The study of the contribution of repetition-dependent mechanisms to consolidation processes in the primary motor cortex and the electrophysiological manifestations of monetary reward processing over cortical motor areas
Texte intégral
Figure
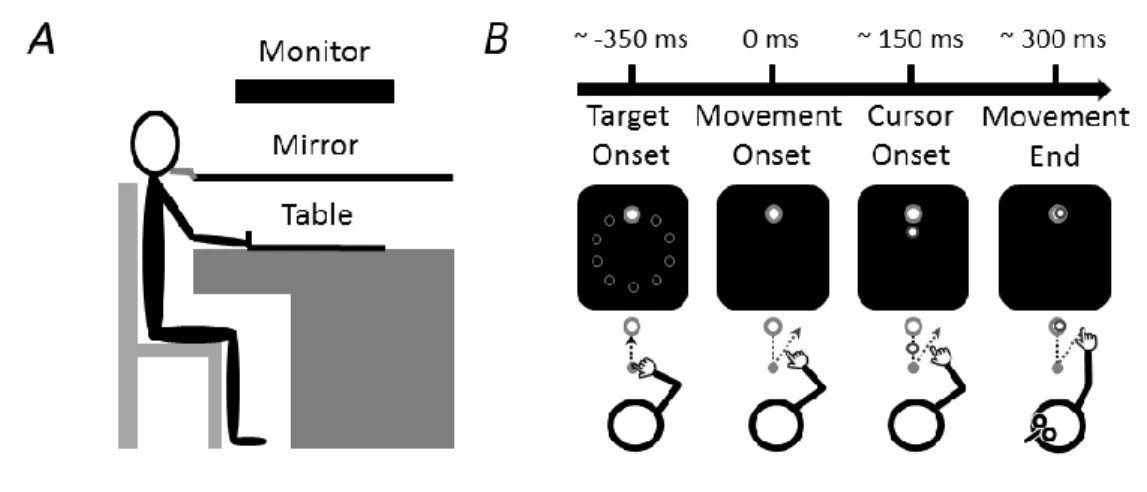


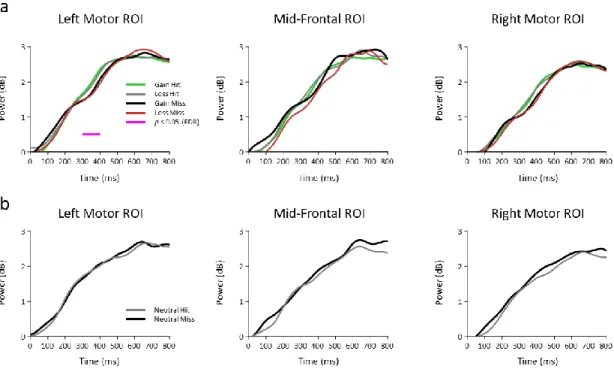

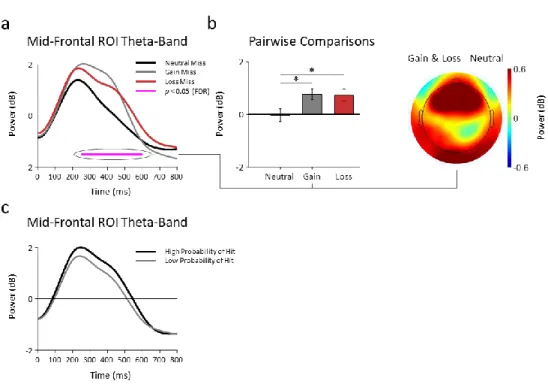
Documents relatifs
Using targeted chemogenetic in vivo approaches in the adult visual cortex from male and female mice, we found that transient inhibition of PV interneurons,
Second, inhibition of the motor system is expressed by an increase in excitability of GABA B ergic intracortical inhibitory circuits as early as 134 msec after the stop signal..
Note that this is made without the use of any type of fuel, because the energy, which moves the Gravitational Motor comes from Earth’s gravitational field, i.e., the
Il s’agit pour les étudiant.e.s de master (désormais “E-M2”), dans un module consacré à l’intégration du numérique dans l’enseignement/ apprentissage des
C ette édition de la Revue des sciences de l’éducation de McGill est remplie d’histoires, des histoires riches et variées, des histoires de différentes cultures, contextes
Malgré toutes ces épreuves, où elle a dû tomber pour se relever, elle s’accrochait à la Vie, Vie qui ne la pas épargner : ce qui était facile pour les uns, lui paraissait dur
Elle adapte cela en fonction des besoins des élèves, car le fait de réaliser ce rituel avec toute la classe permet de compter un nombre plus grand et cela est plus
This study presents an application and extension of Raupach’s model to surface shear- stress measurements in live plant canopies of a single species (Lolium perenne) of
