeLa.
2010: Encadrement Al Amine Issulahi, master II «imagerie pour la biologie de Rouen» 30% Métrologie appliquée à la microscopie photonique.
2007-2012: encadrement technologique de 3 ingénieurs d’étude plateforme
Encadrements technologiques au cours de M1 (ex : Pauline Grard, (2016), de M2 (ex : François Sipieter) et de thèse (ex : Sylvain Slaby).
productions artistiques
Coresponsable du projet « Cells Fictions #2», avec production d’un catalogue et de deux exposi-tions dans le cadre du programme de recherche Images, Sciences et Technologies.
Mené par l’École Supérieure d’Art du Nord-Pas de Calais, Dunkerque / Tourcoing (L’ESÄ), L’Espace Croisé, Centre d’Art et le laboratoire TISBio. L’objectif est de faire émerger une commu-nauté d’artistes et de scientifiques afin de réaliser ensemble des formes plastiques innovantes autour des questions du vivant, suscitant des rencontres et des interactions avec les publics. Ce programme débouche également sur la création d’un module de master transversal pour les étudiants de Polytech’Lille et de l’ESÄ.
•
Expositions
Cell Fiction # 2,
23 mars au 7 avril 2017,
Galerie commune, École supérieure du Nord Pas-de-Calais , site de Tourcoing
16 mai au 2 septembre 2017, espace culture université de Lille sciences et technologies, Villeneuve-d’Ascq
•
Conférence invitée
Comment créer les conditions adéquates pour que les artistes produisent en collaboration avec le monde académique?
FOOR : Forum Ouvert Oeuvres et Recherche (2017).
enseignement
Depuis mes débuts dans le monde de la recherche, j’ai toujours pris beaucoup de plaisir à m’investir dans différentes activités de formation en tant qu’intervenant et/ou organisateur. Mon parcours m’a donné l’occasion de réaliser ces formations pour des publics interdisciplinaires depuis la licence jusqu’à la formation pour chercheurs confirmés. Voici quelques exemples repré-sentatifs de cette activité.
• MiFoBio
2004, responsable logistique et coordinateur. 2006 et 2008 membre du comité d’organisa-tion et du comité scientifique
Période : 2004, 2006, 2008, 2010. Durée : 2h CM + 37h TP
Effectifs : ~300 pour les CM et ~20 pour les TP
Public : Étudiants chercheurs et ITA, biologistes physiciens et chimistes Disciplines : Biologie et physique
Contenu : École thématique interdisciplinaire de microscopie fonctionnelle du vivant. Cours : « étude des interactions moléculaires en cellules vivantes », Table ronde : « mesure d’interactions entre molécules par FRET : FLIM, Phase et modulation, Polarisation... »TP: « FRAP nucléaire », « Techniques de FLIM », « Interactions moléculaires », « initiation FLIM/FRET », « dynamique et interactions moléculaires », « microscopie multimodale FLIM/SLiM »
Figur e 1 : É cole thémati que MiF oBio 2008, Carqueiranne.
• Teck’FRET, Teck’FLIM, BIAT
Organisateur/coorganisateur. Période : 2006, 2008.
Durée : 7h CM + 17h TD
Effectifs : ~15 pour les CM et ~4 pour les TD
Public : Chercheurs et ITA, biologistes et physiciens Disciplines : Instrumentation et physique
Contenu : Formation technologique aux mesures de FRET et aux études de temps de vie par TCSPC. Application à l’étude d’interactions moléculaires en cellules vivantes. Cours : « Sys-tème de mesure de temps de vie par TCSPC et applications. », « Interactions dans les tissus : apport de la microscopie multimodale », « Outils mathématiques et procédures d’analyses pour l’étude des interactions. » TD : « FRET par mesure de FLIM (TCSPC) », « Microscopie multimodale pour les mesures d’interactions en animal vivant », « FRET transversal (TCSPC/Phase et modula-tion/ratio d’intensité) »
• Tutorat de microscopie à destination des personnels non francophones
Organisateur
Période : 2013-2016 Durée : 8h CM + 8h TP
Effectifs : ~30 pour les CM et ~4 pour les TP
Public : doctorants et post-doctorants, biologistes et chimistes Disciplines : Biologie et physique
Contenu : Vidéomicroscopie, microscopie confocale et analyse d’image Cours et TP : « Introduction to microscopy », « Introduction to confocal microscopy » « Image analysis using imageJ » « Automation using imageJ »
• Exploration du vivant: du Micron à l’Angström
Organisateur
Période : 2012 - présent Durée : 8h
Effectifs : ~40
Public : enseignants de collèges et lycées. Toutes disciplines. Disciplines : Biologie et physique
Nano-Ecole Lille afin de vulgariser les nanotechnologies et le « nano monde » pour les ensei-gnants.
• Vidéomicroscopie et microscopie confocale
Organisateur
Période : 2005 - présent Durée : 6h CM + 48h TP
Effectifs : ~40 pour les CM et ~4 pour les TP Public : Etudiants chercheurs et ITA, biologistes Disciplines : Biologie et physique
Contenu : Vidéomicroscopie et microscopie confocale : des bases à la pratique, du tissu à la molécule unique. Cours : « Les bases de la microscopie : introduction à l’optique », « Introduc-tion à la microscopie confocale ». TP : « microscopie confocale II : acquisition multiparamétrique et colocalisation ».
