UNIVERSITÉ D QUÉBEC À MO 'TR ··AL
TYPOLOGIE STRUCTURALE DES PEUPLEME TS MÉL 'GÉS
E FORÊT BORÉALE DE L'OUEST DU Q ÉBE
MÉMOIRE
PRÉSE TÉ
OMME EXIGE CE PARTIELLE
DE LA MAÎTRISE E BIOLOGIE
PAR
SYLVAI LAR UCH
UNIVERSITÉ DU QUÉBEC À MONTRÉAL Service des bibliothèques
Avertissement
La diffusion de ce mémoire se fait dans le respect des droits de son auteur, qui a signé le formulaire Autorisation de reproduire et de diffuser un travail de recherche de cycles supérieurs (SDU-522 - Rév.01-2006). Cette autorisation stipule que «conformément
à
l'article 11 du Règlement no 8 des études de cycles supérieurs, [l'auteur] concèdeà
l'Université du Québecà
Montréal une licence non exclusive d'utilisation et de publication de la totalité ou d'une partie importante de [son] travail de recherche pour des fins pédagogiques et non commerciales. Plus précisément, [l'auteur] autorise l'Université du Québec à Montréal à reproduire, diffuser, prêter, distribuer ou vendre des copies de [son] travail de rechercheà
des fins non commerciales sur quelque support que ce soit, y compris l'Internet. Cette licence et cette autorisation n'entraînent pas une renonciation de [la] part [de l'auteur]à
[ses] droits moraux nià
[ses] droits de propriété intellectuelle. Sauf entente contraire, [l'auteur] conserve la liberté de diffuser et de commercialiser ou non ce travail dont [il] possède un exemplaire.»REMERCIMENTS
Mon passage à la maîtrise représente un accomplissement qui me rend extrêmement fier. e ont les gens qui nt fait ce parcours à mes côté , qui m'ont soutenu t épaulé, encouragé et in piré, qui en ont fait une expérience exceptionnelle. Avant quiconque, je remercie mes directeur Alain Leduc et Brian Harvey pour leur disponibilité, leurs judicieux con eils, leur enthou iasme et leur générosité. Un merci tout particulier à Alain pour son support technique et ses innombrables id· e lumineuse . Un merci particulier à Brian qui, malgré les 620 kilomètres qui nous séparaient, s'est toujours montré intéressé à mes travaux. Merci pour la collaboration essentielle clïan Moss, an quoi ce projet n'aurait pu exi ter. Pour son encadrement technique exemplaire lor des activités de terrain, et pour m'avoir fait confiance en 2009 à titre cl' Aide de terrain, là où tout à commencé, je remercie Danielle Charron. Aus i, un gros merci à mon aide de teiTain, Lucile Muller. Merci à
0
valdo Valéria, à Frédérik Doyon el à Yan Boucher pour leur collaboration à titre de membre de mon conlité de recherche. Un gro merci à tous les profe sionJ1 ls de recherche du CEF, en particulier Tech, Mélanie et Luc. Et pour le support tatistique, et sa capacité hors du commun à transmettre sa passion pour l'é<X>Iogie et la statistique, je remercie Pierre Legendre. San votre pré ence, mon projet n'aurait sans doute pas existé sous cette fo1me.Merci à toute l'équipe du labo de la Chaire industrielle CRSNG - UQAT- UQAM en aménagement forestier durable. D'abord, merci à D minic (et à Sancl1ine) pour avoir littéralement transfon11é l'ambiance du labo en la rendant davantage propice à un travail efficace! Merci à Popo qui, avec ses connais ances inouïes sur la structure des forêt , m-a fait comprendre ce que c'était concrètement. Merci à Pipi et à Annie-Claude pour leur support technique avec R et ArcGIS. Vous êtes trop fortes les filles' Merci à Cécile, J hann, Pien·e et Amar pour avoir enrichi l'ambiance du labo. Au i, merci à Jeanne pour son upport moral lor de derniers mille (les plus pénibles') de ma maîtrise. Finalement, merci à lous ceux que je n'ai pas nommés ci-haut, mais qui m'ont accompagné lors des pauses diner et café (et concs), au B nelux ou dans quelconques soirées, souvent inoubliable . Ici, je pense particulièrement à Simon Paradis et à Marc-Olivier Audet, les deux eule person.nes de mon entourage à qui j'ai pu parler quotidiennement et continuellement de hockey. Sans votre présence dan ma vie, l'envergure de mon projet aurait été nettement plu limitée.
IV
Le plu gro des mercis va à Laurie pour son support constant hors du bureau. Sans toi, je ne serai sans doute jamais devenu biol gi te et je ne serais pas en train d remercier tout c beau monde. Merci pour les belles années qu'on a passées ensemble dans la jungle montréalaise, j'en garderai de ouvenirs inoubliables. Évidenunent, je n'aurais pu y parvenir ans le support de ma famille tout au long de mes études. Merci à me parent , Nicole et Jean-Rock, qui m'ont supporté dans la poursuite de mes études en écologie, un domaine plutôt indigeste pour de parents typiques' Merci à ma sœur, Marie-Line, à mon beau-frère Bruno et à mon filleul Ludovic. Votre présence au quotidien me manquera énormément.
Par ailleurs, ce travail de recherche a été upp011é financièrement par la subvention RDP J 395368 (Eastern boreal mixedwoods: Multiscale analysis of stand structure, dynamics and si/vieu/ture) du programme Recherche et développement coopérative lu
RSNG.
Finalement, je tien à mentiormer que j'ai beaucoup apprécié !"école de pen ée sur !"aménagement écosy témique véhiculée par les chercheurs de la Chaire en aménagement fore ·tier durable. Celle-ci a radicalement changé ma perception de l'aménagement forestier, pour le m.ieux, en contra tant ma vision préconçue, elle-même forgée par ma fom1ation technique en pâtes et papiers et mes quatre étés de reboisement intensif d'épinette noire. r e père être en mesure de transmettre efficacement et rigoureusement ces com1ais ances au lac, une région qui, je crois, en a vivement besoin.
TABLE DES MATIÈRES
LISTE DES FIG RE VIl
LISTE DES TABLE UX... LX RÉSUMÉ GÉNÉRAL ... . Xl INTRODUCTION GÉ1 ÉRALE ... ..
Pro b 1 éma tique ... .
État des con.nai sances ... .. 3
3 Particularités de la forêt boréale mixte ... . Évolution de la structure en absence de perturbations... 5
Altération de la structure par les perturbations secondaires ... _... 9
Analyse d la structure... 11
CHAPJTRE l TYPOLOGIE STRUCTURALE DES PEUPLEME TS MÉLA GÉS DE LA FORÊT BORÉALE DE L'OUEST DU QUÉBEC.. _ ... ... 17
1.1 1.2 1.3 1.4 1.5 1.6 Résumé ... .. Introduction ... .. Aire d'étude ... ··-·-···-···-··· Donnée et méthodes ... .. 1.4.1 Paramètres typologiques ... . 17 19 22 24 24 1.4.2 Confrontation des typologies c-means et k-means ... 27
1.4.3 Triangle des structures ... .
28
1.4.4 lnfluence de la campo ition ur la structure... 29
1.4.5 Détem1ination du potentiel de régénération ylvicole ... 29
1.4.6 ù1fluence des perturbations secondaires sur la structure... 30
Résultats._ ... _ ... _ ... . 1.51 1.5.2 1.5.3 Paramètre typologiques ... . Confrontation des typologies c-means et k-means ... .. Triangle des structures ... .. 31 31 37 38 l .5.4 Influence de la composition sur la lructure ... 38
1.5.5 Strate de gaule et le potentiel de régénération sylvicole... 39
1.5.6 Influence des perturbations secondaires ur la tructure... 43
VI 1.7 Conclu ion ... . APPENDICE A ... . APPENDICE 8 ... . APPENDICE 51 53 55 59 APPENDICE D... 61 APPENDICE E ... ... 63 APPENDICE F ... ... 65 APPENDICE G... 67
1.8 Bibliographie ... . 68
CHAPITRE II ROBUSTESSE D'UNE TYPOLOGIE STRUCTURALE: TNVESTIGATION SUR L'HÉTÉROGÉ ÉITÉ STRUCTURALE DE PEUPLEMENTS EN FORÊT BORÉALE MIXTE ... .
