Architecture génétique des caractères reliés au bois, à
la croissance et à la résistance contre la tordeuse des
bourgeons de l’épinette chez l’épinette blanche
Thèse
Mebarek Lamara
Doctorat en sciences forestières
Philosophiae doctor (Ph.D.)
Québec, Canada
Architecture génétique des caractères reliés au bois, à
la croissance et à la résistance contre la tordeuse des
bourgeons de l’épinette chez l’épinette blanche
Thèse
Mebarek Lamara
Sous la direction de :
Jean Bousquet, directeur de recherche
John MacKay, codirecteur de recherche
Résumé
Les analyses d’association visent à mieux comprendre l’architecture génétique des caractères quantitatifs comme ceux reliés à l’adaptation, la croissance et la structure du bois, et mettre en évidence les gènes qui les contrôlent. Ces connaissances sont utiles pour notamment mieux appréhender le potentiel de résilience dans le contexte de changements climatiques, ainsi que pour la conservation et l’amélioration génétique des arbres forestiers. Cette thèse porte sur l’épinette blanche (Picea glauca [Moench] Voss) avec deux objectifs principaux : 1) développer une compréhension plus intégrée des caractères quantitatifs, et ; 2) développer des stratégies permettant de mieux exploiter les analyses d’association face aux contraintes génétiques et statistiques liées à leur utilisation chez les arbres forestiers. La thèse s’appuie sur deux études d’association qui portent respectivement sur la croissance et les caractéristiques physiques du bois (chapitre 2), et la résistance contre la tordeuse des bourgeons de l’épinette (TBE, Choristoneura fumiferana Clemens) (chapitre 3). Nous avons d’abord analysé les caractéristiques du bois ainsi que la croissance en utilisant un seuil de significativité statistique permissif (P < 0,05; sans correction pour tests multiples) pour maximiser la découverte de gènes comportant des SNPs significatifs. Selon les caractères étudiés, entre 229 et 292 gènes ont été identifiés, qui ont ensuite été soumis à des analyses complémentaires portant sur leurs annotations fonctionnelles et leur expression tissulaire. Les gènes associés aux caractères du bois se sont montrés surreprésentés dans un groupe de co-expression préférentiel au xylème secondaire. Une analyse de réseau de ce groupe de gènes a permis d’identifier certains facteurs de transcription de type MYB et NAC ayant un niveau de connectivité élevé, dont notamment PgNAC-7 qui était le gène ‘hub’ le plus connecté, suggérant un rôle dans la régulation des caractères physiques du bois. L’étude des caractères de défense contre la TBE s’est appuyée sur trois méthodes d’analyse d’association considérant soit un locus à la fois, plusieurs loci, ou encore plusieurs caractères simultanément. Les analyses ont permis d’identifier 33 gènes ayant des fonctions métaboliques variées et expliquant une proportion élevée de la variation phénotypique observée. Des corrélations phénotypiques faiblement positives ont également été observées entre la croissance et les facteurs de résistance contre la TBE, indiquant l’absence d’un compromis métabolique entre ces caractères. Nos résultats indiquent que les stratégies proposées dans la présente thèse pourraient être appliquées à d’autres caractères quantitatifs et permettre de développer des connaissances utiles pour mieux comprendre l'architecture génétique des caractères complexes chez les arbres forestiers.
Abstract
The general aim of association analyses is to decipher the genetic architecture of quantitative traits such as those related to adaptation, growth and wood structure, and to highlight the genes that control them. This knowledge is useful to better understand the potential for resilience toward climate change, and for the genetic conservation and improvement of forest trees. This thesis focuses on white spruce (Picea glauca [Moench] Voss) and has two main objectives: 1) develop a more integrated understanding of quantitative traits, and; 2) develop strategies to better exploit association analyses despite the genetic and statistical constraints related to their use in forest trees. The thesis is based on two association studies, which focus respectively on growth and wood physical traits (Chapter 2), and resistance to spruce budworm (SBW, Choristoneura fumiferana Clemens) (Chapter 3). We first analyzed wood and growth traits using a permissive threshold of statistical significance (P < 0.05, without correction for multiple tests) to maximize the discovery of genes carrying significant SNPs. For each of the traits studied, between 229 and 292 such genes were identified, which were then subjected to further analyses based on their functional annotations and their tissue expression. The genes associated with wood traits were over-represented in a co-expression group that is preferential to secondary xylem. A network analysis of this group of genes allowed identifying certain transcription factors of MYB and NAC types with a high level of connectivity, including PgNAC-7 which was the most connected hub gene, suggesting a role in the regulation of wood physical traits. The study of SBW defence traits was based on three methods of association analysis considering one locus at a time, several loci, or several traits simultaneously. The analyses identified 33 genes with various metabolic functions, which explained a major proportion of the observed phenotypic variation. Weakly positive phenotypic correlations were also observed between growth and resistance traits against SBW, indicating the absence of a metabolic trade-off between these traits. Our results indicate that the strategies proposed in this thesis could be applied to other quantitative traits to develop a better understanding of the genetic architecture of complex traits in forest trees.
Table des matières
Résumé ... iii
Abstract ... iv
Liste des tableaux ... viii
Liste des figures ... ix
Remerciements ... xi