• Interface physique biologie et régulation transcriptionnelle
Organisateur
Période : 2004, 2005, 2006. Durée : 4h CM
Effectifs : ~20
Public : Etudiants Master 2 et IUP, biologistes. Disciplines : Biologie et physique
Contenu : Module universitaire développant l’apport de la microscopie photonique à l’étude du vivant et en particulier à la compréhension de la régulation transcriptionnelle. Cours : « Interface physique biologie : régulation transcriptionnelle », « Interface physique biologie : Les F-techniques », « Nouveaux outils pour l’étude d’interactions moléculaires à l’intérieur du noyau ».
Formation universitaire
Enseignement de la Biophotonique à l’Université de Lille et dans le cadre du Master européen EMMI.
Cours, TP, TD. Niveau IUP, Master I et II de physiques et de biologie.
Formations CNRS/INSERM
Enseignement allant d’introduction à l’optique jusqu’à des cours avancés comme les mesures d’interactions entre molécules par FRET: FLIM, Phase et modulation, Polarisation...
Actions de vulgarisation scientifique
Présentation des métiers de la recherche, salon des métiers, formation à destination des ensei-gnants du secondaire...
• Exemples d’enseignements donnés
“introduction à la microscopie” M2 master «Chimie et Sciences du Vivant», 2015- , Lille .
«Formation pour les doctorants : que faire de vos données de recherche». 2016- , Lille.
License III/Master I-II in physics and biology Université Lille 1 (2008-ongoing ~15H /year) Introduction à la microscopie/microscopie confocale
INSERM/CNRS Teaching. Lille, Bordeaux (2008-ongoing ~50 H/Year)
Second and Third Harmonic Generation for biological imaging
European Master in Molecular Imaging (2010-2013 ~3H/Year), Université Paris Sud
« Journée thématique MB2: Dynamique et robustesse des réseaux de régulation cellulaire » 2008-2012, Lille
communication, responsabiLités coLLectives et
vaLori-sation
En plus de mon activité scientifique et d’enseignement, j’ai eu l’occasion de participer à différentes actions de communication grand public (présentation de plateforme, salon des collé-giens, posters de vulgarisation) et scientifique, notamment au moment de la création de l’Institut de Recherche Interdisciplinaire (conférence pédagogique sur la nécessité de l’interdisciplinarité et la place de la biophotonique dans ce contexte) ou pour la mise en place de collaborations ou de projets européens. Dans ce contexte, j’ai également pu participer à la rédaction et obtenir des demandes de fonds pour des projets Feder, ANR, BQR et Intereg.
Un élément récurant de mon parcours étant le développement et le transfert de nouvelle technologie et procédures d’analyse, j’ai pu participer à la valorisation de ces dernières. Pour ce faire, j’ai eu un rôle régulier de formation dans le cadre de l’activité plateforme des différents instituts au sein desquels j’ai travaillé (IBL, IRI, DKFZ, UGSF) afin de rendre accessible les diffé-rentes technologies présentes (vidéomicroscopie, traitement d’image, déconvolution, microsco-pie confocale, FRAP, FRET, FCS). J’ai également participé à la valorisation du système de mesure de SLiM développé au cours de ma thèse avec la fabrication d’un deuxième prototype installé à l’ENS Cachan. J’ai également coordonné un brevet couvrant ce système.
J’ai été fortement impliqué dans la création du groupement de recherche « microscopie fonctionnelle du vivant » (GDR2588) et du réseau technologique de « microscopie photonique de fluorescence multidimensionnelle » (RT mfm). J’ai donc pu participer activement à la création et à l’animation de ces deux groupements, notamment par différentes animations scientifiques et par des mobilités régulières au sein des laboratoires membres afin d’aider à la mise en place de technologies de pointe (FLIM par TCSPC, SLiM, Cavity dumper, FRET pour les mesures en sen-seurs…)
J’ai eu l’occasion de présenter mon travail de recherche à différentes communautés (bio-logistes, physiciens, interdisciplinaires) au cours de différentes conférences nationales (congrès de l’Association française de cytométrie, congrès de la société de biologie cellulaire française …) et internationales (Focus On Microscopy, European Light Microscopy Initiative, first congrès of the fédération Réaumur des sciences du vivant…), notamment en tant que conférencier invité en moyenne 2 fois par an.
Participation aux réseaux nationaux
• Responsable national «axe FLIM» du RTmfm(2008-2013) • Membre du GDR Microscopie et Imagerie du Vivant (MIV)
• Membre du RT Microscopie Photonique de Fluorescence Multidimensionnelle (mfm) • Membre du conseil de laboratoire de L’IRI (2011-2014)
• Conférences
Participation régulière à des séminaires et conférences, notamment:
Conférences invitées
Le FRET par TCSPC, pourquoi et comment ? Corentin spriet, Dave Trinel
Introduction aux « F-techniques » : Dynamiques et interactions moléculaires dans le vivant. 5e Journées scientifiques et techniques des microscopistes INRA 2014
François Waharte, Dave Trinel, Corentin Spriet
Super continuum laser source for time correlated single photon counting with adjustable excita-tion wavelength.
Corentin Spriet, Aymeric Leray, Dave Trinel, Franck Riquet, Laurent Héliot Webcast Leica 2012
Conférence internationale
Fluorescence Lifetime Imaging Microscopy Corentin Spriet
NICAC 2011, Heidelberg, Germany
Combined FLIM and FCS approach to quantify dynamic and interaction during transcription early elongation phase.
Corentin Spriet, Nicolas Dross, Elodie Richard, Aymeric Leray, Olivier Bensaude, Xavier Dar-zacq, Jörg Langowski, Laurent Heliot.