79
79
81 82 84 2.1 2.2 2.3 2.4 2.5 2.6 2.7 Résumé ... . Introduction ... . Aire d'étude ... . Donn ·es et méthodes ... ." ... .2.4.1 Typologie structurale... 84
2.4.2 Description de placelles d'un hectare étudiés... 84
2.4.3 Évaluation de la robustesse typologiqu Résultats ... . 85 86 2.5.1 Robustesse typologique à l'échelle de 400m2... 86
2.5.2 Robustesse typologique des échelles supérieures... 86
2.5.3 Sensibilité typologique à la détection de tiges de fort calibre... 88
Discussion ... . Conclu ion ... . 91
94
APPENDJ E 1-I... 95 2.8 Bibliographie ... .96
0 CLUSlO GÉ ÉRALE... ... .. ... ... 99 BIBLIOGRAPHIE... 103LISTE DES FIGURES
Figure
1.1 Répartition des plac !tes-échantillons temporair s utilisées dans le ten·itoire d'étude (Sa : n = 396 et 6a: n = 65) ... ... . 1.2 Schéma du triangle des structures, représentation équilatérale (Adapté de
Herbert et Allegrini, 2000). Les ela se de diamètre (DHP) sont PB=
20-Page
23
25cm, MB=30-45cm et GB=50 cm et plus. ... 28 1.3 Silhouette plot des 461 placette -échantillon temporaires... 32 1.4 Repré entation des types -me ms sous leur fonne cumulée relative. A) la
surface terrière moyenne cumulée absolue; B) la den si té moyenne cumulée
absolue... 33 1.5 Représentation des types k-rneans sous leur fom1e cumulée relative. A) la
urface terrière moyenne cumulée absolue; B) la densité moyenne cumulée
ab olue... 34 1.6 Repré entation des types c-means sous leur forme non cumulée absolue. A)
surface tenière moyenne ab olue: 8) densité moyenne absolue ... . 1.7 Boîte à moustache représentant les p1incipaux paramètres typologique en
fonction du type c-means, en occurrence la surface tetTière (A), la densité
35
(B), le volume ( ) et 1 DHP moyen (D) ... 36 1.8 Triangle des structures représentant la distribution moyerme des types (Tl à
T8) en fonction de la dimension des arbres... 40 1.9 Déviances des quatre p1incipales trajectoire successionnelles en forêt
boréale mixte... 41 1.10 Pourcentage cumulé exprimant la composition moyenne des types
tructuralement ordonnés en tem1es de A) surface terrière et de B) densité... 42 1.11 Pourcentage de chaque groupe de gaulis : résineux tolérants, feuillu
intolérants, arbustes lign ux et absence d régénération pour chaque type
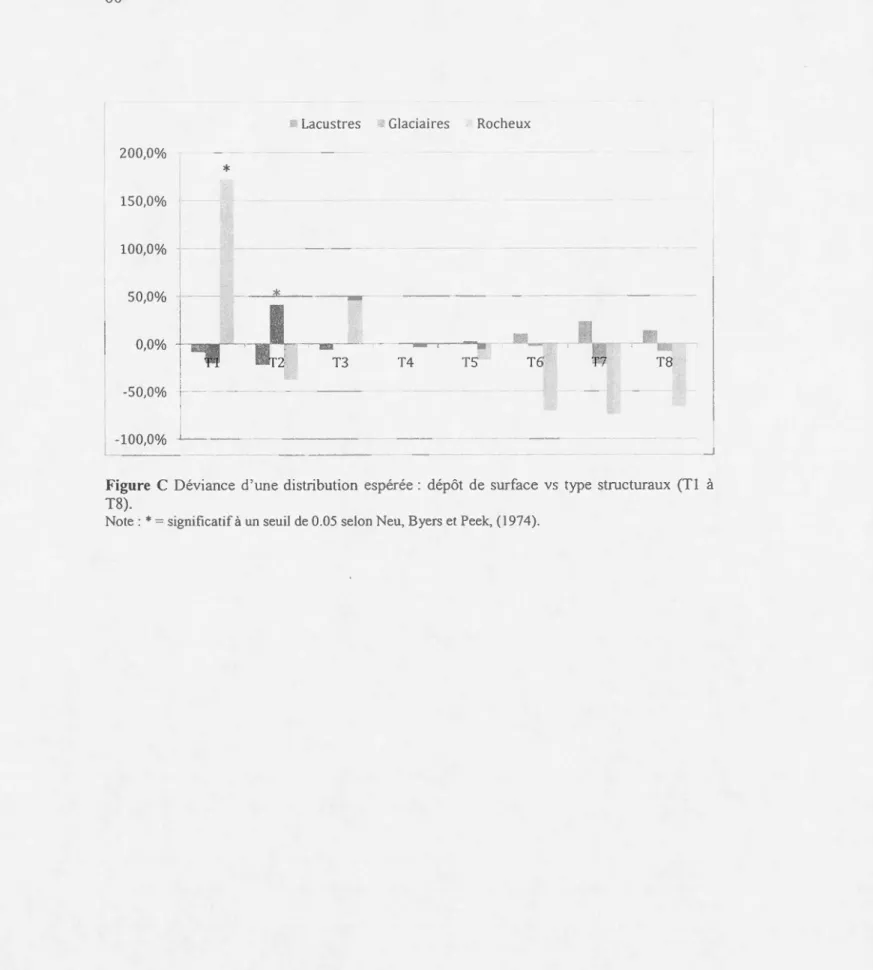
tructural, selon leur abondance en terme de ur face tetTière. ... ... 43 1.12 Déviance d'une di tribution espérée: composition v structure... 44
Vlll
2.1 Carte du lac Duparquet (Abitibi, Québec, Canada) montrant la locali ation des zones incendiée d'intérêt, avec l'am1ée du plus récent tèu. Le points noirs représentent les bomes qui délimitent chaque hectare.
2.2 Plan de découpage de quatre hectare pem1anent . Les superficie de 400 m2, 600 m2 et 800 m2 sont représentées pour chacun d'entre eux (A: 1916; B: 1870; C: 1847; D: 1823). En rouge: les po1iions non utili ées suite au
83
découpage; en ve1t: les p01iion appartenant à un autre polygone forestier... 89 2.3 Boite à moustache représentant la distribution de la taille de la plus grosse
LISTE DES TABLEAUX
Tableau Page
1.1 Résultats de mesure de validation inteme obtenus par les algoritlunes de
groupement c-means et k-means ... 31
1.2 Distribution des objets résultant d'une typologie à 8 types avec les algorithmes de groupement k-mean · et c-means. En rouge gras sont mis en
évidence les placettes appar1enant à un type commun. . .. ... ... ... .. .. ... .. .. ... .. 3 7
2.1 Distribution des type tructuraux au sein des quatre hectare pem1anents à l'étude, à l'échelle du 400 m2, de 600 m2 et de 800 m2 Pour chacu11e de échelles, les résultats sont présentés sou fonne de valeur absolue (Abs) et
de pourcentage cumulé(%)... ... ... 87
2.2 Pourcentage d'erreur différence de pourcentage entre deux analyses
x
RÉSUMÉ
La structure interne des peuplements m'langés boréaux est un attribut clef, entre autr s pour le maintien de la biodiversité et de la productivité forestière. Dans le contexte où !"on vise à diversifier de pratiques sylvicoles et à pem1ettre une prise de décision alim nléc par une reconnaissance plus fine de la structure et la dynamique des peuplements mélangés, les outil~ actuellement disponibles aux aménagistes s'avèrent largement inadéquats. Afin de combler cette lacune, notre p1incipal objectif visait à élaborer une typologie structurale fine des peuplements mélangés de l'Ouest du Québec en se basant sur la distribution de la densité et de la surface teJTière des arbr s commerciaux. L'analyse a été effe tuée sur 461 placette -échantillons temporaires provenant du Ministère des ressources naturelles situées dans les régions écologiques Sa et 6a. Le types structuraux de référence ont ensuite été ordonnés en suivant leur succession structurale théorique afin d'évaluer a posteriori l'influence de la compo ition et des perturbations secondaires sur la structure fore tière. La robuste se de la typologie, à savoir son pouvoir d'extrapolation spatiale à une unité d'intervention (i.e. le peuplement fore tier cartographique), a été testée sur quatre hectares de forêt mélangée qui contiennent une variabilité structurale se rapprochant de celle observée à l'échelle de l'unité d'intervention. Dan chacun d'entre eux, tous les arbres ont été cartographiés et mesuré , ce qui a permi de recréer des sous-placettes de taille variable. Le résultats suggèrent qu'il est possible de résumer le gradient structural du territoire d'étude en huit types structuraux à l'échelle de la placette-échantillon ( 400 m2) et représentatifs des parcelle forestières plus grande . Des relations entre la structure des peuplements et les effet de la compo ilion el de la tordeuse des bourgeons de l'épinette ont pu être identifiées. Ceci suggère un cadre duquel il sera possible de prescrire des cibles d'aménagement adaptées à la tructure interne de peuplements et ainsi mitiger le conséquences de l'aménagement sur la biodiversité et la productivité des forêts.