Avant-propos ... xii
Chapitre 1. Introduction générale ... 1
1.1 Introduction ... 1
1.2 Les caractères quantitatifs et leur contrôle génétique ... 2
1.2.1 Phénologie et croissance ... 2
1.2.2 Qualité du bois ... 4
1.2.3 Résistance aux insectes et développements récents concernant la tordeuse des bourgeons de l’épinette ... 5
1.3 Corrélations entre les caractères ... 7
1.4 Analyses des bases moléculaires des caractères quantitatifs ... 8
1.4.1 Objectifs de dissection génétique des caractères quantitatifs ... 8
1.5 Méthodes d’analyses moléculaires des caractères quantitatifs ... 11
1.5.1 Approches classiques d’identification de QTL ... 11
1.5.2 Les analyses d’association génétique ... 12
1.6 Fonctions, rôles biologiques et expression des gènes identifiés par les études d’associations .. 17
1.7 Mise en contexte du projet de recherche ... 20
1.7.1 Développement de la génomique forestière et ressources disponibles ... 21
1.8 Hypothèses et objectifs de recherche ... 22
1.8.1 Génétique des caractères de croissance et de qualité du bois ... 22
1.8.2 Génétique des caractères de résistance à la tordeuse des bourgeons de l’épinette ... 24
1.9 Références ... 25
Chapitre 2. ... 39
Genetic architecture of wood properties based on association analysis and co-expression networks in white spruce ... 39
Résumé ... 40
Summary ... 41
2.1 Introduction ... 42
2.2 Materials and methods ... 44
2.2.1 Plant material and tissue sampling used for association analysis ... 44
2.2.2 Phenotypic data ... 44
2.2.4 Estimation of quantitative genetic parameters ... 45
2.2.5 Association analysis ... 46
2.2.6 Microarray experiments and gene expression network ... 46
2.2.7 Functional annotations and enrichment analyses ... 47
2.3 Results ... 47
2.3.1 Quantitative genetic analysis ... 47
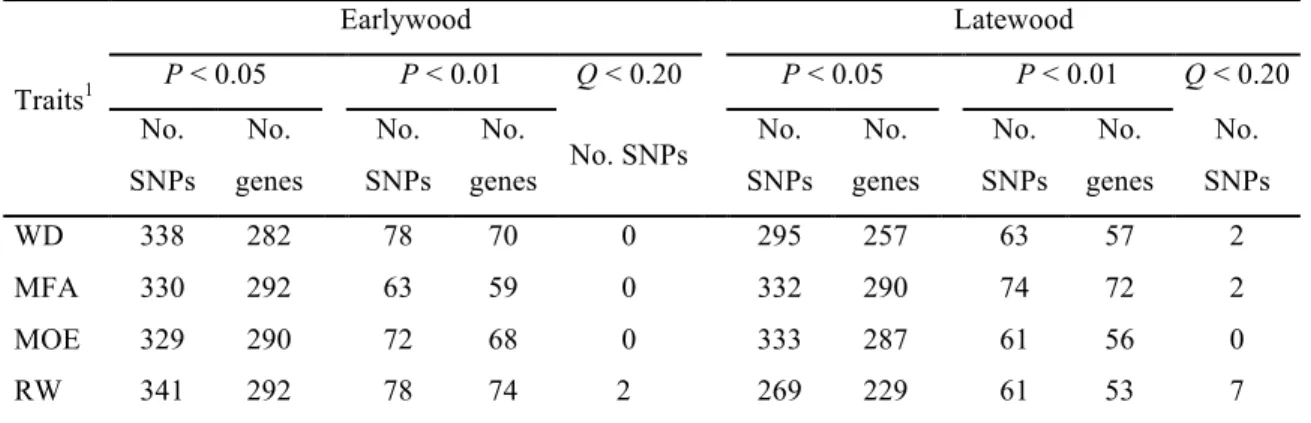
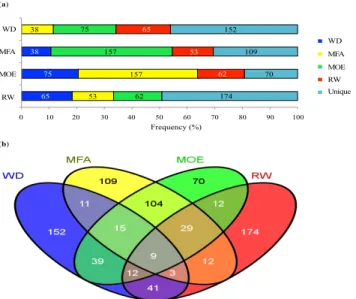
2.3.2 Association study ... 48
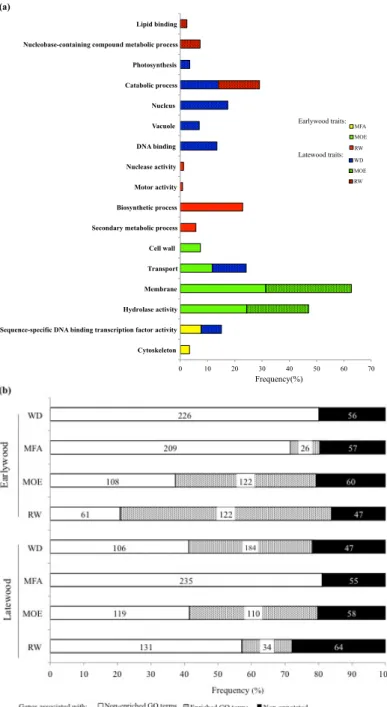
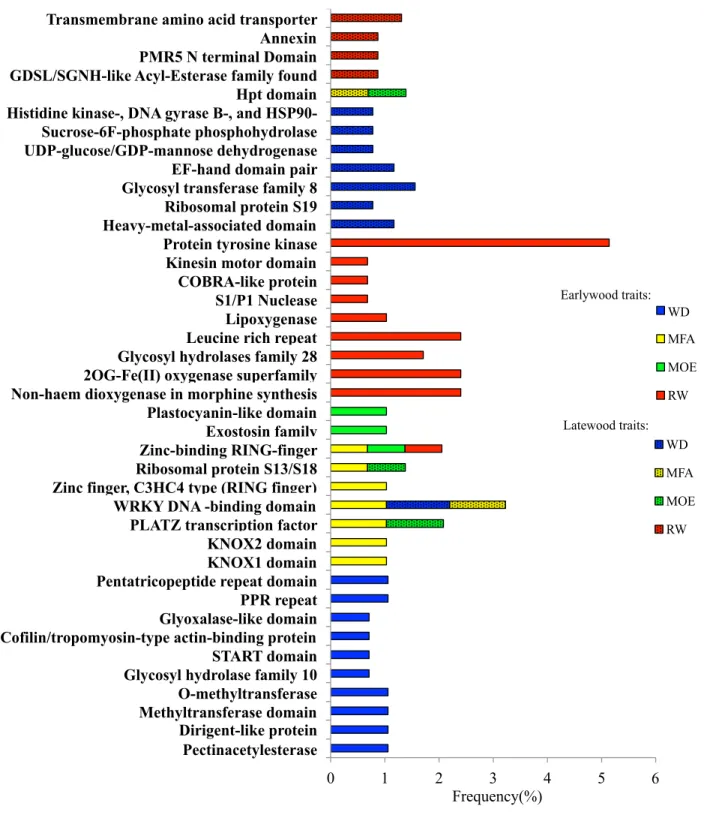
2.3.3 Gene ontology and Pfam enrichment analyses ... 52
2.3.4 Gene expression ... 53
2.3.5 Gene network reconstruction ... 56
2.4 Discussion ... 62
2.4.1 Genes associated with physical wood traits ... 62
2.4.2 Relationship between genetic architecture and quantitative genetic parameters ... 62
2.4.3 Gene functions associated with physical wood traits ... 63
2.4.4 Expression profiles and reconstruction of a wood formation gene network ... 65
2.5 Conclusion ... 67
2.6 Acknowledgements ... 68
2.7 References ... 68
2.8 Supporting methods ... 75
2.8.1 Methods S2.1 Candidate genes selection ... 75
2.8.2 Methods S2.2 Information and formulas used for estimation of quantitative genetic parameters 75 2.8.3 Methods S2.3 The hypergeometric test used for the evaluation of the over- and under-representation of candidate and significant genes in the co-expression groups ... 76
2.9 Supporting figures ... 78
2.10 Supporting tables ... 81
2.11 Supporting references ... 81
Chapitre 3. ... 83
Association genetics of acetophenone defence against spruce budworm in mature white spruce ... 83 3.1 Résumé ... 84 3.1.1 Contexte ... 84 3.1.2 Résultats ... 84 3.1.3 Conclusion ... 84 3.2 Abstract ... 85 3.2.1 Background ... 85 3.2.2 Results ... 85 3.2.3 Conclusion ... 85 3.3 Introduction ... 86
3.4 Materials and methods ... 88
3.4.2 RNA extraction and transcript determination assays ... 89
3.4.3 Acetophenone extractions and determinations ... 89
3.4.4 Genotypic data ... 90
3.4.5 Simulations ... 90
3.4.6 Association analyses ... 91
3.4.7 Trade-offs between defence and growth traits ... 92
3.5 Results ... 92
3.5.1 Phenotypic variation ... 92
3.5.2 Simulations ... 94
3.5.3 Identification of genes associated with defence traits ... 94
3.5.4 Functional annotations and expression of genes associated with defence traits against spruce budworm ... 96
3.5.5 Defence-growth trade-offs ... 99
3.6 Discussion ... 101
3.6.1 Candidate genes associated with defence traits ... 101
3.6.2 Molecular basis of acetophenone accumulation ... 102
3.6.3 Trade-offs between defence and growth traits ... 106
3.6.4 Practical applications of identifying genes associated with defence traits ... 107
3.7 Acknowledgements ... 107
3.8 References ... 108
3.9 Supporting figures ... 118
3.10 Supporting references ... 119
Chapitre 4. Conclusion générale ... 120
4.1. Principaux résultats et conclusions ... 120
4.1.1 Caractères de croissance et de qualité du bois ... 120
4.1.2 Caractères de défense contre la tordeuse des bourgeons de l’épinette ... 123
4.2. Analyse critique ... 126
4.3. Perspectives de recherche ... 128
4.4. Applications pratiques ... 130
Liste des tableaux
Tableau 1.1 Synthèse des résultats de plusieurs études de cartographie de QTL des caractères quantitatifs par chez les arbres forestiers ... 14 Tableau 1.2 Synthèse des résultats de quelques études d'association par l’approche de gènes candidats chez les arbres forestiers ... 19 Table 2.1a Genetic (above diagonal) and phenotypic (below the diagonal) correlations between traits and individual trait narrow-sense heritabilities (on the diagonal, in bold) in white spruce. ... 49 Table 2.1b Genetic (bold) and phenotypic correlations between and within traits and between earlywood and latewood in white spruce. ... 49 Table 2.2 Number of significantly associated SNPs and genes after association testing with earlywood and latewood traits in white spruce, and number of significant SNPs after correction for false discovery rate (FDR) (Q < 0.20). ... 50 Table 2.3 Distribution in co-expression groups of white spruce1 of total frequencies of genes tested and frequencies of significantly associated genes after association testing. ... 57 Table 2.4 Numbers of wood associated genes with xylem preferential expression and subsets selected to reconstruct a co-expression network. ... 58 Table 2.5 The 20 hub genes and their functions in the wood co-expression network in white spruce.