ELIM 2010, Heidelberg, Germany
Fluorescence lifetime and spectral measurements to determine protein-protein interactions in cells.
Corentin Spriet, Sophie Laffray, Dave Trinel, Didier Deslee, Bernard Vandenbunder, Jacques Bar-billat, Laurent Heliot
FOM2007, Valencia, Espagne
Poster
More FRET contrast in lifetime imaging
Corentin Spriet, Dave Trinel, Bernard Vandenbunder, Laurent Heliot. ELMI 2006, OFIR , Portugal
Multimodal microscopy for transcription regulation studies
Corentin Spriet, Nicolas Dross, Elodie Richard, Aymeric Leray, Olivier Bensaude, Xavier Dar-zacq, Jörg Langowski, Laurent Heliot.
FOM2009, Krakow, Pologne
Spectrally-resolved fluorescence lifetime measurements to determine protein-protein interactions in living cells.
Corentin Spriet, François Waharte, Dave Trinel, Didier Deslee, Bernard Vandenbunder, Jacques Barbillat, Laurent Heliot
rayonnement scientifique et impact sociocuLtureL
Au-delà de son implication dans l’enseignement universitaire et dans la formation des différents personnels de l’institut, le plateau TISBio participe également à la visibilité de l’UGSF auprès du grand public, que ce soit :
- vers les scolaires, notamment par l’organisation de la journée « Exploration du vivant: du Micron
à l’Angström » du Plan d’Action et de Formation de Professeurs de collèges et de lycées (collabora-tion IEMN/IRCICA et C.Bompart de l’équipe glycobiologie végétale)
- vers un public scolaire/ grand public avec l’organisation de la fête de la science avec les équipes
« Régulation de la glycosylation terminale » et « Régulation des signaux de division »
- ou vers le grand public, avec le développement, en partenariat avec l’École Supérieure d’Art et
l’espace Croisé, d’un axe de recherche « Images, sciences et Technologies ».
Dans la logique de cette implication, j’ai pris en charge la responsabilité de la communication de l’UGSF avec le CNRS depuis 2017.
• Fête de la science
TISBio s’est fortement investi dans l’organisation et la coordination du stand de l’UGSF pour la fête de la science. Cet événement a été l’occasion de mettre en place des partenariats avec d’autres acteurs de la région comme le fabricarium de Polytech’Lille. L’implication des dif-férents animateurs de l’institut a donné à notre stand un fort retentissement auprès du grand public et a été l’objet d’articles de blogs de vulgarisation scientifique renommés, ainsi que dans « la Voix du Nord » ou wéo, chaîne de télévision locale des hauts de France.
Figure 2: fête de la science 2016 avec le stand de l’UGSF mis à l’honneur dans l’article « Rencontrez
peut-être le futur prix Nobel pendant la fête de la Science » de la Voix du Nord (A), le kit pédagogique (B) et le jeu sur le développement embryonnaire réalisé pour l’occasion en partenariat avec l’équipe de « régulation
tisbio : traitement de L’image et du signaL pour La
bio-Logie
Nous avons fondé en 2013, avec Dave Trinel, le plateau TISBio que je dirige depuis lors. Il a pour objectif de rendre la biophotonique accessible aux biologistes. L’étude du vivant par la microscopie photonique peut être difficile à appréhender, car l’analyse des données nécessite des connaissances pluridisciplinaires. À l’interface entre la Biologie, l’Informatique et la Physique, TISBio accompagne les projets des biologistes, de la conception des expériences et des contrôles jusqu’à l’analyse et l’interprétation des données et les développements logiciels associés. TISBio gère également un parc de microscopes photoniques. Il est composé de systèmes de proximité, mais également de systèmes originaux et attractifs pour les utilisateurs de la région. Les activi-tés, hors recherche et développement, du plateau sont présentées ci après.
• Prestation de service
Mise en place de stratégies de biophotonique
Nous proposons les méthodes de biophotoniques les mieux adaptées aux problématiques des utilisateurs de la plateforme. À titre d’exemple, l’équipe « Glycobiologie végétale » est deve-nue utilisatrice régulière de la plateforme (Fig. 3) et des problématiques plus complexes nous amènent désormais à collaborer de façon très étroite.
Figure 3: vidéomicroscopie de chloroplastes de Chlamydomonas exprimant la protéine GFP seule (A) ou fusionnée à la protéine Mex (E). (B) et (F) correspondent à la fluorescence de la chlorophylle et (C) et (G)
• Développements méthodologiques
Notre rôle consiste également à optimiser les stratégies de biophotonique de ses utili-sateurs, que ce soit par le choix des meilleures procédures d’acquisition à adopter ou par le développement de stratégies d’analyse adaptées à leur besoin. À titre d’exemple, on peut citer l’équipe « Mécanismes moléculaires de la N-glycosylation … » qui, dans son étude des CDG (Congenital disorders of glycosylation), a toujours accordé une place importante à l’imagerie et notamment à l’étude de la sous-localisation de protéines impliquées dans ces glycosylations aberrantes. Cependant, aucune stratégie de segmentation automatique n’avait été mise en place, ce qui excluait toute analyse quantitative. Nous avons donc développé un plug-in ImageJ permettant de quantifier la répartition de protéines fluorescentes dans les vésicules et l’appareil de Golgi (Fig. 4). Ce plug-in est désormais utilisé en routine au laboratoire.