MOTS-CLÉS: Forêt boréale mixte; Structure forestière; Composition forestière: Tordeuse de bourgeons de l'épinette; Aménagement forestier écosy témique.
INTRODUCTION GÉ ÉRALE
Problématique
La forêt boréale occupe près du tiers de la superficie forestière mondiale (CCMF 20 12). Ce biome se distingue par deux traits fondamentaux. Avant tout, son climat froid représente une contrainte à l'établissement et au maintien de espèces végétales dont le métabolisme est presque nul en hiver (Bonan et Shugart, 1989). La forêt boréale est aussi un biome fortement perturbé. Ses paysages sont dynamisés par la récutTence de incendie fore tiers (Rowe et Scotter, 1973 ; Heinselman, 1981 ; Johnson, 1992) el de épidémie d'insectes (Blais, 1983). Au fil des génération , certaine espèces arborescentes de la forêt boréale se sont adaptées à ces pertmbation , par exemple en développant des mécani mes leur pem1ettant de coloniser les brûlis ou en profitant d'autres traits qui favorisent leur survie et maintien en périodes d'absence de feux (Bergeron 2000).
En Amérique du Nord, dans les années 1970, une décetmie as ociée au développement de la mécanisation des opérations de la récolte, la forêt boréal est devenue économiquement attrayante pour l'industrie, on a alors commencé à y fa ire un aménagement extensif piani fié (Coulombe et al., 2004; Boucher el al., 2009). La recherche scientifique, qui a connu à cet époque un véritable essor, s'est orientée ver l'étud d feux de forêts, principale menace à la ressource (Heinselman, 1973 ; Rowe et Scotter, 1973 ; Van Wagner, 1978). et intérêt pour les feux a ouvert la voie à des recherches plus fondamentales ur la dynamique forestière qui avaient pour objectif de documenter les interaction entre le espèces, les éco ystèmes, les paysages et les régimes de perturbation (Dix et Swan, 1971 ; Hein elman,
1981 ; Pickett et White, 1985).
De 1986 jusqu'à tout récenm1ent, la Loi sur les .forêt· du Québec planifiait une nom1alisation de la structure d'âge des paysages fore tier dans l'optique d'assurer un rendement soutenu de la matière ligneuse (Coulombe el al., 2004). Or, les recherches menées au cours des dernière années ont remis en question le uppositi n qui avaient jusqu·alor justifié un aménagement extensif de la for~t boréale de l'Est de l'Amérique du ord (Gauthier, Leduc et Bergeron, 1996 ; Bergeron el al .. 1999 ; Bergeron el al., 2006). Par exemple, on reconnaît maintenant que la forêt de 1· e 1 du a nada pré entait historiquement une moyet1J1e d'âge nettement plus élevée que celle de la révolution fore tière qui varie
2
généralement entre 60 et lOO ans. Les intervalles entre les feux se sont clone avérés plus longs en moyenne que ceux qui avaient été a sumés à pa11ir le recherches menée clans le anacla continental (Gauthier et al., 1996 ; Bergeron et al., 2006 ; Bouchard, Kneeshaw et Bergeron, 2008). Tl en résulte aujourd'hui que les pay age forestiers aménagés depuis plusieurs décennies apparaissent signiticativement plus jeunes que ceux des territoires non aménagés, soum1s à un régime de perturbations naturel clepui les 8000 clemière année (Cyr et al., 2009)
L'altération des paysages forestiers par l'aménagement soulève des enjeux quant à la résilience des forêts aménagées, quant à de la durabilité de la re source fore tièr et quant au maintien de la diversité biologique (CCMF, 2003; Coulombe el al., 2004; Gauthier et al., 2008). Par exemple, en Fennoscandinavie, l'aménagement intensif de la forêt boréale qui
·opère depuis plus d·un siècle a profondément changé la dynamique et le caractère naturel de écosystèmes fore tiers, ce qui a entre autre caus' une diminution impo11ante de la variabilité stru tw·ale de peuplements (Uihde, Loile et orokorpi, 1999) et de la cliver ité biologique (Berg et al., 1994 ; Ostlund, Zackrisson et Axelsson, 1997 ; Ra si et al., 200 1). Dan la forêt boréale de l' st de l'Amérique du Nord, l'homogénéi ation de pay age forestiers par 1 'aménagement t la raréfaction des viei Il s forêts sont identi Gées comme des menace importantes au maintien de la biodiversité (Gauthier et al., 2008; Cyr et al., 2009), un élément clef associé à la ré ilience des écosy tème (Walker, Kinzig et Langridge, 1999).
Avec l'adoption d la Loi sur l'aménagement durable du territoire forestier, le gouvemement du Québec a changé ses paradigmes en préconisant la mise en œuvre de l'aménagement fore ti er écosystémique (Québec, 20 13). Cette stratégie a comme objectif de concilier le maintien d'écosystèmes sains et ré ilients de même que la durabilité de la ressource ligneuse el de l'industrie forestière. Pour ce faire, l'aménagement forestier écosystémique mise sur une réduction des éca11s ntre le paysage naturel et aménagés (Grumbine, 1994; hristen en et al., 1996; Gauthier et al., 2008). Le concept, riginalement développé pour 1 'aménagement des forêts de la Côte-Ouest américaine, a été adapté pour répondre aux besoin de divers environnements. La mi e en œuvre de l'aménagement écosystémique e t donc intimement liée à une connai sance et à une compréh nsion plu approfondie de écosystèmes naturels et aménagés (Christen en et al .. 1996).
À
cet égard, un aspect important dans la mise en application de l'AFÉ est l'acquisition de connaissances..., .)
fines sur la structure, la composition el la dynamique naturelle des peuplements forestier (Harvey et al., 2002).
État des connaissances
Une brève revue de littérature ur le sujet d'étude est synthétisée en quatre sections. Celles-ci déclineront succinctement ( l) les principales particularités de la forêt boréale mixte, (2) conu11cnt la tructure est susceptible d'y évoluer en absence de perturbation et (3) conu11ent les perturbation secondaires sont su ceptibles de l'altérer. Finalement, une dernière section pré entera ( 4) les principale approche de classification structurale el plus
péci fiquement l'approche privilégiée dans cette étude.
Particularités de la(orêt boréale mixte
La région d'étude, c'est-à-dire l'ensemble des régions écologiques Sa et 6a (Saucier et Robitaille, 1998), est située dans la ceinture d'argile de Québec et de J'Ontario, une large région physiographique créée par les dépôt lacustres ré ultant de l'extension des lacs preglaciaires Barlow et Ojibway (Vincent et Hardy, 1977). Au sein de cette région, le plaines pa11ie!Jement remaniée ou érodées avec des dépôts morainiques sont intercalées dans les lacs preglaciaire (Bergeron et al., 1982). Les dépôts lacustres, glaciolacustres de texture fine (argile) et grossière (sable) ainsi que les dépôt organiques sont les dépôts dominants (Bouin et Berger, 2002; 2005). L'abondance de dépôts riches à texture fines est en partie responsable de 1 'abondance du peuplier faux-tremble (Populus tremuloides [Mich.]) et de la prolifération de nombreux peuplements m'langés (Sims et Uhlig, 1996). En Abitibi spécifiquement, ajoutons les effets anthropiques de la colonisation et du défrichage à partir des années 1920 ainsi que, subséquemment, le développement forestier et minier et 1 'abandon des teJTes agricole dans la région.
L'abondance des peuplements feuillus et mélangés au em du territoire d'étude ·explique aussi par un cycle de feu plutôt cou11, comparé à celui de la Côt -Nord par exemple (Bergeron et al., 2004; Cyr, Gauthier et Bergeron, 2007; Bouchard, K.neeshaw et Bergeron, 2008), en rai on de la plus faible quantité de précipitation (Biouin et Berger, 2002; 2005). Dans la région d'étude, la pe11urbation naturelle qui module le paysage de la façon la plu significative demeure le feu (Bergeron et al., 2004). Conséquemment, la
4
structure et la compo ilion des forêt reflètent le temps écoulé depui le dernier incendie (Gauthier, De randJ ré ct Oergeron,
2
000:
Arseneault,200
1
)
de même que par les traits fonctionnel des e pèce , incluant les modes de reproduction aprè et en absence de perturbations, la tolérance à 1· ombre, le taux de croissance et la longévité relative. Outre les incendies fore tier , la structure des peuplements boréaux mixtes est également modulée par de perturbations secondaires telles que les épidémies d'in ectes, le chablis, les maladies et la sénescence naturelle des arbres. D'ailleurs, les épidémie d'insecte peuvent opérer à des échelles supérieure que celle des feux, mais celle -ci génèrent des patron de pe11urbation plus fins (Knee haw et Berg ron, 1998).u fil du temps, les espèces d'arbre de la forêt boréale ont développées diverses adaptations qui leurs pennettent de tolérer les perturbations naturelles récurrentes. Incidemment, le trait vitaux actuel de ces e pèce (voir Appendice F) nou pcm1ettent entre autre d'expliquer leur stade successionnel, leur spécialisation envers ce11aines niches 'co logique ou encore leur répartition sur 1 'ensemble du teiTitoire. Conséquemment, le essences ré meuse 1 es pl us conu11une de la forêt boréale m.ixte sont l'épinette noire (Picea mariana [(Miller) Britt.]), l'épinette blanche (Picea glauca [(Moench) Vo ]), le apin baum.ier (Abies balsamea [(L) Miller]), le pin gris (Pinu banksiana [Lambert]), le thuya de l'Est (Thuja occidenrali [L.]) t le mélèze laricin (Larix /aricina [(Du Roy) K. Koch]) tandi qu les principales e sences feuillues sont le peuplier faux-tremble, le peuplier baumier (Populu bal am((era [L.]) et le bouleau à papier (Berula papyri(era [Marshall]) (Grondin, Blouin et Racine, 1998).