... 59 Table 3.1 Summary statistics of constitutive defence traits in the white spruce association population. ... 93 Table 3.2 Simulation results of detecting significant1 SNP-trait associations using SLMM and MLMM approaches ... 94 Table 3.3 Single nucleotide polymorphisms (SNPs) associated with defence traits in white spruce using SLMM and MLMM approaches1, and their combined percentage of phenotypic variation explained (PVE). ... 95 Table 3.4 Number of single nucleotide polymorphisms (SNPs) associated with defence traits in white spruce using the multi-trait mixed model (MTMM) approach. ... 95 Table 3.5 Phenotypic correlations between defence traits and between defence and growth traits in white spruce . ... 99 Table 3.6 Factor loadings of the three first principal components (PCs) for all defence and growth traits analysed in this study. ... 99 Tableau 4.1 Corrélations phénotypiques entre les caractères de défense contre la tordeuse des bourgeons de l’épinette et la qualité du bois chez l’épinette blanche. ... 130
Liste des figures
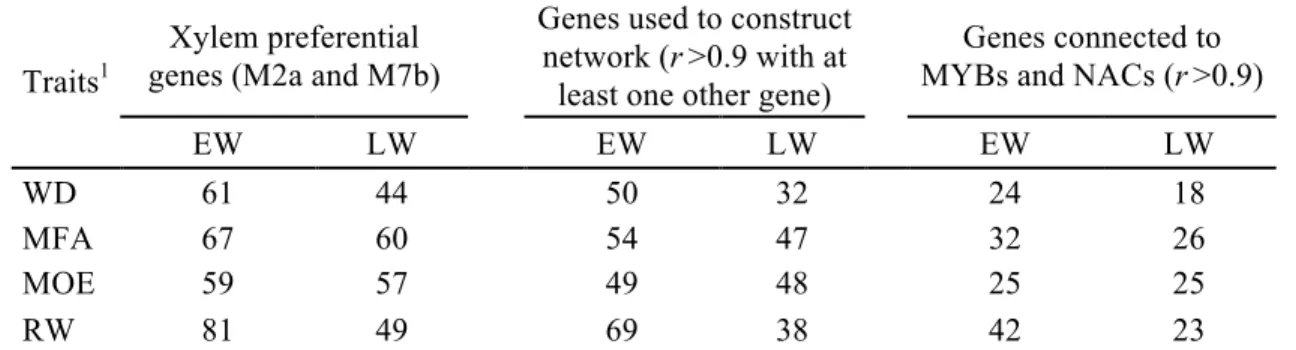
Figure 2.1 Flowchart of the different analyses and data sources.. ... 45 Figure 2.2 Numbers of significantly associated genes after association tests, specific to earlywood (EW), latewood (LW) or shared (in common) to EW and LW for wood density (WD), microfibril angle (MFA), modulus of elasticity (MOE) and ring width (RW) in white spruce.. ... 51 Figure 2.3 Overlap among sets of significantly associated genes after association testing (P < 0.05) between the different traits as determined for earlywood in white spruce.. ... 51 Figure 2.4 Functional annotations and gene ontology analysis of significant genes associated with wood traits (WD, wood density; MFA, microfibril angle; MOE, modulus of elasticity; RW, ring width) in white spruce.. ... 54 Figure 2.5 Enrichment analysis of Pfam protein domains in significantly associated genes after association testing in white spruce for all wood traits (WD, wood density; MFA, microfibril angle; MOE, modulus of elasticity; RW, ring width) in earlywood and latewood.. ... 55 Figure 2.6 Co-expression network and expression profile of 93 significantly associated genes highly expressed in the xylem of white spruce.. ... 60 Figure 2.7 Map of the earlywood trait associations (wood density (WD); microfibril angle (MFA); modulus of elasticity (MOE) and ring width (RW)) across the gene co-expression network. .. 61 Figure S2.1 Plots of the 1694 white spruce trees on the plane of the two first eigenvectors derived from the principal component analysis (PCA).. ... 78 Figure S2.2 Overlap among sets of significantly associated genes (P < 0.05) between the different traits as determined for latewood. ... 78 Figure S2.3 Gene co-expression groups in white spruce (Picea glauca) according to Raherison et al. (2015).. ... 80 Figure 3.1 Histogram and density plot showing residual distribution in all traits. A, C, E, G before normalization and B, D, F, H after normalization. ... 93 Figure 3.2 Heatmap of tissue-specific expression patterns of significantly associated genes and functional annotations. Expression data are from the PiceaGenExpress database (Raherison et al., 2012).. ... 98 Figure 3.3 Biplots of the three first components of principal component analysis of defence and growth traits.. ... 100 Figure 3.4 Venn diagrams illustrating the extent of overlaps of significantly associated genes between defence traits and between defence and growth traits at P < 0.05 .. ... 100 Figure S3.1 Heatmap of tissue-specific expression pattern of candidate genes involved in phenylpropanoid pathway used in this study and their functional annotations.. ... 118
Remerciements
Je remercie John MacKay qui a agit a titre de directeur de thèse et ensuite à titre de co-directeur (après son départ de l’Université Laval), qui m'a donné l'opportunité de travailler dans son laboratoire, et m'a confié un projet de recherche très stimulant. Je le remercie pour la qualité de son encadrement et sa grande disponibilité tout au long du projet.
Je remercie également mon directeur de thèse Jean Bousquet pour sa contribution à ce projet et pour ses précieux conseils.
Je remercie Jean Beaulieu pour sa collaboration et ses conseils qui ont été d'un grand apport pour la réussite de ce travail, ainsi que d’avoir accepté d’évaluer cette thèse.
Merci aussi à Martin Perron et Christophe Plomion d'avoir également accepté d'évaluer cette thèse.
Je tiens à remercier aussi tous les membres de l'équipe qui ont contribué de près ou de loin à la réalisation de ce travail, et en particulier Patrick Lenz, pour son aide en génétique quantitative, et Geneviève Parent pour les échanges de points de vue au plan scientifique qui m'ont été très bénéfiques.
Je tiens à remercier toute ma famille et mes amis qui m'ont toujours soutenu.
Avant-propos
Cette thèse comprend une introduction (chapitre 1), une conclusion générale (chapitre 4), ainsi que deux chapitres principaux de résultats originaux (chapitres 2 et 3) rédigés et présentés sous forme de manuscrits scientifiques dont un a déjà été publié.
Le chapitre 2 est publié sous la référence : Lamara M, Raherison E, Lenz P, Beaulieu J,
Bousquet J, MacKay J. 2016. Genetic architecture of wood properties based on association
analysis and co-expression networks in white spruce. New Phytologist 210: 240–255.
Le chapitre 3 prend la forme d’un manuscrit en préparation pour soumission sous la référence:
Lamara M, Parent G, Giguère I, Beaulieu J, Bousquet J, MacKay J. 2017. Association
genetics of acetophenone defence against spruce budworm in mature white spruce.