Figure 4: déve-loppement d’un plug-in pour ImageJ permettant la seg-mentation automa-tique de protéines dans l’appareil de golgi et les vésicules. Taille de l’image, 3 9 0 µ m * 3 9 0 µ m (Potelle et al., 2016,
• Description synthétique des systèmes de microscopie
dispo-nibles
Loupe Binoculaire, LEICA
• Spécificités instrumentales- Équipée d’une caméra couleur - Équipée pour la fluorescence - Imagerie multi-Échelle
• Développement de stratégies d’analyse
- Imagerie multi échelle d’objets millimétriques
- Analyse morpho métrique du développement embryonnaire (Fig.1.A)
Vidéomicroscope, LEICA AF6000X
• Spécificités instrumentales- Imagerie de fluorescence : localisation, colocalisation sur des échantillons fixés et vivants - Imagerie en transmission : contraste de phase, contraste interférentiel (DIC), polarisation - Imagerie multidimensionnelle : tridimensionnelle (XYZ), longueur d’onde (λ), temps (t) - Imagerie rapide en fluorescence et transmission
- Imagerie régulée en température et en CO2
• Développement de stratégies d’analyse
- Imagerie dynamique d’activité de protéines par biosenseurs basés sur le FRET - Tracking d’organites subcellulaires
- Visualisation 3D d’imagerie DIC et analyse morpho métrique (Fig.1.B, C)
Microscope Confocal Droit, Nikon A1R
• Spécificités instrumentales- Imagerie de fluorescence : localisation et colocalisation sur échantillons fixés et vivants - Imagerie multidimensionnelle : tridimensionnelle (XYZ), longueur d’onde (λ), temps (t)
- Imagerie spectrale : détecteur 32 canaux
- Possède une platine fixe dédiée à la recherche en électrophysiologie
• Développement de stratégies d’analyse
- Reconstruction 3D (Fig.1.F)
- Imagerie spectrale d’autofluorescence
- Segmentation 2-3D et « Machine learning » (Fig.1.D)
Biostation, Nikon
• Spécificités instrumentales- Imagerie de fluorescence : localisation, colocalisation sur des échantillons vivants - Imagerie en transmission : contraste de phase et polarisation
- Imagerie multidimensionnelle : tridimensionnelle (XYZ), longueur d’onde (λ), temps (t) - Imagerie régulée en température et en CO2
- Grande simplicité d’utilisation
• Développement de stratégies d’analyse
- Création de mosaïque d’images 3D
- Tracking longue distance et longue durée (Fig.1.E)
- Analyse de réparation de blessure et de prolifération/apoptose
Microscope Droit
- Imagerie de fluorescence : localisation et colocalisation sur échantillons fixés et vivants
- Imagerie en transmission : contraste de phase, contraste interférentiel (DIC), pola-risation
Figure 5:exemples d’applications TISBio. A) analyse morphométrique de la tête d’un embryon de Xénope
(J.F.Bodart, UGSF). B) Segmentation et analyse du rayon de courbure bactérienne (E.Biondi, LCB). C) recons-truction d’imagerie DIC 3D de racine de lys (TISBio). D) Reconnaissance automatique et quantification de fluorescence dans l’appareil de Golgi et les vésicules (F.Foulquier, UGSF) E) tracking de cellules cancéreuses, sur une reconstruction en mosaïque (L. Vyacheslav, PhyCell). F) Imagerie 3D de l’autofluorescence du
tabLe des matières
Introduction 32
FLIM/SLiM 35
• Description générale du système d’acquisition 35
• Caractérisation du système 39
• Étude de l’hétérodimère GABAB dans les neurones de souris 40
Signature SLiM et autofluorescence neuronale 40
Étude de l’impact de la protéine 14-3-3ζ sur l’hétérodimerisation du récepteur GABAB 44
FCS 45
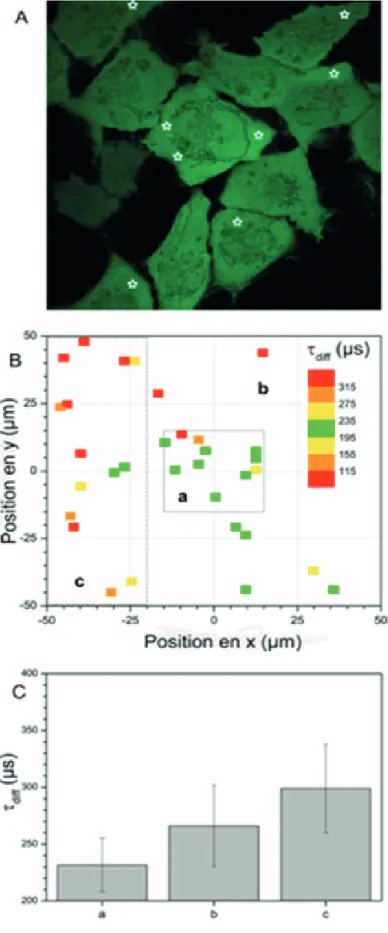
• Introduction 45• Topologie des temps de diffusion en cellule vivante 47
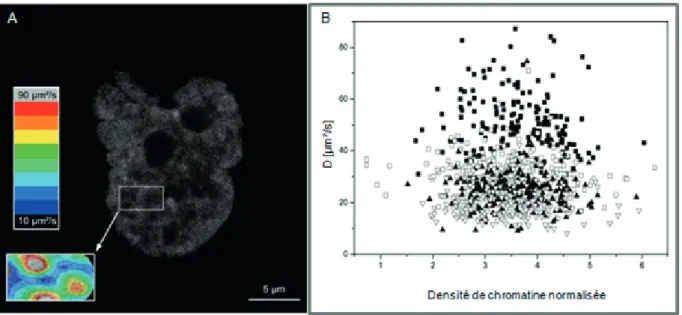
• Corrélation entre la densité chromatinienne et la diffusion d’oligomères de eGFP 49
Conclusion et perspectives
50
• Couplage SLiM/FCS 50
• Nouvelle procédure d’analyse des données de SLiM 50
• Étude dans un contexte d’autofluorescence systématique 53
• Perspectives 54
Dynamique et
interactions moléculaires
introduction
La cellule est un système tridimensionnel complexe et dynamique. L’ensemble de ses fonc-tions est régulé par des complexes multiprotéiques. L’étude de la formation de ces complexes dans la cellule vivante est donc une question rencontrée de façon récurrente en biologie. Dif-férentes techniques existent pour étudier les interactions moléculaires, aussi bien in vitro qu’in
vivo, par des techniques physico-chimiques ou microscopiques. L’émergence des protéines GFP,
popularisées dans les années 1990 par Roger Tsien (Tsien, 1998, 2005), a grandement participé à l’essor de ces dernières (prix Nobel de chimie 2008 décerné à Osamu Shimomura, Martin Chalfie et Roger Y. Tsien pour la découverte et l’utilisation de la GFP).
