Rôle de la protéase mastocytaire de type 4 dans la conversion de la big-endothéline-1 en endothéline-1(1-31) chez la souris
Texte intégral
Figure
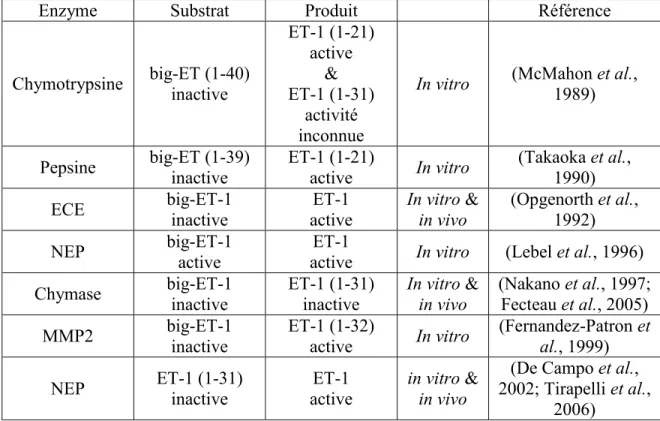


![Tableau 2 : Substrats connus de la chymase et leurs références [tableau copié intégralement à partir de (Pejler et al., 2007)]](https://thumb-eu.123doks.com/thumbv2/123doknet/5397727.125640/31.918.163.733.230.1005/tableau-substrats-connus-chymase-références-tableau-intégralement-pejler.webp)
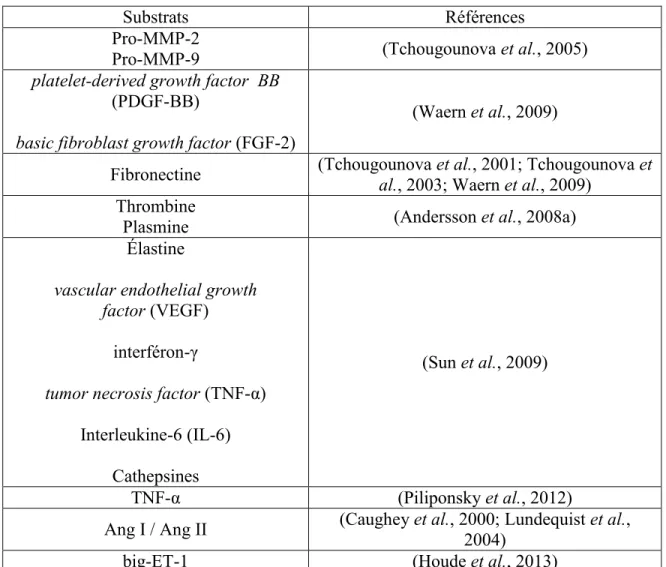
![Figure 5 : Schématisation des voies de synthèse de l’endothéline-1 [modifiée d’après (D'Orleans-Juste et al., 2003)]](https://thumb-eu.123doks.com/thumbv2/123doknet/5397727.125640/33.918.171.800.278.728/figure-schématisation-voies-synthèse-endothéline-modifiée-orleans-al.webp)



Outline
Documents relatifs
2) On voudrait calculer les distances qui séparent Barnabé (noté B sur la carte) et Charlotte (notée C sur la carte) de la côte (d) la plage de Dinard.. Activité 2.. 1) a)
Une origine (point d’application) Un point où l’on souhaite mesurer la vitesse Sa direction : La droite tangente à la trajectoire au point M. Son sens : Celui
[r]
Donner la mesure de l’angle marqué en justifiant
[r]
Exemple : on peut placer les souris face contre la table, celui qui pioche une souris particulière (souris blanche, zèbre, panthère, marquée d’un signe particulier…) a le droit
RuNGE das Verdienst gelassen werden, diese yon ihm unab- hangig aufgefundene Methode in ausserordentlich durchsichtiger und ele- ganter Weise begr~indet zu haben;
La prévalence du VIH est très élevée dans notre étude (80,7 %, car les patients hospitalisés sont plus souvent atteints de pathologies sévères, en particulier infection par le VIH