Dans les deux études dont je suis le premier auteur, j’ai réalisé la majorité des analyses pour l’obtention de résultats, leur interprétation et la rédaction de chaque chapitre sous la supervision de Jean Bousquet, directeur de thèse et de John MacKay, codirecteur de thèse. Les coauteurs ont contribué à la réalisation de certaines parties des expériences, notamment à l’acquisition de certaines données phénotypiques et la réalisation du design expérimental.
Chapitre 1. Introduction générale
1.1 Introduction
Un des enjeux contemporains de la gestion des forêts et de l’amélioration génétique des arbres forestiers est de développer des connaissances mieux intégrées des différentes caractéristiques d’intérêt économique dont la croissance, la qualité des bois et la résistance aux insectes et des aspects écologiques dont les caractères adaptatifs liés à la résilience aux stress abiotiques. Cette situation résulte notamment des changements climatiques et leurs effets tels que les perturbations abiotiques (ex. sècheresse) et un accroissement de certains stress biotiques (ex. dégâts causés par les insectes ravageurs et les maladies) affectant la productivité et le fonctionnement des forêts et des plantations d’arbres forestiers (Aitken et al., 2008; Netherer & Scholf, 2010). Une meilleure connaissance de la biologie des arbres et du fonctionnement des forêts permettrait de mieux décrire les phénomènes sous-jacents et de développer des solutions à ces enjeux. La présente thèse vise précisément à bonifier nos connaissances de base par l’étude de l’architecture génétique de différents caractères reliés à la productivité et la résilience chez l’épinette blanche.
L’étude de l'architecture génétique des caractères quantitatifs s’amorce par l'identification des loci impliqués dans la variabilité du caractère, leurs effets et leurs positions sur le génome (Mackay, 2001). Plusieurs études de ce type ont été réalisées chez diverses espèces forestières d’intérêt économique et ont visé notamment les propriétés du bois, la croissance, la résistance aux pathogènes et la tolérance à la sècheresse. Elles ont souligné que la plupart de ces caractères sont des caractères complexes sous le contrôle de plusieurs gènes à effet faible (Neale & Savolainen, 2004).
Le but de cette thèse est d’utiliser les ressources génétiques disponibles pour l’épinette blanche afin d’étudier les bases moléculaires de plusieurs caractères quantitatifs dont la qualité du bois, la croissance et la résistance à la tordeuse des bourgeons de l’épinette, et de mettre en évidence les gènes impliqués dans leur contrôle génétique.
Le premier chapitre de la thèse sera consacré à une revue bibliographique qui fait état des connaissances actuelles sur les caractères quantitatifs d’intérêt économique qui sont l’objet de l’amélioration génétique. Il dresse une synthèse de l’utilisation des marqueurs
moléculaires et des approches utilisées pour l’identification des loci contrôlant la variabilité génétique sous-jacente à ces caractères quantitatifs («Quantitative Trait Loci», QTL). Les hypothèses de recherche et les objectifs sont décrits à la fin du chapitre.
1.2 Les caractères quantitatifs et leur contrôle génétique
La recherche présentée dans cette thèse porte sur l’épinette blanche (Picea glauca [Moench] Voss), une espèce de la famille des Pinacées (Pinaceae). Il s’agit d’un des conifères les mieux étudiés au niveau génétique et génomique, ce qui facilite les analyses à la fine pointe des connaissances et fait en sorte que les conclusions seront d’intérêt pour les chercheurs travaillant sur plusieurs autres espèces d’arbres, notamment ceux visant les membres de la famille des Pinacées. Par surcroit, l’épinette blanche est l’une des espèces de conifère les plus largement distribuées dans la forêt boréale de l’Amérique du nord de l’Alaska au Labrador. Elle est fortement utilisée par l’industrie forestière étant donné la qualité de son bois et son bon taux de croissance (Beaulieu, 1994). Son bois a une large gamme d’utilisation et constitue une source importante de matière première pour la fabrication de pâte de haute qualité grâce aux longues fibres, critère recherché par l’industrie du papier. Le bois de l’épinette blanche est aussi utilisé dans le domaine de la construction pour la fabrication des planches, poutrelles et autres composants structuraux.
1.2.1 Phénologie et croissance
Au cours d’une saison de croissance, la phénologie est le déroulement et la séquence des
différents évènements biologiques marquant la croissance et le développement
morphologique de l’arbre (Schwartz 2003). D’importants évènements phénologiques sont le débourrement des bourgeons au printemps, la période de croissance active en été et l’aoûtement des bourgeons à la fin de l’été et l’endurcissement au froid durant l’automne; ces phases phénologiques sont toutes essentielles à la survie et la productivité des arbres (Delpech et al., 1985). Les caractères phénologiques sont parmi les paramètres les plus étudiés chez les arbres forestiers parce qu’ils constituent un élément clé pour déterminer le fonctionnement et l’adaptation des arbres à leur climat local. On distingue deux grands types d’adaptation : 1) l’adaptation aux conditions défavorables et 2) la capacité d’utilisation des ressources disponibles d’une manière efficace (Hänninen, 1990; Kramer et al., 2000). À titre d’exemple, les changements de température et de photopériode au cours
de l’année influencent grandement la phénologie dont notamment la date de débourrement et la date de formation des bourgeons apicaux, respectivement, et entraînent une compétition pour la lumière dans une population d’espèces mixtes (Kramer et al., 2000; Lechowicz, 2001). La phénologie influence aussi les interactions biotiques. À titre d’exemple, la phénologie du débourrement des bourgeons au printemps affecte directement les performances biologiques de la tordeuse des bourgeons de l'épinette (Fleming & Voney, 1995; Rauchfuss & Ziegler, 2011).
L’amélioration génétique des arbres forestiers a pour objectif majeur l’augmentation de la productivité (Grattapaglia et al., 2009). Plusieurs caractères d’adaptation sont liés à la
phénologie annuelle de la croissance chez les arbres et sont directement impliqués dans la réponse aux variations climatiques saisonnières. Dans ce contexte, l’étude de la variation génétique intraspécifique de la phénologie des caractères adaptatifs, tels que la croissance saisonnière, la date de débourrement et la date de formation des bourgeons apicaux est pertinente pour optimiser la survie, la croissance et la productivité (Badeck et al., 2004; Holliday et al., 2010; Pelgas et al., 2011). Plusieurs études ont montré que les caractères liés à la croissance et à la phénologie des bourgeons chez les arbres forestiers sont des caractères quantitatifs contrôlés par plusieurs gènes à effet faible et sont aussi fortement influencés par l’environnement (Jermstad et al., 2003; Rohde et al., 2011; Pelgas et al., 2011).
La phénologie influence aussi la formation et la structure du bois chez les arbres. Selon les différentes étapes du développement saisonnier de l’arbre, on peut distinguer plusieurs types de bois. Par exemple, le bois initial est formé au début de la saison de croissance (il est parfois appelé bois de printemps) tandis que le bois final est formé à la fin de la saison de croissance (il est parfois appelé bois d'été). La variation de la densité du bois dans un cerne annuel de croissance permet de distinguer ces deux types de bois et fournit également des informations sur les processus physiologiques liés à la formation du bois (Koubaa et al., 2002). Le nombre et la largeur des cernes annuels nous renseignent sur l'âge et les conditions de croissance de l’arbre respectivement. Chez les conifères, le bois final est composé de trachéides généralement plus longues, à faible diamètre, à faible angle d’inclinaison des microfibrilles (donc plus dense), à paroi cellulaire plus épaisse et à teneur plus élevée en cellulose (Uggla et al., 2001; Barnett & Jeronimidis, 2003). L’étude de
Rozenberg et al. (2002) a démontré que les fluctuations de la densité au sein d’un cerne du bois de l'épinette de Norvège (Picea abies [L.] Karst) entre le bois initial et bois final sont influencées par la réaction de l'arbre au déficit hydrique qui apparaît au cours de l’été. Chez les conifères, un cerne de croissance annuelle est principalement formé du bois initial (> 70%), ce qui fait que les propriétés du bois initial sont généralement représentatives du cerne complet et que le bois final est distinct des deux derniers (Beaulieu et al., 2011; Fernández et al., 2012). À l’échelle de plusieurs années de croissance, un autre type de variation concerne la transition de la formation du bois juvénile à la formation du bois mature. Une description détaillée de la variabilité phénotypique sur plusieurs cernes de croissance représente la base de l'analyse génétique et l'estimation des paramètres génétiques quantitatifs par rapport à l'âge de développement (Lenz et al., 2010). Chez l'épinette blanche, le nombre et les effets des gènes sous-jacents à ces caractères adaptatifs n'ont pas encore été caractérisés.