En particulier, les mesures de temps de vie de fluorescence par comptage de photons uniques corrélés dans le temps (TCSPC,Becker et al., 2004) permettent une analyse quantitative des interactions moléculaires par l’étude du transfert d’énergie non radiatif entre deux fluoro-phores distants de quelques nanomètres (FRET). L’utilisation d’un système de mesure de temps de vie précis et caractérisé de façon rigoureuse (Waharte et al., 2006) permet alors l’étude des interactions moléculaires en marquant deux protéines d’intérêt par des fluorophores appropriés (Camuzeaux et al., 2005; Karpova et al., 2008; Mies et al., 2007).
Les mesures de temps de vie de fluorescence en cellule présentent cependant certaines contraintes quant à la complexité des courbes de décroissance obtenues et à la faible statistique de photons disponibles pour réaliser leurs ajustements. La fluorescence présentant d’autres caractéristiques, il était prévisible que le « couplage corrélatif des axes de mesures » allait ap-porter une complémentarité de l’information permettant de combler les manques de chacune des techniques. Le spectre de fluorescence est une dimension complémentaire particulièrement intéressante pour l’étude du FRET.
Au cours de mon travail de doctorat au sein de l’équipe d’ « imagerie cellulaire fonction-nelle » de L’Institut de Biologie de Lille (2003-2006, Lille, France), j’ai entrepris le développe-ment et la caractérisation d’un système permettant les mesures corrélées de spectre et de temps de vie de fluorescence ou SLiM. J’ai également développé des stratégies d’analyse permettant l’exploitation des données de SLiM pour l’étude des interactions moléculaires.
Les études de SLiM améliorent la capacité d’analyse des interactions moléculaires. Cepen-dant, la détection d’interactions par mesure de FRET peut avoir des significations biologiquement très différentes. La FCS apporte des informations sur la vitesse de diffusion et la concentration des protéines (Dittrich et al., 2001; Módos et al., 2004). Il est alors possible de déterminer si deux protéines sont libres, incluses dans un complexe multiprotéique ou fixées à des éléments structuraux de la cellule comme la chromatine. Associées aux études de FRET, les études de FCS permettent d’atteindre une bien meilleure compréhension des mécanismes biologiques à l’œuvre.
Mon séjour postdoctoral dans l’équipe « biophysics of macromolecules » (2007-2009, DKFZ, Heidelberg) m’a permis de quantifier la mobilité de protéines en fonction de leur masse moléculaire (oligomères de eGFP) et de la densité de chromatine (marqueurs d’histones). Pour ce faire, j’ai développé un protocole de caractérisation adapté aux mesures topologiques de FCS dans le noyau des cellules vivantes.
Focus sur la fluorescence
La fluorescence est un outil pré-cieux pour l’étude du vivant grâce à la faible « invasivité » et la spécificité de marquage qu’elle permet d’obtenir. La première observation empirique de la fluorescence réalisée sur la quinine en 1845 par Sir John Frederick Wil-liam Herschel (Herschel, 1845) n’a pu être expliquée avec les connaissances scientifiques de l’époque. De nos jours, les phénomènes photo physiques à l’œuvre sont beaucoup mieux connus.
Pour émettre de la fluorescence, une molécule doit posséder au moins un électron délocalisé. Lorsque cette molécule absorbe un photon, elle passe de l’état électronique fondamental à un état électronique excité. La fluores-cence est un des modes permettant de revenir à l’état fondamental par l’émis-sion d’un photon. Perrin et Jablonski ont proposé une représentation des différents niveaux énergétiques et des sauts entre ces niveaux associés aux phénomènes de fluorescence.
Un diagramme de Perrin-Jablonski typique est présenté ci-dessus. Les sauts énergétiques entre les états singulets S0 (l’état fondamental), S1 et S2 (états excités) sont quantiques, mais il existe différents niveaux vibra-tionnels entre chacun de ces états. La fluorescence correspond à un saut énergétique de l’état S1 à un des niveaux vibrationnels de l’état fondamental.
Il existe une grande diversité de molécules fluorescentes. Le choix d’une molécule pour une expérience donnée sera défini selon trois grands critères :
- Son efficacité en tant que molécule fluorescente, - Les propriétés de la fluorescence émise,
- Sa sensibilité quant à son environnement.