En moindre parti , on retrouve d'autres espèce dan le territoire d'étude, telles que le p111 blanc (Pinus sl!·obus [L.]), le pin rouge (Pinus resinosa [Ait.]), le frêne noir (Fraxinus nigra [L.]), l'orn1e d'Amérique (Ulmus americana [L.]), le bouleau jaune (Beru/a al/eghaniensis [Britt.]) et l'érable rouge (Acer rubrum [L.]). Pour la plupart, ce espèces se retrouvent à la limite nord de leur répa11ition et occupent généralement de niches écologiques davantage pécialisées.
Diver e adaptations expliquent la présence de ces espèces dans le territoire d'étude. Par exemple, la résistanc aux feux de surfac des gr pins rouge et blanc e t attribuable à leur épai e écorce tandis que les peuplement feuillus rési tent un peu plus la propagation du feu, simplement parce qu leur feuillage est moins inflanm1able que celui des conifères. Les
5
adaptations de J'épinette noire, du pin gri , du peuplier faux-tremble et elu bouleau à papier agissent pour assurer la résilience (plutôt que la résistance) des peuplement en favori ant la régénération aprè feu. Dans le cas du sapin baumier, de l'épinette blanche et du thuya de I'E t, leur maintien est plutôt as ocié à leur tolérance à l'ombre et leur longévité, à l'exception elu sapin.
Évolution de la structure en ab ence de perturbations
En forêt boréale, le feu est la perturbation à l'origine de !"initiation d'une majorité de peuplements (Johnson, 1992; Payette, 1992). À petite échelle, la usceptibilité au feu d'un peuplement varie selon les facteurs endogènes tels que a topographie, son altitude, son type de dépôt, sa végétation, son degré d'exposition au soleil, à la présence de et selon les facteurs exogènes c mme le conditions météorologiques précédant 1 'allumage, la vites e et la direction elu vent (Dan ereau et Bergeron, 1993; 1-lély, Berg ron t Flannigan, 2000; Cyr, Gauthier et 8 rgeron, 2007). La sévérité variable de feux contribue à diversifier la structure des forêts et peuplements en préservant des îlots intacts de tailles variables et des mélanges d'arbres mo1is et vivants dans différentes proportions (Bergeron et al., 2002). La évérité d'un feu influence également les caractéristiques du sol, notamment le degré de combustion de l'horizon organique, un facteur essentiel pour expliquer les divers patrons de sucee sion (Bergeron et Dubuc, 1989; Gauthier et al., 2008), de même que la composition des peuplements (Arseneault, 2001 ).
Une foi le peuplement initié, sa structure interne sera modelée par les caractéristiques biophysiques du milieu, les taux de croissance de arbres présents, la mo11alité des arbre liés à la compétition intra-et interspécifique et au vieilli sement naturel et/ou aux évènements de pe1turbations n1.ineurs (i.e. énescence chablis, épidémies légères, etc.) ainsi que l'établissement de la régénération dans le peuplements mûrs. Réciproquement, les caractéristiques structurales d'un écosystème forestier influenceront le conditions microclimatiques (i.e. la disponibilité de lumière, l'humiclité, la température, etc.) et les conditions d'habitat qui supp011ent les communautés d'espèce animales et végétales qui y sont associées (Brais et al., 1995). Ain i, la configuration structurale d'un peuplement lui conférera une résistance et une résilience particulière qui se répercut ra ur la sélectivité et la sévérité des évènement perturbateurs et entraînera des répercus ions divergentes sur la
6
structure de ce peuplement (e.g. Cooke et Lorenzetti, 2006; Cyr, Gauthier et Bergeron, 2007) À 1 'échelle du peuplement, selon le type de dépôt dominant, on distingue différentes trajectoires successionnelles qui sont propices au développem nt de peuplements mélangés (Bergeron et Dubuc, 1989; Bra ard et Chen, 2006). Plus généralement, on illustre ce trajectoires par le concept de succession forestière, défini comme étant la somme de changement dans la structure et la composition des écosystèmes forestiers dan le temps (Oliver et Larson, 1990). En forêt boréale mixte, Brassard et Chen (2006) distinguent quatre stades développementaux d'un peuplement mélangé en se basant ur la distinction structurale induite par le régime de perturbation : 1) l'initiation du peuplement; 2) l'exclusion des Liges: 3) la transition de la canopée et 4) la dynamique des trouée .
En se servant du temps moyen depuis le dernier feu comme un proxy du cycle de feu moyen, ceJiains chercheurs ont proposé une approche de modéliser les proportions théoriques des divers stades de d 'veloppemenl de la forêt pour des ten·itoire dont le paysage serait en équilibre avec le cycle naturel du feu (Bergeron et al, 1999; Harvey et al., 2002). Ceux-ci défini ent alor ces stades de développement par cohor!e slruclurale. La succession forestière y est caractérisée par trois stades larges de développement dans la ucces IOn forestière auxquels certaines combinai ons de structure et composition sont a sociées. Notons que, à l'exception de la 11"0 cohorte de Bergeron el Harvey (1997), le concept de« cohorte structurale » de ces auteurs ne se concorde pas exactem nt avec le sen du lenne « cohorte» qui réfère généralement à un groupe d'arbres qui s'est établit à peu pré en même temps ou dont la croissance libre a été initiée à la suite d'une perturbation majeure (Gauthier el al., 2008).
Sur site mésique, la « 1re cohorte » est généralement composée de peupliers fau x-trembles et de bouleaux blancs se régénérant de façons végétatives après feu, dans la mesure où ce e pèce étaient déjà présentes dans le peuplement d'origine (Bergeron et Dubuc, 1989; Bergeron et Harvey, 1997; Bergeron, 2000). Ces espèces dominent rapidement la ca nopée en raison de leur fort taux de croissance, comparativement à celui des conifère . La première cohOJie est composée d'arbres qui régénèrent le peuplement et dominent la canopée immédiatement aprè la perturbation, souvent des suites d'un feu sévère. Les peuplement immatures et matures de première cohorte sont généralement relativement hom gènes et équiens, en pessière du moins. Toutefoi , une proportion variable d'arbres vétérans ayant
7
survécu à la pe11urbation anté1·ieure peut contribuer à complexifi r la structure d'âge el à diversifier la régénération ubséquente (Gauthier el al., 2008).
Une fois la « 1re cohorte» équienne complètement établie, l'éclaircis ement naturel des
tiges débute environ 70 ans après l'établissement du peuplement, dépendammenl de la
productivité du site, en raison de l'intense compétition envers le ressources environ11antes :
c'est le tade d'exclusion de tiges (Chen et Popadiouk, 2002). La moitalité des arbres de la « 1re cohoite}) engendre des trouées dont la taille influence la quantité de lumière qui atteint le sol. Une faible taille des trouées rend difficile l'établissement d'une« 2e c hotte» de feuillus intolérants à l'ombre el peu d'entre eux parviendront à se hisser en haut de canopée et à composer la 2e coho11e.
