Extent of intra-isolate genetic polymorphism in glomus etunicatum using a molecular genetic approach
Texte intégral
Figure

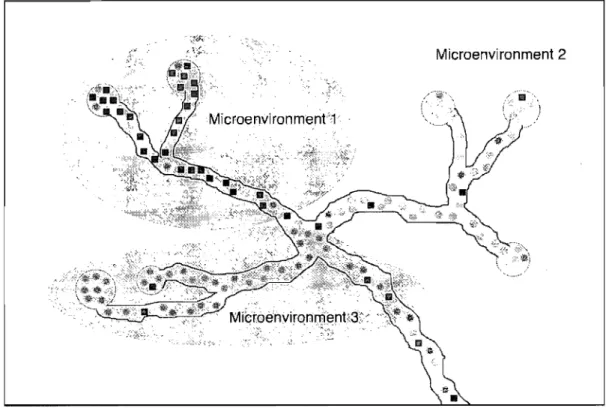
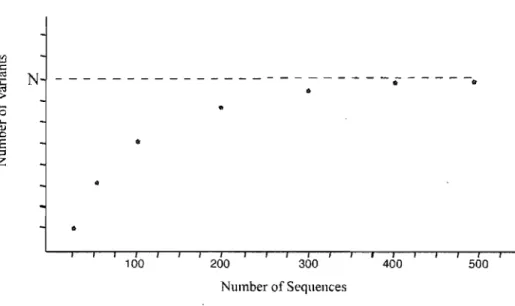
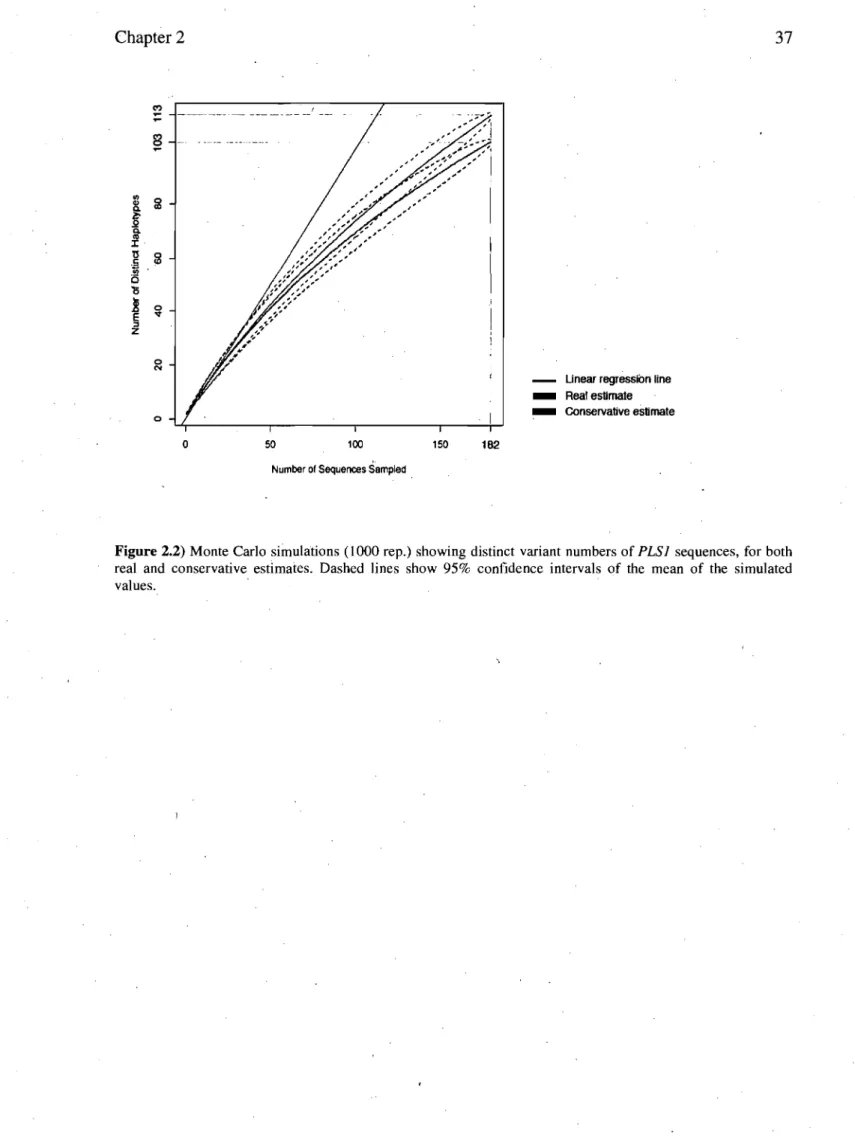
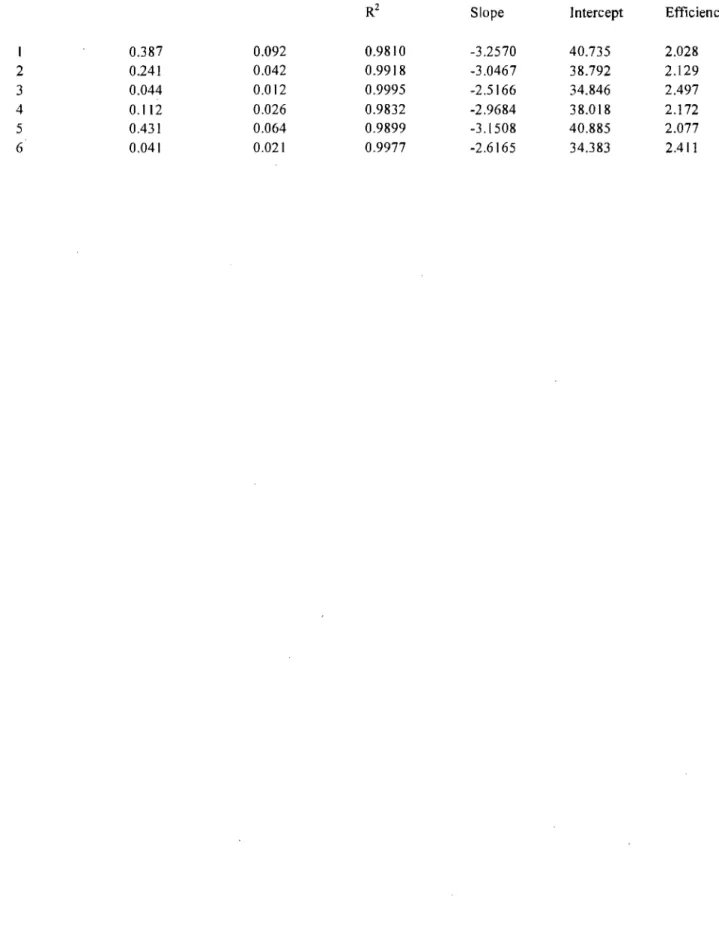
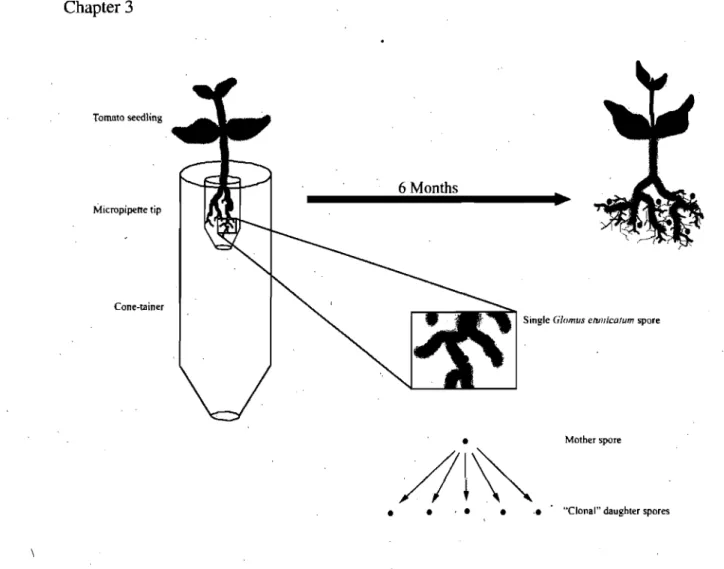



Documents relatifs
Diederik van Tuinen, Marie-Lara Bouffaud, Eric Bernaud, Annie Colombet, Daniel Wipf, Dirk Redecker.. To cite
To study the potential of the mycorrhizal fungi Glomus mosseae and Glomus intraradices to stimulate the growth of micropropagated olive plants and to compare their capacity
Several studies investigating the role of AMF in protection against salt stress have demonstrated that the symbiosis often results in increased nutrient uptake, accumulation
Notre seconde hypothèse étant : “un score élevé au questionnaire de catastrophisme de la douleur est lié à une stratégie de mouvement thoraco-pelvienne plus en-phase
ries infantiles modernes plagiant plus ou moins la doctrine génitale mater-nelle du yin et du yang, la part zoociométrique est femelle comme la zoociotonique
Miss rates of nativelikeness criteria for a single task depending on the size of the native control sample (8, 32, or 128 speakers), the distribution of the task scores in the
Amino acids and phenolic compound synthesis combined to P nutrition are potential mechanisms used by these biological control agents for plant protection. These results