1.2.2 Qualité du bois
Les caractères de qualités du bois sont des critères de sélection à considérer dans le but de maintenir une bonne qualité des billes, des sciages et de la fibre de bois tout en visant un taux de croissance élevé (Beaulieu et al., 2009). Parmi les caractères d’intérêt, on retrouve notamment des propriétés physiques du bois (e.g. densité), des propriétés mécaniques (e.g. module d’élasticité, MOE), des propriétés anatomiques (e.g. angle des microfibrilles de cellulose, AMF) et la longueur des fibres.
La densité du bois est souvent considérée comme un caractère et un facteur clé pour la détermination de la qualité du bois (Zobel & van Buijtenen, 1989). Pour différents secteurs de l’industrie, la densité du bois est une propriété affectant le rendement et la valeur des produits car elle influence directement les propriétés mécaniques des planches obtenues par sciage et l’homogénéité du bois (Mäkinen et al., 2002; Raymond, 2002). La densité du bois influence aussi le rendement en pâte à papier à partir d’un volume de bois donné (Mäkinen et al., 2002). L’angle des microfibrilles de la cellulose de la paroi secondaire influence aussi la qualité du bois; un angle des microfibrilles élevé est associé à une moins grande rigidité. La variabilité des propriétés du bois a été étudiée chez plusieurs espèces dans différentes populations. Chez les épinettes (blanche et noire), diverses études ont montré
que la variation d’origine génétique influence significativement les caractères du bois tels la densité et la longueur des fibres et ont des impacts sur le rendement et la performance des produits du bois dans l'industrie du sciage et des pâtes et papiers (Beaulieu, 2003).
Contrairement aux caractères de croissance, plusieurs études ont montré que les niveaux d’héritabilité des caractères liés à la qualité du bois sont relativement élevés indiquant ainsi un contrôle génétique modéré à élevé, et un effet environnemental plutôt faible sauf en conditions extrêmes (Hamilton & Potts, 2008; Grans et al., 2009; Hallingback et al., 2010; Resende et al., 2017).
Chez l’épinette blanche, l’étude de Lenz et al. (2010) a montré que cette héritabilité varie de 0,28 à 0,34 pour l’AMF, de 0,27 à 0,41 pour le MOE et de 0,13 à 0,69 pour la densité du bois, qui sont des valeurs généralement plus élevées que l’héritabilité pour la croissance. Étant donné l’héritabilité et la variation phénotypique élevées de ces caractères, on peut s’attendre à ce que la sélection donne de bons gains génétiques, ce qui représente un intérêt important dans les programmes d’amélioration.
1.2.3 Résistance aux insectes et développements récents concernant la tordeuse des bourgeons de l’épinette
Tout au long de leur cycle de vie, les arbres sont confrontés à de nombreuses attaques par des insectes ravageurs (Alfaro & Fuentealba, 2016). Les dégâts causés par ces insectes conduisent à une diminution importante de la productivité des forêts (Pinkard et al., 2010; Fuentealba & Bauce, 2016). Par conséquent, les arbres ont développé divers mécanismes de résistance qui leur permettent de faire face, par exemple, à la défoliation par des insectes herbivores (Fuentealba & Bauce, 2016). On peut distinguer deux grandes catégories de mécanismes de défense: 1) la formation de barrières physiques comme la lignification des parois cellulaires, les cuticules cirées, l’écorce et les épines; 2) les mécanismes de défense chimique, avec des effets toxiques qui fournissent une barrière défensive efficace (Kovalchuk et al., 2013). Les composés de défense chimique (métabolites secondaires) peuvent être constitutifs (préformés), et ils sont donc toujours présents dans l’arbre en prévision d'une attaque par des insectes ou des réponses induites lors d'une attaque d'herbivores (Mithöfer & Boland, 2012). Les composés du métabolisme secondaire impliquent l’activation de différentes voies de biosynthèse telle que les phénylpropanoïdes
(composés phénoliques) et les terpénoïdes (oléorésine). Au cours des dernières décennies, des insectes ravageurs dont les scolytes (coléoptères) et la tordeuse des bourgeons de l’épinette (TBE, Choristoneura fumiferana [Clemens]) (lépidoptère) ont causé des pertes économiques importantes dans les forêts de conifères d'Amérique du Nord (Nowak et al., 2008; van Mantgem et al., 2009; Waring et al., 2009; Delvas et al., 2011). Bien que les mécanismes de résistance soient encore relativement peu élucidés, des études récentes ont montré que les oléorésines (composés terpéniques) constituent une barrière chimique contre les scolytes et les champignons (Franceschi et al., 2005; Kane & Kolb, 2010), alors que les composés phénoliques sont impliqués dans la résistance contre la TBE (Delvas et al., 2011).
La TBE est l’un des principaux insectes défoliateurs de la forêt boréale d'Amérique du Nord (Blais, 1983; Royama, 1984). Les épidémies de TBE peuvent se propager sur de très grandes superficies forestières et causer des dégâts économiques importants en réduisant la croissance et en causant de la mortalité ce qui diminue très considérablement la qualité du bois de l’hôte (Blais, 1983; Sanders, 1991; Rauchfuss & Ziegler, 2011). Dans l'Est du Canada, ces pertes sont estimées à 44 millions de m3 de bois par an lors des épidémies (Sterner & Davidson, 1982). Cette capacité de la TBE à défolier des millions d'hectares de forêt pendant une infestation (Rauchfuss & Ziegler, 2011) a mené à la réalisation d’études visant à découvrir quels seraient les mécanismes de résistance des arbres hôtes.
Le potentiel de résistance à la TBE a été décrit à la suite de la découverte de quelques épinettes blanches résistantes et l’étude comparative des composés chimiques accumulés dans les aiguilles (Daoust et al., 2009). L’étude de Delvas et al. (2011) a mis en évidence un mécanisme constitutif de résistance contre la TBE lié à l’accumulation de deux composés phénoliques spécifiques aux arbres résistants, soit le picéol et le pungénol. En revanche, les arbres non résistants contiennent principalement les formes glucosilées de ces composés phénoliques, soit la picéine et la pungénine. Récemment, l’étude de Mageroy et al. (2015) a montré qu’une enzyme de la famille des β-glucosidases agit sur les composés glycosylés pour libérer le picéol et pungénol dans les arbres résistants (Delvas et al., 2011). Ces composés ont un effet toxique sur la TBE puisqu’ils augmentent la mortalité des larves, retardent leur développement et réduisent la masse de la pupe (Delvas et al., 2011). Une meilleure compréhension du mécanisme moléculaire de résistance des arbres hôtes à la
TBE et l’identification des gènes qui confèrent cette résistance permettrait de proposer son origine d’un point de vue évolutif et de développer des méthodes facilitant la sélection des arbres plus résistants aux attaques de TBE.