L'étape de la transition de la canopée s'enclenche alors que la proportion de feuillus
intolérants dominants diminue, souvent dû à la mortalité «densité indépendante». Ceux-ci sont alors remplacés par des conifère tolérants à l'ombre d'emblé préétablis en sous-canopée, mais upprimés en raison des conditions ombragées. Ils parvie11J1ent alors à croître librement en profitant des ouve11ures créée par la mort d'individus dan la strate arborescente. Par ailleurs, en se décomposant graduellement, les débris 1 igneux au sol, a socié à la mortalité de arbres de la « 1re cohorte », devien11ent des substrats de germination favorables à l'établissement d'une Je cohotte de conifère tolérants à l'ombre
(Messier el al., 1999; Robert el al. 2012). Le peuplement dit de« 2e coh011e »est alor mixte
et souvent à dominance résineuse. La rapidité avec laquelle un 1 euplement feuillu pion_nier évolue vers un peuplement mélangé ayant une structure moins régulière varie largement
(Chen et Popadiouk, 2002). On as ume généralement que la seconde cohorte débute entre 75
et 125 ans après la pe1iurbation initiale (Bergeron el al., 1999; Harvev el al., 2002) mais, en réalité, cette fenêtre de transition est probablement plus étendue.
Finalement, en absence prolongée de perturbation majeure, la «Je cohotte » est caractéri ée par une dynamique des trouées opérant à une échelle fine (Chen et P padiouk,
2002). Park et al., (2005) ont observé que ces processus tendent à favori er de fortes
associations, résultant du replacement du peuplement après feu et de la dynamique des
perturbations econdaire créant de trouée . De plus, les ouvertures dispersées dans la
ca nopée favorisent le recrutement de nouvelles cohorte de r ·génération et tendent à générer
8
diversité dan la taille des arbres (Paré et Bergeron, 1995). L'abondance de ré meux tend donc à augmenter avec le temps depuis le dernier feu, au détriment des feuillus pionnier (Bergeron et Dubuc, 1989; Bergeron et Dansereau, 1993; Bergeron, 2000). En ab ence de perturbation sévère, la troi ième cohorte est maintenue en place grâce à une balance entre les taux de mortalité et de régénération (Chen el Popadiouk, 2002). En fin de succession, les peuplements sont donc composés d'arbres appartenant à diverses classes d'âge, car leur recrutement résulte de divers évènements perturbateurs (Bergeron, 2000). Il est généralement reconnu que la stmcture inteme des peuplements tend à se complexifier au fil du temps (Park et al., 2005). De plus, ces peuplements de structure irrégulière, avec de multiples espèces d'arbres et de tailles, nt souvent associés à une haute biodiversité (Buongiomo et al., 1994). Cette diversité de substrat permet en outre la présence simultanée d'organismes a sociés au bois à différents stades de dégradation (Saint-Germain, Drapeau el M. Buddle, 2007).
Enfin, de manière générale, on suppose qu'après 150 ans, voire 200 an , il n'y a pratiquement plus de d'arbres vivants de la première cohorte, composée d'es nees piomlières peu longévives, établis i1ru11édiatement après la perturbation d'origine (Harvey et al., 2002). Ces peuplements multi-étagés de deuxième et de troisième cohorl ont des structures d'âges qui peuvent se rapprocher grandement d'une structur inéquien11e.
En utilisant le temps moyen depuis le dernier feu comme proxy du cycle de feu, il est donc possible de prédire, théoriquement, la propo11ion les divers stades de développement -les trois cohorte - que devrait occuper une forêt. La forêt préindustrielle de la région écologique Sa, les Basses-ten·es de l'Abitibi (Saucier et al., 1998), avec un temps moyen depuis le dentier feu de 140 ans, serait composée de 45 à 55% de peuplements de 1re cohorte, 23 à 26 % de peuplements de seconde cohorte et 20 à 30% de peuplements de troisième cohorte (Bergeron et al., 2002; Harvey et al., 2002). Quant à elle, avec un temps moyen d. environ 150 ans, la forêt préindustrielle de la région écologique 6a contiendrait une proportion de forêts mûre et surannées (> 100 ans) de 57%, dont 20% aurait plu de 200 ans (Gauthier et al., 2004). Ces proportions fournissent notamment un indice de la structure d'âge de la forêt, mais aussi de la compo ition et de la structure des peuplements (Harvey et al., 2002).
9
Altérarion de la structure par les perturbations secondaires
Les effets des pe1turbations secondaires sur les caractéristiques des forêts, telles que la tructure et la composition, sont la conséquence de trois principaux traits que représentent lïntervalle, la taille et la sévérité, de moindre ampleur que celle d'une perturbation majeure à l'origine d'un peuplement (Gauthier et al., 2008). Cette enveloppe de variabilité, qui varie d'un événement pe1turbateur à un autre, fait en sorte que les pe1turbations secondaires laissent une empreinte totalement différente du feu sur les peuplements forestiers (Gauthier et al., 2008). Dans tous les cas, elles contribuent à diversifier significativement la structure des peuplements forestiers en provoquant la mort diffuse de nombreux arbres, souvent d'espèces particulièrement vulnérables, par exemple le sapin et la tordeuse des bourgeons de l'épinette, en ouvrant la canopée el en recrutant de nouvelles espèces, majoritairement résineuses (Messier et al., 1999).
En absence prolongée de feu, les épidémies de tordeuse des bourgeons de 1 'épinelle (Choristoneura jitm~ferana [Clem.] ; TBE) deviennent les perturbations cycliques majeures contrôlant la dynamique forestière dans des forêts ayant une composante importante de sapin baumier (MacLean, 1984; Holling, 1992; Morin, Laprise et Bergeron, 1993). La composition en essences des peuplements avant une épidém.ie semble influencer positivement la mortalité due aux épidém.ies de tordeuse (Bergeron et Leduc, 1989). En effet, les épidém.ies de tordeuse surviennent davantage dans les forêts à dominance résineuse, où la concentration d"arbres hôtes, particulièrement le sapin, est élevée, que dans les peuplements mélangés (D' Aoust Kneeshaw et Bergeron, 2004) et feuillus, où leurs effets sont généralement moindres et négligeables, respectivement. La sévérité des épidémies de TBE serait donc atténuée par une composante feuillue croissante (MacLean, 1984; Bergeron et al., 1995; Kneeshaw et Bergeron, 1998). Les conséquences de la TBE sur la structure des peuplements s'expriment par la création de trouées de faible dimension et sur la composition par la perte de croissance, !"affaiblissement et, ultimement, la mo11alité des arbres hôtes (Kneeshaw el Bergeron, 1998).
Dans la région écologique Sa, la majorité des ouvertures recensées par D' Aoust, Kneeshaw el Bergeron, (2004) étaient inférieures à 0,1 ba. De plus grandes ouvertures pouvant atteindre plus d"un ha et un maximum de 6 ha. Les auteurs ont remarqué que la dimension des ouvertures augmente avec 1 'augmentation de l'âge depuis le dernier feu. La distribution des trouées n·est pas aléatoire, les effets de la TBE se faisant ressentir plus
1
0
inten ément dan certain secteur que clan d'autres. Selon le MR (2012), lors de la plus récente épidémie, les effet de la tordeuse ont touché environ 17% du teJTitoire de I'Abitibi -Témi can1Jngue.
Selon le MRN (20 13),
r
extrême ud de la région écologique 6a coïncid avec la limite nordique de la répartition de la tordeuse. Ce territoire est dominé par de vastes pessières, où l'épinette noire, une hôte secondaire défoliée par la tordeuse, est affectée mortellement lors de niveau épidémique (Boulanger et al., 20 12). Ain i, les sapi11ières sont peu abondantes, ce qui résulte n des épidémi s moins sévères (Bergeron et al., 1998). L'influence de la tordeuse sur la tructure et la composition de ces rares sapinières est relativement faible (Bergeron el al., 1998). En général, Harper el al., (2002) ont con talé que l'activité de la TBE est très faible pour la région à l'étude, entre 1 et 2 % des peuplement ont été affecté , ceux-ci étant su1iout dans la classe d'âge 100 à 150 ans où l"abondance de sapin baumier est plus élevée.Les trouée créée par la tord u e pem1ettent à la régénération préétablie de sapin et d'épinette de croitre librement (MacLean, 1984). Le plus grande trou 'e contribuent à maintenir une composante, quoique généralement faible, de feuillu dan les vieux peuplem nt en raison de leur plus grande capacité à s'établir en pleine lumière (Bra sare! et hen, 2006). Selon B rgeron et Dansereau ( 1993 ), le impact de épidémie de tord eu e peuvent e traduire en une régression des peuplements résineux vers des peuplements mixtes décidus en favorisant, par la mortalité elu sapin, le recrutement des espèces intolérantes à !"ombre, cela interrompant ystématiquement la ucce sion naturelle ver le peuplements ré ineux. Au Nouveau-Brunswick, selon la composition et la structure du peuplement avant l'épidémie, la réponse du peuplement peut prendre diverses tangentes qui résulteront toujours en un cycle continu de forêt dominée par le sapin (Baskerville, 1975). En absence prolongée de feu, le thuya, une e p 'ce c nsidérée longévive, non hôte à la torcieu e et tolérante à l'ombre par Bums et l-Iollkala (1990), devrait devenir de plus en plus abondant (Knee baw et Bergeron, 1998; Bras arc! et hen, 2006).