Efficacité des sondes fluorescentes
Le phénomène de fluorescence requiert l’absorption d’un ou plusieurs photons suivie de l’émission d’un photon d’énergie inférieure. La première caractéristique d’un fluorophore est donc sa capacité à absorber un photon. Elle est définie par le coefficient d’absorption molaire, ε (M-1.cm-1), qui relie, pour une longueur d’onde donnée, la quantité de lumière absorbée à la concentration de fluorophores en solution.
Une fois que la molécule a absorbé l’énergie d’un photon incident et qu’elle passe dans un état excité, différents modes de déssexcitation sont possibles. Parmi ceux-ci, on peut citer une transformation photochi-mique, un changement de conformation, une conversion interne, l’émission d’un électron ou d’un proton, ou un transfert d’énergie qui pourra être radiatif ou non. Le phénomène de fluorescence n’est donc qu’un des nombreux modes de desexcitation possible. Un fluorophore sera donc caractérisé par son rendement quan-tique qui définit l’efficacité relative de la fluorescence par rapport aux autres voies de désexcitation. Il est exprimé par Φ, le ratio entre le nombre de photons émis sur le nombre de photons absorbés.
Enfin, parmi les modes de desexcitation possibles, nous avons mentionné la possibilité d’obtenir des réac-tions photochimiques. Parmi celles-ci, la réaction avec l’oxygène conduisant à la formation de radicaux libres fera perdre au fluorophore sa capacité à émettre des photons. Au fur et à mesure de l’excitation d’un ensemble de fluorophores, une proportion de plus en plus grande sera photo-blanchie et ne sera donc plus observable.
Un fluorophore idéal aura donc une forte capacité à absorber puis à ré-émettre des photons tout en res-tant stable au cours du temps.
Diagramme de Perrin-Jablonski représentant la phase d’excitation (1), de relaxation vibrationnelle (2) et de désexcitation accompagnée de l’émission d’un photon de fluorescence (3). A : D’un point de vue énergétique ; B : D’un
Propriété de la fluorescence émise
Deux grandes caractéristiques de la fluorescence peuvent être déduites du diagramme de Perrin-Jablonski et pourront être utilisées pour observer de façon spécifique un fluorophore :
Une partie de l’énergie reçue par le fluorophore sera dissipée dans les états vibrationnels intermé-diaires de l’état excité. La longueur d’onde étant inver-sement proportionnelle à l’énergie, le spectre d’émis-sion aura toujours une longueur d’onde supérieure au spectre d’excitation. Ce décalage entre le mum de la première bande d’absorption et le maxi-mum d’émission est appelé décalage de Stockes. En regardant précisément les spectres de fluorophores, on remarque cependant un chevauchement entre les spectres d’excitation et d’émission de la majorité des fluorophores. La différence d’énergie provient de la chaleur ambiante qui permet à une molécule dans l’état S0 d’être dans un état vibrationnel excité. L’éner-gie nécessaire pour passer à l’état excité sera donc plus faible, ce qui explique ce chevauchement (Valeur, 2002). Il est à noter que l’énergie pour passer d’un
état singulet à un autre est très supérieure à celle nécessaire à un changement d’état vibrationnel. La probabi-lité que l’échauffement de l’échantillon entraîne la molécule dans un état excité est donc pratiquement nulle. Le calcul de ces probabilités de changement d’état peut être obtenu grâce à la loi de Boltzmann.
Le temps entre l’excitation d’un fluorophore et l’émission d’un photon sera égal à la somme de trois temps - passage de S0 à S1, de l’ordre de 10-12 s
- temps de résidence dans les états excités, de l’ordre de 10-9 s
- retour à l’état fondamental et émission d’un photon, de l’ordre de 10-12 s
Le processus complet d’émission d’un photon sera donc de l’ordre de la nanoseconde pour les fluoro-phores usuels. Après excitation, chaque photon aura
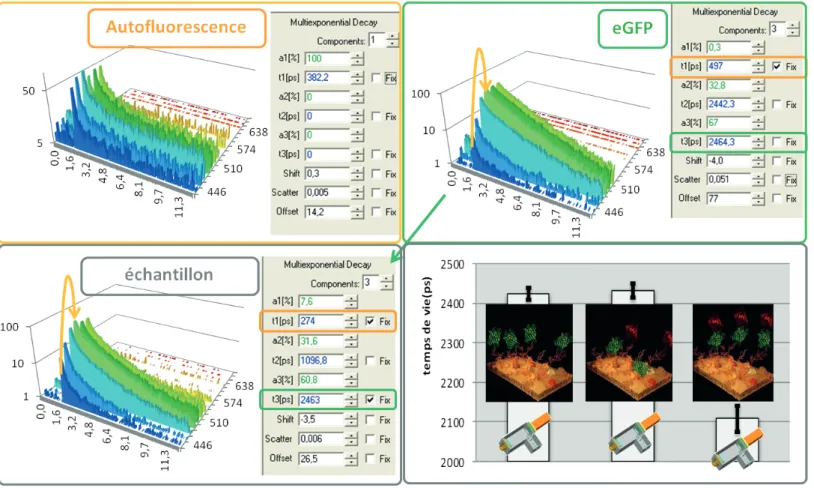
un temps d’émission qui va suivre une loi de proba-bilité de type exponentielle. Après observation d’un grand nombre de temps d’émission de photons, on obtient une courbe de déclin de fluorescence corres-pondant à cette loi de probabilité d’émission. La pente de cette courbe est alors inversement proportionnelle à une autre propriété caractéristique d’un fluoro-phore : le temps de vie de fluorescence. Cette proprié-té peut être considérée comme indépendante de la concentration au regard des niveaux d’expression de fluorophores raisonnablement utilisables en cellule. Elle sera en revanche influencée par l’environnement local et la température du fluorophore ainsi que par les modes de désexcitation dont il dispose.