1.3 Corrélations entre les caractères
Cette thèse présente des analyses visant à mieux comprendre les bases moléculaires de différentes caractéristiques des arbres forestiers dont la structure du bois, la résistance aux insectes et la croissance. La thèse vise par surcroît à développer des connaissances mieux intégrées de ces différents caractères car jusqu’à présent, ils ont été largement étudiés de façon indépendante. Plusieurs études s’appuyant sur des analyses phénotypiques ont mis en évidence des corrélations entre les caractères, qui justifient le besoin d’une meilleure intégration et qui justifient une présentation des connaissances actuelles.
D’abord, il est pertinent de connaître la nature de la corrélation génétique qui existe entre deux caractères pour anticiper quelle pourrait être leur base génétique. Cette connaissance permettra aussi de déterminer la réponse positive ou négative d’un caractère à la sélection génétique effectuée sur l’autre caractère. Deux types de corrélations sont généralement considérés dans les analyses de génétique quantitative, la corrélation génétique et la corrélation phénotypique.
D’un point de vue appliqué, plusieurs études basées sur les corrélations génétiques et phénotypiques ont montré qu’une sélection basée seulement sur des caractères de croissance à pour conséquence de diminuer considérablement la densité et la rigidité du bois chez les conifères (Ivkovich et al., 2002; Lenz et al., 2010, 2013; Kennedy et al., 2013). La densité est aussi fortement corrélée avec différents caractères mécaniques et anatomiques du bois (Zobel & Van Buijtenen, 1989). Chez l’épinette, l’angle des microfibrilles influence directement les caractéristiques mécaniques du bois et peut expliquer jusqu’à 70% de la variation du module d’élasticité MOE qui traduit la capacité du bois de résister à la flexion (rigidité) (Alteyrac et al., 2006). Chez l’eucalyptus, l’angle des microfibrilles et la densité expliquaient 96% de variation de la rigidité (Evans & Ilic, 2001). Pour la qualité de la pâte à papier, la composition chimique du bois telles que la teneur en cellulose, hémicellulose et lignines constitue aussi un caractère d’intérêt majeur (Raymond, 2002). Dans ce contexte, la sélection pour une meilleure adaptation ou un taux de
croissance élevé peut avoir un effet négatif sur plusieurs des caractéristiques du bois (Rozenberg & Cahalan, 1997)
L’étude de Lenz et al. (2010) a montré aussi que les propriétés du bois peuvent varier avec l’âge, donc les corrélations phénotypiques et génotypiques inter-cernes, elles-mêmes indicatrices des corrélations juvénile-adulte, qui sont à prendre en considération afin de déterminer l’âge optimal de sélection.
Les caractères de croissance et de la qualité du bois sont aussi influencés par les conditions environnementales. Par exemple, le déficit hydrique a un effet différentiel sur la densité de différents génotypes (Rozenberg et al., 2002), ce qui indique une variation génétique pour la plasticité phénotypique et une l'interaction entre le génotype et l'environnement (G x E). Ainsi, les interactions G x E doivent être prises en compte dans les analyses génétiques notamment pour prédire les effets des variations climatiques sur l’adaptation des espèces (Assmann, 2013; El-Soda et al., 2014; El-Soda et al., 2015; Iwata et al., 2016).
Les corrélations entre les caractères ne concernent pas seulement les caractères de croissance et de la qualité du bois mais aussi les caractères de résistance. Chez les plantes en général, le déploiement des mécanismes de défense constitutifs et induits contre les différents agresseurs améliore les chances de survie. Ces mécanismes de défense impliquent généralement des coûts énergétiques et s’ils sont assez importants, ils peuvent être au détriment de la croissance des plantes (Huot et al., 2014). Dans un contexte de résistance des arbres aux insectes herbivores, il pourrait y avoir des compromis au niveau de l'allocation des ressources énergétiques entre les caractères de croissance, de la qualité du bois et la biosynthèse des métabolites secondaires liés à la résistance. De tels compromis peuvent se concrétiser par des corrélations phénotypiques ou génétiques négatives entre l’accumulation constitutive de ces métabolites secondaires et la croissance (Sampedro, 2014).
1.4 Analyses des bases moléculaires des caractères quantitatifs
1.4.1 Objectifs de dissection génétique des caractères quantitatifsChez les arbres forestiers, l’analyse de l'architecture génétique des caractères quantitatifs constitue un objectif important non seulement pour l’amélioration génétique mais aussi
pour comprendre l’adaptation des espèces aux conditions climatiques et les différents processus évolutifs sous-jacents, et dans un dessein de mieux asseoir les stratégies de conservation de la diversité génétique.
1.4.1.1 Amélioration génétique des arbres forestiers
Les marqueurs moléculaires représentent des outils potentiellement très puissants pour faciliter la sélection génétique précoce (Neale & Savolainen, 2004; González-Martínez et al., 2007). Les temps de sélection chez les arbres forestiers peuvent varier 7 à plus de 30 ans, par exemple chez les conifères boréaux. De longues périodes de temps sont aussi requises parce que l’amélioration génétique s’effectue selon un cycle continu de sélection, allant de la conduite des croisements à l’évaluation au champ des arbres possédant les caractéristiques désirées. Au contraire, l’identification des marqueurs génétiques liés aux caractères désirables permettrait de sélectionner les arbres les plus performants dès le stade de semis sans attendre qu’ils parviennent à maturité. Ainsi, l’analyse des caractères quantitatifs à l’aide de ces marqueurs a été proposée dans le but de réduire les temps de sélection de plusieurs années. Les bénéfices escomptés peuvent être réalisés si l’obtention des données génotypiques permet d’évaluer correctement le potentiel des descendances.
L’amélioration génétique des arbres forestiers est basée sur l’exploitation de la grande variabilité génétique des arbres forestiers et vise la production d’une ressource de qualité s’appuyant sur la compréhension des mécanismes génétiques des caractères d’intérêt. La génétique quantitative classique permet de décrire leur contrôle génétique mais ne permet pas de déterminer les gènes impliqués, ni leurs effets et leurs interactions. Or, la connaissance des bases moléculaires des caractères d’intérêt pour l’amélioration génétique pourrait permettre la sélection directe des arbres, et ainsi accélérer le processus d’amélioration. Ainsi, le développement des marqueurs moléculaires performants permettrait d’accéder directement à la variabilité génétique présente au sein des populations. Il en résulterait une meilleure mise en valeur et conservation de la diversité génétique dans les populations naturelles qui est à la base de tous les programmes d’amélioration et de conservation des ressources génétiques.
1.4.1.2 Adaptation et évolution
Les arbres sont soumis aux variations des facteurs abiotiques ou biotiques au fil des saisons, créant des pressions de sélection qui entraînent une adaptation plus ou moins grande aux conditions historiques. La diversité génétique qui existe à l’intérieur des espèces d’arbre permet l’adaptation au fil des générations, qui se manifeste notamment par des changements du métabolisme et de la morphologie de l’arbre (El Kayal, 2011; St.Clair & Howe, 2011). Les études de génétique quantitative classique montrent que les caractères adaptatifs tels que la phénologie, l'acclimatation au froid, la résistance à la sècheresse et aux insectes sont héritables et les variations phénotypiques sont liées partiellement à des variations de l'information génétique au niveau de l’ADN. Le développement des technologies de séquençage et de génotypage à haut débit permet la mise en évidence d’un nombre élevé de marqueurs moléculaires (Prunier et al., 2016). Il en résulte la capacité d’analyser les bases génomiques de ces caractères au sein des populations d’arbres forestiers afin d’en déterminer leur potentiel génétique adaptatif et éventuellement les risques liés aux changements climatiques. Plusieurs études sur l'architecture génétique se sont intéressées aux caractères adaptatifs chez différentes espèces d'arbres dont les conifères (revue par Neale & Kremer, 2011). À titre d’exemple, de nombreux gènes ont été reliés à l’aoûtement qui est une caractéristique phénologique importante de l'acclimatation au froid chez les arbres (Frewen et al., 2000; Howe et al., 2003; Holliday et al., 2008) et d’autres ont établi un lien entre les polymorphismes de l’ADN et les variables climatiques (Prunier et al., 2011; Prunier et al., 2013; Hornoy et al., 2015).