Le épidémies de la livrée des forêt (Malacosoma disslria [Hubner]) ont des impacts périodiques sur le feuillu , essentiellement sur le peuplier faux-tremble et dan une moindre mesure sur le peuplier baumier et sur le bou! au à papier (Chen et Popadiouk, 2002). D'après ooke et Lorenzetti, (2006), cinq épidémie de livr' de forêts ont eu cours durant les 70 dernière années au Québec, entre 1930 et 1939, 1949 et 1954, 1965 et 1 968, 1978 et 1982
Il
pUJ 1999 et 2003, ce qui con spond plus ou moin au cycle de 10 ans as ocié aux populations naturelle de livrée des forêt . Plusieur années consécutives d'infestation par la livrée affaiblissent les peuplier faux-tremble, causant parfois une mort prématurée (Senécal, Kneeshaw et Me ier, 2004). Néanmoin , la mortalité d'arbres hôtes uite à un seule périod de défoliation est rare, bien que ceux-ci puissent devenir plus vulnérable à d'autres stress et inférer la sénescence de l'arbre. La mo11alité directe ou indirecte des peupliers favoriserait par ailleurs la croissance de conifères suivant la fonnation de trouée , ce qui engendrerait une diversification de la structure et de la composition de peuplements (Gauthier et al., 2008). D'autres perturbations ont cours dans la forêt boréale mixte, le plus documentés étant le chablis ainsi que cl 'autre insectes défoliateurs. De manière g 'néralc, ce pet1urbations, de faible ampleur, affaiblissent parfois les arbres et les rendent plus susceptible à d'autres agent qui contribuent à la mort des arbre . Bien quïls inOuencent significativemenl la
dynamique forestière, ce agent perturbateurs sont moins bi n documenté en raison de leur
moindre importance relative.
Analyse de la structure
Traditiotmellcment, la structure interne d'un peuplement 'tait réduite à la structure de
la communauté végétale. Mueller-Dombois et Ellen berg, ( 1974) ont disséqué la structure d'un peuplement en trois composantes : (l) la compo ante verticale représentée par la distribution des tiges par hectare par classe de taille et souvent estimée par le diamètre ou la hauteur des arbre ; (2) la compo ante horizontale composée de la distribution patiale des arbres et (3) la dernière composante, constituée de mesure d'abondance d'espèces.
Kinm1ins, (2004) définit la structure des peuplements, ou plus généralement la structure de la communauté, comme étant « The vertical arrangement of canopy layers and planrs of different !ife forms or the horizontal variation in canopy dos ure and canopy layers
or both. Community structure also inc/udes standing dead trees (snags) and decomposing
logs on the forest jloor (cocu·se ·woody debris) ». La défini ti n d Smith et al., ( 1997) est plutôt détem1inée par des considérations telle que« the variation in species and age classe (or lack of it), the arrangement of different layers or storie. ·of vegetation (usually di(fering a to species), and the distribution of diameter clas es ». L 'OIFQ, (2003) définit la tructure
12
d'un peuplement ». La structure interne des 1 euplements peut s'expnmer de maintes
manières ou d'une combinaison de mesures, par exemple en représentant la distribution
diamétrale des tige , la den ité de couve11, la hauteur, l'étagement ve11ical de la végétation, la
quantité, la tai Ile et 1 'état de décomposition des chicots et les débris 1 igneux au sol
(Staudhammer et LeM a y, 2001 ).
Smith el al. (1997) ont décrit la pratique de la sylviculture comme étant « [. . .} a kind of process engineering or forest architecture aimed at creating structures or developmental sequences thal will serve inlended pwposes, be in harmony with the environment, and withstand foads imposed by environ mental injluences ». Lindenmayer, Frank! in et Fisher, (2006) ont commenté, en référ nee à l'échelle du peuplement, que « the interna/ truc/ure and compo ilion of harvested un ils can have a signif/cant ù?fluence on the degree to which a managee/forest can su tain biodiver ity and maintain an eco.sy lem».
Appliqué à une problématique visant à reconnaitre un nombre restreint de structures
caractéristiques de peuplements fore tiers, de multiples méthode sont envisageables pour
mesurer et classifier la diversité structurale. Dans la plupa1i des cas, la structure est mesurée
par un indice, lui-même estimé à l'aide d'une ou de plusieurs variables indicatrices. Pam1i les variables les plus fréquemment utilisée dans des typologies forestières, on évoque l'espèce, la hauteur ou le diamètre, en relation allométrique avec la hauteur, qui, ensemble, traduisent des différences dans la structure verticale et horizontale d'un peuplement (Staudhammer et
LeMay, 2001 ). D'autre variable comme la urface terrière et le nombre de tiges par hectare,
le couvert de la canopée et la biomasse peuvent également être utilisées avec une certaine
efficacité (Staudhammer et LeMay, 2001). Toutefois, selon Farnden, Moss et Earle, (2003),
les variables les plus évidentes comprennent la di tribution des classes de diamètre par espèce
arborescente et leur arrangement spatial sans aire dom1ée (c'est-à-dire en tenant compte que de la di tribu ti on du diamètre des arbres dans la placette d'échantillon, sans leur position exacte à l'intérieur de cette dernière).
Le choix des indicateurs biologiques de la cliver ité structurale est au si très imponant. Ceux-ci sont regroupés sous trois grandes catégories : ceux basés sur 1' identification de espèces clés, ceux basés ur l'identification cl s Sl111ctures clés et ceux qui intègrent une combinaison des deux (Lindei1J11ayer, Margules et Borkin, 2000). McEihinny er al., (2005) remarquent actuellement un intérêt accru envers les indicateurs des attributs structuraux clés
1
3
parce qu'ils constituent à la fois un ubstitut pratique à la description de la biodiversité et un élément révélateur de la source de bi diversité de ces écosystème forestiers.
Il existe aussi quelques courants de pen ée en ce qui concerne l'élaboration d\me clas ification structurale. Traditioru1ellement, en foresterie, la compo ition d'un peuplement oriente souvent le pratiques sylvicoles à privilégier. On a alors tendance à cerner au préalable des types en ne tenant compte que de la composition puis à les décliner en sous -types structuraux (e.g. École ProSylva). Un autre courant de pensée européen consiste à décrire la structure via une indication sommaire de la répartition du volume sur pied en troi grandes classes de diamètre (Schütz, 1997). Ce diagramme à troi entrées, communément appelé « triangle des tructure >>, dans lequel est intégré les propo1iion relatives de trois catégories de bois (Petits Bois, Moyens Bois et Gro Bois) selon le nombre de tiges, la surface terrière ou le volume. Cette représentation graphique facilite l'approche dynam.ique de l'évolution des peuplement et, d'un point de vue de l'aménagement, J'approche foumit un outil d'estimation des différents produits de transfom1ation (e.g. déroulage, ciage, bois de pâte). Cependant, on doit déterminer arbitrairement le limites de chaque taille de bois pour rendre possible la détermination des types tructuraux (Rosa, 1999).
Au Québec, on retrouve une caracté1isation structurale des peuplements forestiers productifs résultant de photo-interprétation (OIFQ, 2009). Celle-ci est décrite par le paramètre « classe d'âge » qui infonne à la fois sur la structure du peuplement (régulier, iiTégulier ou étagé) el sur l'âge des tiges (équien ou inéquien) qui Je composent. Actuellement, le MRN distingue alors les peuplements (1) équiens de tructure régulière, (2) inéquiens de structure régulière, (3) de structure irrégulière et (4) de structure étagée. es appellations, résultent d'une photo interprétation, en pa11ie vérifiée par l'âge comptée sur les carottes des arbres-études provenant d ïnventaire en forêt, et sont appliquées à la totalité des peuplements forestiers productifs du Québec méridional Bien qu ces c1itères soient facilement évaluable par le photo-interprète, il résume la large variabilité structurale observée en forêt boréale mixte en quelques groupes.
Traditiom1ell ment, la composition des essences dominante (avec le volume de tiges à l'hectare) est le principal détem1inant du traitement ylvicole à privilégier, parfoi complémentée par la structure du peuplement. Toutefois, il est possible d'élaborer une typologie pur ment trucrurale puis d'évaluer lïnfluence de la composition a posteriori. clans
14
la mesure où les changements de composition entrainent de changement de Lructure. En forêt boréale mixte, la succession naturelle de espèces, caractérisée par le recrutement d'une
1re c horte feuillue et équienne, d'une 2e cohorte mixte pui d'une 3e cohor1e à dominance ré ineu e et inéquienne, suggère un lien entre la structure ella composition d'un p uplemenl. Tl est alors possible d'évaluer la relation entre la composition et la structure puis de développer de pratiques sylvicoles à la fois adaptées à la structure des peuplement , ans pour autant négliger leur composition, les deux étant indissociables dans une optique d'aménagement forestier durable.