Le temps de vie de fluorescence pourra être cal-culé en ajustant la courbe de déclin par la formule sui-vante i t n i i e a of t I /τ 1 . ) ( − =
∑
+ =Avec : I l’intensité ; of le niveau de bruit t le temps ; n le nombre d’espèces exponentielles ; a la proportion de l’espèce et τ le temps de vie de l’espèce.
Chacune de ces propriétés est caractéristique du fluorophore étudié. Une molécule fluorescente pourra donc être identifiée en observant son spectre d’absorption ou d’émission. La cinétique d’émission des photons, dont le temps de vie de fluorescence est extrait, est une autre méthode d’identification de ces molécules.
Spectre d’absorption (pointillé) et d’émission (trait plein) de la CFP.
Représentation semi-logarithmique d’une courbe typique de déclin de fluorescence pour une espèce mono-expo-nentielle. Le temps de vie de fluorescence (τ) peut être
fLim/sLim
Mon travail de thèse a été réalisé au sein de l’équipe d’imagerie cellulaire fonctionnelle de l’Institut de Biologie de Lille (FRE 2963, Lille), sous la direction des docteurs Laurent Heliot et Ber-nard Vandenbunder. Mon objectif était de développer et caractériser un système d’acquisition couplant mesures de temps de vie de fluorescence et de spectre d’émission, adapté à l’étude des interactions moléculaires dans la cellule vivante. Ce travail a été réalisé en étroite collaboration avec le docteur Jacques Barbillat (Lasir, UMR 8516, Lille) pour le développement instrumental. Nous avons ensuite appliqué ces nouvelles stratégies à certaines problématiques du docteur Marc Landry (LPRNM, INSERM E358, Bordeaux).
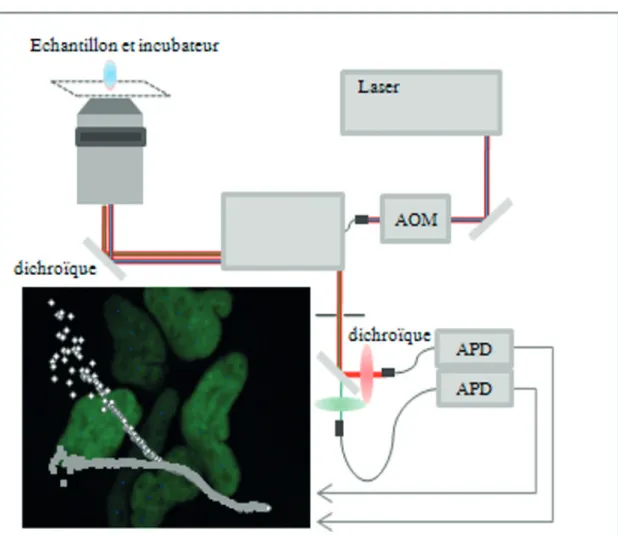
• Description générale du système d’acquisition
Le système que nous avons développé (Spriet et al., 2006) s’articule autour du micros-cope confocal Leica SP2 équipé d’une source pulsée Ti :Sa (MIRA 900-M). Nous avons choisi de travailler avec le système le plus précis pour réaliser des mesures de temps de vie de quelques nanosecondes: le TCSPC (SPC730, Becker et Hickl).
Le scanner de microscope SP2 présentant une sortie descannée, nous avons choisi d’y adapter directement le spectrographe que nous avons développé, le SPRC160.
En évitant l’utilisation d’une fibre optique, nous limitons au maximum les pertes de fluo-rescence le long du trajet optique, mais aussi une potentielle dégradation de la réponse
mentale. Nous avons également développé, en collaboration avec Dave Trinel, un logiciel per-mettant d’utiliser des routines d’acquisition allant de l’image complète de SLiM à une imagerie biphotonique associée à des mesures de SLiM localisées et répétées au cours du temps (figure 7).
L’angle solide du faisceau lumineux émergeant du scanner (α~3°) étant différent de l’angle d’entrée du spectrographe (α~16°), nous avons utilisé une optique de couplage entre le « pin-hole » de sortie du scanner et l’entrée du spectrographe.
Concrètement, la lumière en sortie du pinhole est colimatée par un doublet achromatique (DBL13944/100, JML Optical Industries, Inc) puis projetée par un second doublet (DBL14076/100, JML Optical Industries, Inc) sur le trou d’entrée du spectrographe. De cette façon, le faisceau entrant couvre parfaitement le réseau. Les photons entrants pourront alors être dispersés cor-rectement sur un détecteur multianodes (R5900U-L16, Hamamatsu). Les deux doublets achro-matiques ont été montés sur un système de translation XYZ maison conçu avec le LASIR. Cet ensemble permet de focaliser précisément le faisceau de sortie du microscope confocal sur la fente d’entrée du SPRC160 et assure ainsi la portabilité du système entre les microscopes Leica SP2 et SP5, ces derniers présentant un réglage au niveau de la position apparente du pinhole et de l’angle de sortie du scanner.