1.4.1.3 Conservation de la diversité génétique
Une très grande diversité génétique a été produite par les forces évolutives au cours des 3,5 milliards d'années sur notre planète (Eisner et al., 1995) et la conservation génétique vise à préserver la diversité qui est présente dans les populations d’organismes vivants. Les développements de la génétique moléculaire contribuent significativement à la création de connaissances en biologie de la conservation depuis plusieurs années. À titre d’exemple, l'information génétique fournit une indication des pertes de la diversité génétique, ce qui pourrait réduire la capacité d'adaptation de l'espèce (Beardmore & Winder, 2011; Namroud et al., 2012). La possibilité d’examiner des milliers de marqueurs, aide à répondre à
plusieurs questions importantes sur la conservation de cette diversité au sein des populations (Allendorf et al., 2010).
Dans ce contexte, l’étude de l’architecture génétique identifiant les gènes et les variations intrinsèques influençant les caractères d’adaptation est utile pour la conservation des ressources génétiques forestières. À titre d’exemple, ces connaissances pourraient contribuer aux efforts de migration assistée comme stratégie d’adaptation pour les espèces vulnérables au risque d’extinction. La migration assistée consiste à effectuer un déplacement actif par l’homme d'espèces de leur aire de répartition indigène vers de nouvelles zones où elles devraient évoluer à mesure que le climat change ou encore, d’effectuer des déplacements de sources de graines à l’intérieur même de l’aire de répartition d’une espèce (Hunter, 2007; Schwartz et al., 2012; Hällfors et al., 2017). D’autres auteurs proposent des stratégies bien différentes reliées à la création d’organismes génétiquement modifiés; par exemple, Adams et al. (2002) ont suggéré que le transfert de gènes de résistance par génie génétique chez certaines espèces vulnérables pourrait prévenir la perte de certaines essences forestières importantes.
1.5 Méthodes d’analyses moléculaires des caractères quantitatifs
Diverses approches ont été utilisées pour identifier des marqueurs génétiques qui pourraient expliquer une partie importante de la variation phénotypique observée dans les caractères quantitatifs d'intérêt. Ces approches ont évolué de la détection des QTL impliqués dans la variabilité des caractères d’intérêt dans des pedigree, aux approches d’études d'associations et enfin, à la sélection génomique.
1.5.1 Approches classiques d’identification de QTL
Les analyses visant à identifier des QTL (Quantitative Trait Loci) sous-jacents ont été parmi les premières approches moléculaires utilisées pour l’analyse des bases moléculaires de la variation des caractères quantitatifs chez diverses espèces forestières (Tableau 1.1). L’identification des QTL se fait habituellement dans une population issue d’un ou plusieurs croisements contrôlés entre des parents, idéalement aux phénotypes contrastés pour le caractère à l’étude ou aux génotypes contrastés. Ces analyses ont identifié plusieurs QTL dans le contrôle des caractères quantitatifs et ont permis de déterminer le nombre de QTL, les effets de chaque QTL et leur position approximative sur le génome (Tableau 1.1).
Initialement, un des objectifs visé par le développement de la cartographie QTL était de fournir des outils pour la sélection assistée par marqueurs (SAM). Malgré les nombreuses études réalisés et les résultats encourageants, cette approche s’avère peu appropriée pour la SAM appliquée aux caractères quantitatifs chez les arbres forestiers en raison de différentes contraintes. Ces contraintes sont dues à 1) la résolution des QTL qui est généralement faible principalement parce que la zone du génome contenant un QTL est souvent très grande et contient un grand nombre de gènes potentiellement impliqués dans le contrôle génétique du caractère quantitatif et plusieurs autres qui n’auraient pas été cartographiés (Pelgas et al., 2011; Rohde et al., 2011; Lind et al., 2014); 2) les effets des QTL détectés sont souvent surestimés car les analyses de cartographie des QTL sont généralement effectuées avec un faible nombre de marqueurs dans des populations de faible taille, et les QTL détectés sont rarement validés dans une autre population (Beavis, 1998; Jannink et al., 2001; Pelgas et al., 2011). Cette approche est donc utile pour localiser des régions génomiques mais elle demeure peu puissante pour décrire avec précision l'architecture génétique des caractères quantitatifs complexes.
1.5.2 Les analyses d’association génétique 1.5.2.1 L’approche GWAS
Les études d’association constituent une approche alternative pour surmonter les contraintes intrinsèques des études traditionnelles de détection de QTL (Neale & Savolainen, 2004). Cette approche consiste à établir une corrélation entre une ou plusieurs variations alléliques et la variation phénotypique observée. Elle présente plusieurs caractéristiques distinctes par rapport aux études de QTL, notamment les études d’associations: 1) reposent sur la caractérisation d’une population dite ‘naturelle’ et ne nécessite pas une forte structure; 2) exploitent une plus grande diversité allélique; 3) offrent une plus grande résolution en raison de l’exploitation de tous les évènements de recombinaison historique des individus de la collection choisie, contrairement à la cartographie QTL qui est limitée par l'information de recombinaison provenant de la ségrégation biparentale; 4) reposent sur l’existence du déséquilibre de liaison (ou LD pour «linkage disequilibrium») entre les marqueurs et le QTL (Khan & Korban, 2012).
Chez les arbres forestiers, les populations ont généralement des grandes tailles effectives (des dizaines de milliers d’individus, Bouillé & Bousquet, 2005) et une faible structure de populations (Neale & Savolainen, 2004; González-Martínez et al., 2006), ce qui est favorable pour les analyses d’association. Toutefois, une approche d'association pan-génomique aussi connue sous le nom de («Genome Wide Associaton Study», GWAS), telle que utilisée chez l'humain (Hirschhorn & Daly, 2005) et certaines plantes (Atwell et al., 2010; Huang et al., 2010) présente des défis analytiques d’envergure pour la plupart des arbres forestiers. Ceci est dû au fait que les arbres forestiers possèdent également des caractéristiques qui réduisent le potentiel de succès de telles analyses, les plus importants étant, surtout pour les conifères, la grande taille de leur génome et la proportion élevé de séquences répétées, et la grande distance entre les gènes (Murray, 1998; Rigault et al., 2011), ainsi que le faible déséquilibre de liaison (linkage disequilibrium, LD) qui décroît rapidement en quelques centaines à quelques milliers de paires de bases, chez les populations naturelles (Namroud et al., 2010; Pavy et al., 2012). Pour contrer l’effet de faible LD, il faudrait utiliser une densité très élevée de marqueurs de l’ordre de plusieurs centaines de milliers et possiblement plus chez les espèces à faible LD comme chez les pins et les épinettes, afin de repêcher de façon efficace les liaisons entre marqueurs et QTN (quantitative trait nucleotide), ce qui entraînerait des coûts très élevés de génotypage. De plus, mises à part quelques espèces forestières modèles des genres Populus, Eucalyptus, Pinus, et Picea, les ressources génomiques pour ce faire sont peu disponibles. Ces contraintes sont donc très limitantes pour l’application à grande échelle des approches GWAS chez la vaste majorité des arbres forestiers malgré les avancées rapide des technologies de séquençage et de génotypage à haut débit (Prunier et al., 2016).