L'approche privilégiée dans le cadre de la présente étude, inspirée de travaux de Farnden, Moss et Earle, (2003), est basée sur la distribution cumulée relative de la urface teiTière et de la densité de Liges par classe de diamètre. Elle reconnait implicitement le large continuum que représente la structure des peuplements à travers le paysage forestier et y distingue certain patrons en se ba ant uniquement sur la fréquence relative de différentes tai Ile d'arbres (Farnden, Moss et Earle, 2003 ). À partir de ces patrons, le continuum peul être rompu en n'impor1e quel nombre de groupes' con istants d'individus similaires et la variation quasi infinie qu'il existe est alors implifiée et réduite en quelques groupes facilement identifiables (Farnden, Mos et Earle, 2003). Ultimement, cet outil pouiTait servir d'assi e et intégrer plus efficacement : l) des cibles structurales, inspirées de la structure des peuplements naturels, dans le pratiques sylvicoles; 2) la dynamique naturel! dans les traitements sylvicoles; et 3) de objectif tructuraux d'habitats dan le prises de décision
sylvicoles.
L'échelle à laquelle e manifeste l'hétérogénéité structurale varie largement (Dutilleuil, 1993), et conséquemment, 1 'échelle à laquelle on estime les attributs structuraux s'avère capitale. À on origine, la foresterie en forêt boréale canadienne a été fortement influencée par une vision d'exploitation à court tenne où la principale préoccupation visait l'extraction de la re ource ligneuse au moindre coût. Au Québec, cette influence est illu trée par un régime équien d'aménagement, longt mp appliqu' sy tématiquement à l'ensemble du terTitoire fore Li r boréal (Boucher, De Grandpré et Gauthier, 2003).
1
En terme stati tique, les groupes résultants sont également nommé types. Puisque ces types regroupent des patrons structuraux, on les nommera «types structuraux».
15
Dan une optique de caractérisation le la structure des peuplements, une t lie vision homogénéisatrice des peuplement e t réaliste lorsque le ten·itoire soumis à un aménagement régulier st déjà relativement homogène (Puettmann, Coate et Me sier, 2009) et, clans une telle si tua ti on, la petite upcrfïcie des placettes-échantillons standard ( 400 m]) erait probablement adéquate. Toutefois, une telle démarche est peut-être moin judicieuse lorsque
le territoire en que tion e t caractérisé par une fo11e hétérogénéité structurale, où la taille de échantillons pourrait être rehaus ée.
À partir de « stmcture-area-curves », Zeru1er, (2005) a investigué J'hétérogénéité de
divers attributs structuraux dans dix peuplements ancien de sapin Douglas situés en Orégon. Aux échelles de 100, 500, 1000, 2500 et 5000 m2, il a réalisé de multiples itérations en
évaluant ces attributs pour en arriver à la conclu ion que la variation de forêts les plu complexe devenait table à partir de placettes d'une superficie de 2500 m2 et plus.
Naturellement, cette uperfïcie minimale varie selon la complexité des forêts. Dans l'optique
d'appliquer des pratiques sylvicoles à l'échelle du peuplement, en fonction des att1ibuts
électionnés, de tels ré ultats questionn nt le choix de la taille minimale de placett es-échantillons dans les forêts de sapin Dougla en Oregon.
La présente étude ne concerne que les peuplements mélangés et exclus les peuplements
feuillu purs, ouvent équien , et conifériens purs, souvent inéquiens. Celte exclu ion vi cà
exclure les diverses trajectoires successionnelle qui ne pa ent pa par l'évolution de
peuplements mélangés. De plu , l'étude exclue les peuplements de ela e d'âge de 30 ans et moin afin de ne con erver que les p uplements rendus à l'âge d'exploitation. Ces exclu ions
simplifient grandement le gradient structural à analyser mais, rend potentiellement les distinctions entre Je type plus nuancées. éanmoin , en considérant la forte hétérogénéité
structmale qui caracté1ise les peuplem nt boréaux mixtes (Park et al. 2005), nous sommes en mesure de nou questionner sur la convenance de la taille des placettes-échantillons (d'une
taille conventionnelle de 400 m2) et de la capacité de la typologie à décrire la tructure de superficies forestières plus grandes.
CHAPITRE T
Typologie structurale des peuplements mélangés de la forêt boréale de l'Ouest du Québec
Sylvain LAROUCl-IE1, lain LEDUC1, BrianD. HARVEY2, lan MOSS' 1
Chaire industrielle CRS G-UQAT-UQAM en Aménagement Forestier Durable. Université du Québec à Montréal. 141, Avenue du Président-Ke1medy,
Montréal, Qc. H2X 1 Y 4 2
Chaire industrielle CRSNG-UQAT-UQAM en Aménagement Forestier Durable. Université du Québec en Abitibi-Témiscamingue. 445, bou!. de l'Université,
Rouyn-Noranda, Québec. J9X 5E4 3
ForestTree Dynamic Ltd. 1458, Kingsview R ad, Duncan, Briti h-Columbia. V9L 5P l
1.1 Résumé
La structure inteme des peuplements mélangés bor' aux est un attribut clef, entre autre pour le maintien de la biodiver ité et de la productivité forestière. Dans la prise de décision concernant le traitement sylvicole à appliquer dans ces peuplements, leur structure fine pouiTait être considérée dans une optique de diversification des pratiques sylvicoles. Afin de répondre à ce besoin, le principal objectif de notre étude visait à élaborer une typologie structurale fine des peuplement mélangés d"une région de la forêt boréale méridionale de
l'Est canadien en se basant sur la distribution de la densité et de la surface teiTière des arbres commerciaux. L'analyse a été effectuée sur 461 placettes-échantillons temporaires provenant du Mini tère des Ressources naturelles (MRN) elu Québec. Une foi la typologie élaborée, le types structuraux de référence ont été ordon11és en uivant leur ucces ion structurale théorique afin d'évaluer a posteriori l'influence de la composition et des perturbation secondaires sur la structure forestière. Le résultats suggèrent qu'il est pos ible de résumer le gradient structural des peuplements mélangés du territoire cl' étude en huit types tructuraux à l'échelle de la placette-échantillon (400m2). De relations entre la structure des peuplements et la composition de même que les effet de la tordeuse des bourgeons de l'épinette ont pu être caractérisées. Nos résultats suggèrent un cadre sylvicole duquel il sera possible de prescrire des cibles d'aménagement adaptées à la structure interne des peuplements clans un objectif de mitiger les conséquences de !"aménagement sur la biocliversité et la productivité des forêts.
Mots-clés: Forêt boréale mixte: Structure forestière: Composition fore tière; Tordeuse des bourgeons de !"épinette: Aménagem nt fore-ti r éco ystémique.
1
9
1.2 Introduction
Les paysages boréaux à peuplements mélangé sont modelés par des régm1 s de
p rturbations (Van Wagner, 1978; Johnson et Gut ell, 1994; Angelstam et Kuuluvainen, 2004) généralement dominé par le feu (Johnson, 1992; Payette. 1992). En Am' ri que du Nord, ce régime circonscrit une mosaïque de peuplements qui diffèrent les uns des autres par leur âge, leur structure inteme et leur composition (Dix et Swan, 1971; Rowe et Scotter,
1973; Johnson, 1992). Le jeunes forêts y ont ouvent a ociées, une canopée dense et unifonne composée de feuillus intolérants. Avec le temps, et sous 1 'action de perturbations
eco nd aires ( e.g. Kneeshaw et Bergeron, 1998; Cooke t Lorenzetti, 2006), le individus qui dominent la canopée seront graduellement remplacés via une dynamique de trouées
(Bergeron, 2000; Chen et Popadiouk, 2002). En l'ab ence du feu, les peuplement équien de
feuillus intolérants peuvent évoluer vers des peuplements de structure inéquien11e et de composition mixte (Chen et Popadiouk, 2002). Ces peuplements ont alors caractérisés par une f011e hétérogénéité structurale, en raison des nombreux proce SU qui oi1t COUrS à ce stade de la uccession (Park et al., 2005). D'une par1, cette cl1ronoséquence suggère une relation
entre le temps écoulé depuis le plus récent feu et les changements compositionnels (Berger n ,·
et Dubuc, 1 989; Frelich et Reich, 1995) et structuraux (Chen et Popadiouk, 2002). D'autre
part, dans la portion mélangée de la forêt boréale, la mortalité des ré ineux mature provoquée par les épidémies de la tordeuse des bourgeons de l'épinett (Blais, 1983: Morin,
Laprise et Bergeron, 1993) pourrait modifier la relation entre la compo ition t la structure des peuplements ainsi qu'entre le temps écoulé depuis le dernier feu et ces deux descripteurs
des peuplements.