Le spectrographe par lui-même est composé d’un réseau holographique concave (Ref. 53300100, Horiba group, Jobin & Yvon SA) et d’un miroir pour réduire l’encombrement spatial du système. De par ce choix de réseau, aucun autre élément optique n’est nécessaire sur le trajet (par exemple une optique de focalisation entre le réseau et le détecteur). Le système que nous avons développé est alors très compact (figure 7) et réduit au maximum les pertes de photons.
Figure 7: le SPRC160, représentation schématique (A) et intégration sur un microscope confocal Leica SP2 avec chaîne d’excitation bi-photonique (B)
Focus sur Le FRET (Förster Resonance Energy Transfer)
Différentes techniques existent pour étudier les interactions moléculaires, aussi bien in vitro qu’in vivo, par des techniques physico-chimiques ou microscopiques. L’émergence des protéines GFP, popularisées dans les années 1990 par Roger Tsien (Tsien, 1998, 2005), a grandement participé à l’essor de ces dernières, en par-ticulier par mesure de FRET (Förster Resonance Energy Transfer) entre deux fluorophores.
Chaque fluorophore est caractérisé par un spectre d’excitation et par un spectre d’émission. S’il existe un recouvrement entre le spectre d’émission d’un fluorophore appelé donneur et celui d’excitation d’un autre appelé accepteur, il se produit un transfert d’énergie du donneur vers l’accepteur ou FRET
L’efficacité de ce transfert est définie par l’équation suivante :
Avec E l’efficacité de FRET ; F l’intensité et τ le temps de vie de la fluorescence du donneur respectivement en présence (DA) ou absence (D) de l’accepteur ; r la distance entre les deux fluorochromes ; R0 la distance de demi-transfert qui est caractéristique d’un couple de fluorophores donné et dont l’expression est:
Qd : le rendement quantique du donneur ; n : l’indice de réfraction du milieu (considéré comme étant de 1.4 pour les protéines) ; Nav : le nombre d’Avogadro (Nav = 6.02 x 1023 mol-1) ; J : la fonction intégrale expri-mant le taux de recouvrement entre le spectre d’émission du donneur et le spectre d’excitation de l’accepteur et K : le facteur d’orientation.
Dans le cas de deux molécules ayant une liberté de mouvement équivalente dans toutes les directions, le facteur d’orientation est défini par l’équation suivante :
qT : l’angle entre le moment de transition d’émission du donneur et le moment de transition d’absorption
de l’accepteur ; qD et qA : les angles entre les lignes de connections donneur/accepteur et les moments de tran-sition respectivement d’émission du donneur et d’absorption de l’accepteur.
Un événement de FRET va se traduire par des modifications des caractéristiques de la fluorescence émise par les différents fluorophores présents:
• Les propriétés spectrales du mélange de fluorophores seront altérées par la diminution de l’émission du donneur et l’augmentation de celle de l’accepteur.
• Ce transfert constitue également un mode de désexcitation supplémentaire pour le donneur. Cela implique l’apparition une diminution globale du temps de vie moyen.
Il en découle deux grandes catégories de techniques de mesure de FRET : les premières, basées sur des mesures d’intensité demandent des temps d’acquisition assez courts (quelques dizaines de millisecondes) et sont principalement qualitatives, alors que les mesures de temps de vie de fluorescence, bien que plus précises, nécessiteront des temps d’acquisition beaucoup plus longs (quelques secondes à quelques minutes).
Facteur entrant en jeu dans le calcul de l'efficacité de FRET. Gauche : spectre d’excitation et d’émission d’un couple de FRET. La zone marquée J représente le taux de recouvrement de l’équation 2. Droite : représentation schématique des angles de l’équation 3. ; Le bleu représente le donneur et le jaune,
• Caractérisation du système
Tout d’abord, le système de mesure spectrale intégré au microscope Leica SP2 nous a per-mis de confirmer la largeur et la position des gammes spectrales correspondant aux différentes anodes du SPRC160. Nous avons ensuite confirmé la reproductibilité de nos mesures spectrales sur des solutions d’acide picrique, les mesures étant effectuées à plusieurs jours d’intervalle (figure 8).
Nous avons également tiré parti d’un système de mesures de temps de vie par TCSPC précédemment implé-menté et caractérisé au laboratoire (Waharte et al., 2006). Nous avons réa-lisé des mesures sur différents échan-tillons de référence avec nos deux systèmes. L’ensemble des résultats est résumé figure 9.
Nous avons effectué des mesures sur la rhodamineB en solution aqueuse afin de déterminer la capacité du sys-tème à résoudre des déclins mono-ex-ponentiels en solution.
Nous avons ensuite réalisé les mêmes mesures sur des cellules HeLa exprimant de la CeruleanFP ou de la CFP, ces deux protéines étant celles que nous avons le plus utilisées pour effectuer nos expériences de FRET.
Enfin, nous avons testé notre système sur une cellule HeLa exprimant la protéine CFP liée à la YFP et constituant un témoin positif de FRET. Cette procédure de caractérisation m’a permis de démontrer la capacité du SPRC160 à réaliser des mesures quantitatives de spectre et de temps de vie de fluorescence dans le contexte de l’étude des interactions moléculaires dans la cellule vivante.
Figure 8: Reproductibilité des mesures de spectre sur une solution d’acide picrique
Figure 9: Mesures de temps de vie de fluorescence (ps) de différents
échantillons de référence, acquises grâce au système de TCSPC utili-sant un détecteur de type MCP (barres pleines) ou le SPRC160 (barres