Toutefoi , ju qu'à récemment, la cormai sance des régimes de feu qui caractérisent la forêt boréale était très fragmentaire (Bergeron et al., 2002). Ce manque de compréhension a
souvent conduit à des généra li ations eJTOnées, telle que 1 'utili ation non jus ti fiée d la coupe
à blanc sur l'en emble de la forêt boréale, celle-ci étant fondée sur l'hypothèse que le régime
de [eu est caractérisé par la pré ence de vaste et fréquent feux évères produi ant de
p uplements équiens sur la vaste majorité du territoir (Bergeron et al., 2002). Ain i, pendant des décennies, la perception de la forêt boréale comme une forêt jeune, équienne et homogène a souvent pem1is de justifier le recours à un eu! mode d'aménagement pour l'ensemble de la forêt boréale. Avec la venue de nouvelles coru1aissance sur la dynamique
20
des forêts boréale mixtes de rest du Canada et leur structure d'âge, il est devenu de plus en plus évident que les courts cycle de feu ne nt dominants que dan certaines portions du territoire, où le conditions régionales ont favorables (Bergeron el al., 2001).
onséquernment, le pratiques sylvicole résultantes ignoraient, à plusieurs égards, la dynamique naturelle intrinsèque aux écosystème forestiers, elle qui implique une transition des types de peuplements (Bergeron el Harvey, 1997). Ce constat explique en partie le manque d'outils disponibles pour classifier efficacement la stmcture des forêts qu 'bécoise (Boucher, De Grand pré et Gauthier, 2003; Grondin et al., 201 0). En effet, la typologie actuelle du MERN ne distingue que les peuplements à structure régulière, inégulière et étagée et ces types s'appliquent à l'ensemble des peuplements du ten·itoire forestier productif québécois. Or, au fil des rotations, des modifications dans la composition du paysage pourraient entraîner de lourde répercussions, notamment ur la diversité biologique (Boucher, De Grandpré et Gauthier, 2003), la productivité des peuplements (Légaré, Paré et Bergeron, 2004) et le propriétés du sol (Brais el al., 2004).
L'aménagement forestier écosy tém.ique, désom1ais en vigueur par la Loi sur 1 'aménagement durable du territoire forestier (Québec, 20 13), a été développé dan 1' optique de maintenir des éco ystèmes sains et résilients en m.inimisant les éca1ts entre les paysages aménagées et naturels (Landres, Morgan et Swanson, 1999; Gauthier el al., 2008). Selon le paradigme écosystém.ique, le maintien de lïntégrité de écosystèmes passe par la préservation des patrons structuraux et c mpositionn ls à 1 ïntérieur des limites de la variabilité historique régionale, produite sous un régime de perturbation naturel (Mladenoff el
al., 1993; Landres, Morgan et Swan on, 1999; Lindenmayer, Margules et Botk.in, 2000). Ce faisant, l'implantation d'une telle stratégie nécessite avant tout une connaissance approfondie des fonctions et processus écosystémiques (Christensen el al., 1996), mai également le développement d'outils pour caractériser efficacement ces écosystèmes et établir des cibles d ·aménagement adaptées (Harvey et al., 2002; Romp ré el al., 20 l 0).
La structure, un attribut clef des écosystèmes boréaux de rE t du Canada décrit l'anangement veitical et horizontal des arbre et diver e manières de clas ifïer la diversité tructurale des peuplements ont été explorées (voir Staubammer et LeMay, 200 l ). Selon Farnden, Moss et Earle, (2003), les caractéri tique tructurales les plus évidentes proposent une distributi n de arbres par classe diam 'traie ainsi que la disposition de ces arbre dans
21
une atr donnée. Ces caractéristiques communes sont en outre faciles à reconnaitre et à quanti fier. De plu , elles intègrent un certain nombre d'autres caractéristiques, telles que la pénétration de la lumière au travers de la canopée, le développement des espèces sous celle -ci, le défilement des tiges et l'agencement des fibres du bois (Mos , 2012). Collectivement, ces attributs sont d'une grande importance pour l'aménagement, en ce qui conceme le habitats (Drapeau el al., 2000; Kash ian et Barn es, 2000; Payer et Harisson, 2000), le volume et la qualité du bois (Moss, 20 12).
Notre p1incipal objectif (l) vis à décrire et à illustrer une approche de classification de types structuraux de peuplement mélangé d'une région de la forêt boréale méridionale de l'Est du Canada. Une fois la typologie structurale élaborée, on propose (2) un enchainement temporel des types représentant la chronoséquence forestière et une évaluation a posreriori l'influence (3) de la compo ition et (4) de perturbations secondaires ur la structure interne des peuplements. Suivant ces objectifs, nous propo ons les hypothèses suivantes : (Hl) que le gradient structural des peuplements mélangés du territoire d'étude est statistiquement et écologiquement résumable en quelque types structw·aux distincts et con istant ; (H2) que la composition spécifique d"un peuplement déterminera sa structure; (H3) conséquemment les types structuraux 'ordonnent le long d'une équence temporelle et (H4) que les pertw-bations secondaires, en provoquant la 1110I1alité dïndividus hôtes, peuvent altérer la relation structure-composition établie à !'I-I2.
22
1.3 Aire d'étude
Notre aire d'étude est clélim.itée par les régions écologiques Sa- Plaines de l'Abitibi et
6a - Plaines elu lac Matagami (réf Figure 1.1 ). Il s'agit d'une vaste plaine argileuse située à
environ 300 m d'altitude, dont le bassin versant est orienté pre qu'entièrement vers la baie
James. Les sédiments glaciolacustres constituent des vestiges de 1 'ancien lac proglaciaire
Ojibway (Vincent et Hardy, 1977). La région écologique 5a (Plaines de l'Abitibi) appartient
au sous-domaine bioclimatique de la sapinière à bouleau blanc de I"Ouest el couvre une superficie de 26 895 km2 (Grondin, Blouin et Racine, 1998). La portion méridionale de la
région écologique est légèrement ondulée tandis que la portion septentrionale est davantage
plane (Gauthier, De Granclpré et Bergeron 2000). Les dépôts d'argiles glaciolacustres sont
omniprésents, suivis des dépôts organiques et des tills minces (LaqueJTe, Harvey et Leduc,
20 Il). La température moye1me annuelle est de 1 °C. La saison de croissance s'étend ur 160 jours et les précipitations annuelle totales sont de 800 à 900 mm (Grondin, Blouin et Racine, 1998). La région écologique 6a (Plaines clq lac Matagami) est située clans le sous-domaine bioclimatique de la pessière à mousses de l'Ouest (Saucier et al., 1998). Elle couvre une
superficie de 49 099 km2 Le relief est relativement plat et les tourbières y abondent. Les sol
orgamque ont dominants, suivis de prè par les dépôt glaciolacustre et les till (Robitaille
et Saucier, 1998). La température moyenne varie entre 0
o
c
et -2.5 °C. La saison decroissance s'étend sur 150 jour el les précipitations totales sont de 700 à 800 mm (Robitaille et Saucier, 1998).
L'histoire postglaciaire (Mes aoud, Bergeron et Leduc, 2007), la dimension et la sévérité des feux (Bergeron et al. 2004) ainsi que la diminution du potentiel reproducteur du sapin vers le nord (Mes aoud, Bergeron et Leduc, 2007) expliquerait la transition de la sapin.ière vers la pessière autour du 49e parallèle nord. Dans la sapinière, la plus forte
abondance de feuillus intolérant , la présence de plans d'eau, de routes et de ten·es agricole agissent à titre de coupe-feu et contribuent à réduire la dimension et la sévérité des feux
(Lefo1i, Gauthier et Bergeron. 2003; Laguerre, Harvey et Leduc, 2011). Invers ment, dan la
pessière à mousses de l'Ouest, dont la vocation agricole est marginale, pré ente un régime de feux peu affecté par 1" homme (Laquen·e, Harvey et Leduc, 20 Il) Pour ces raisons, el aussi parce que la sapinière e t d'emblée plus cliver ifiée, la région écologique 5a contient davantage de peuplement mélangés que la région écologique 6a (Figure l.l ).
81"W 52•N
----
1--\
SO"N•
0 81"W Figu1·e 1.1 Villes PET 50 km 80"W f~ i ~~~ -: ·,j 1i
l
! i i i i i!
79"W 79"W.
.
78"W 77"W6a
<.
.·
1a·w 77"WRépartition des placette -échantillons temporaires utilisées dan d'étude (Sa: n = 396 t 6a: n = 65).