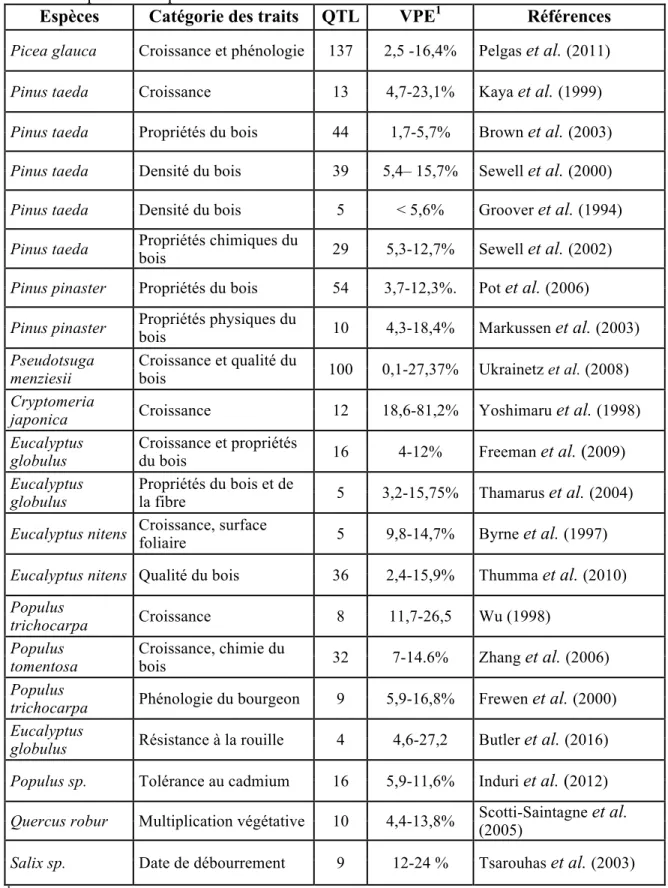
Tableau 1.1 Synthèse des résultats de plusieurs études de cartographie de QTL des
caractères quantitatifs par chez les arbres forestiers.
Espèces Catégorie des traits QTL VPE1 Références
Picea glauca Croissance et phénologie 137 2,5 -16,4% Pelgas et al. (2011) Pinus taeda Croissance 13 4,7-23,1% Kaya et al. (1999) Pinus taeda Propriétés du bois 44 1,7-5,7% Brown et al. (2003) Pinus taeda Densité du bois 39 5,4– 15,7% Sewell et al. (2000) Pinus taeda Densité du bois 5 < 5,6% Groover et al. (1994) Pinus taeda Propriétés chimiques du bois 29 5,3-12,7% Sewell et al. (2002) Pinus pinaster Propriétés du bois 54 3,7-12,3%. Pot et al. (2006)
Pinus pinaster Propriétés physiques du bois 10 4,3-18,4% Markussen et al. (2003) Pseudotsuga
menziesii Croissance et qualité du bois 100 0,1-27,37% Ukrainetz et al. (2008) Cryptomeria
japonica Croissance 12 18,6-81,2% Yoshimaru et al. (1998) Eucalyptus
globulus Croissance et propriétés du bois 16 4-12% Freeman et al. (2009) Eucalyptus
globulus Propriétés du bois et de la fibre 5 3,2-15,75% Thamarus et al. (2004) Eucalyptus nitens Croissance, surface foliaire 5 9,8-14,7% Byrne et al. (1997) Eucalyptus nitens Qualité du bois 36 2,4-15,9% Thumma et al. (2010) Populus
trichocarpa Croissance 8 11,7-26,5 Wu (1998) Populus
tomentosa Croissance, chimie du bois 32 7-14.6% Zhang et al. (2006) Populus
trichocarpa Phénologie du bourgeon 9 5,9-16,8% Frewen et al. (2000) Eucalyptus
globulus Résistance à la rouille 4 4,6-27,2 Butler et al. (2016) Populus sp. Tolérance au cadmium 16 5,9-11,6% Induri et al. (2012) Quercus robur Multiplication végétative 10 4,4-13,8% Scotti-Saintagne et al. (2005) Salix sp. Date de débourrement 9 12-24 % Tsarouhas et al. (2003) 1Variance phénotypique expliquée par les QTL détectés.
1.5.2.2 L’approche par gène candidat
En contrepartie, une approche basée sur l’analyse de gènes candidats a été proposée pour la réalisation des études d’association pour les caractères quantitatifs chez les arbres forestiers pour pallier aux limites liées à l’utilisation d’une approche pan-génomique (Neale & Savolainen, 2004). Cette approche a été décrite par Pflieger et al. (2001) et elle consiste à choisir les meilleurs gènes candidats possibles selon des critères indiquant leur implication potentielle dans le contrôle de caractères d’intérêts. De façon générale, les gènes candidats peuvent être choisis sur la base des critères suivants : 1) la fonction moléculaire démontrée par un impact sur le phénotype d’intérêt; à titre d’exemple, des gènes impliqués dans la biosynthèse de la lignine ou de la cellulose dans le cas des propriétés du bois (Vanholme et al., 2010; Joshi et al., 2011; Duval et al., 2014); 2) l’expression tel qu’indiquée par l’accumulation des transcrits d’ARN; à titre d’exemple, les gènes exprimés dans le feuillage pour des analyses de résistance contre les insectes herbivores (Mageroy et al., 2015) et les gènes fortement exprimés dans le xylème pour la qualité du bois (Paiva et al., 2008; Raherison et al., 2012, 2015); et 3) la position des gènes dans le génome, notamment à l’intérieur de l’intervalle de confiance des QTL détectés lorsque l’intervalle de confiance est restreint.
Les études d’association basées sur l’approche par gènes candidats ont généralement ciblé un nombre restreint de gènes, pour des raisons de manques d’information sur les gènes candidats potentiellement impliqués ou encore, pour diminuer la sévérité de la correction statistique pour le taux de découverte de faux-positifs (« False Discovery Rate », FDR) lorsque le nombre de gènes et de polymorphismes d’ADN testés est très grand. Elles ont néanmoins permis l’identification de certaines associations entre des variations dans ces gènes candidats et la variation phénotypique des caractères adaptatifs, de la qualité du bois, et autres chez plusieurs espèces d’arbres forestiers (Tableau 1.2). Cependant, la proportion de la variance phénotypique expliquée (VPE) par chaque marqueur significatif est très faible et varie de 0,01 à 7% (Table 1.2). Par exemple, chez le pin à encens (Pinus taeda L.), une forte association génétique a été détectée entre un SNP du gène α-tubulin, un gène impliqué dans la formation de microtubules corticaux, et la variation phénotypique de l'angle des microfibrilles du bois initial; toutefois, ce SNP n’expliquait que 3,6% de la variation phénotypique totale (González-Martínez et al., 2007). Pour les caractères
d’adaptation, une faible association a été détectée entre un SNP du gène dehydrin1 et l’efficience d'utilisation de l'eau, mesurée par la discrimination isotopique du carbone, et qui expliquait moins de 1% de la variation phénotypique (González-Martínez et al., 2008). Chez l’épinette blanche, une analyse basée sur 944 SNPs distribués au sein de 549 gènes candidats génotypés chez 492 arbres a identifié 13 SNPs associés aux différentes caractéristiques du bois et la VPE par chaque SNP variait entre 3 et 5% (Beaulieu et al., 2011).
1.5.2.3 Sélection génomique
Face aux différentes contraintes liées aux approches de cartographie des QTLs et les études d’association, la sélection génomique (SG) a été proposée comme approche alternative à la sélection assistée par marqueurs (SAM) (Meuwissen et al., 2001). Alors que la SAM consiste à identifier des associations entre marqueurs et caractères quantitatifs puis les utiliser dans un programme de sélection, la SG porte sur l’incorporation de toutes les informations provenant des marqueurs répartis sur l'ensemble du génome dans la prédiction des phénotypes sans avoir recours à des analyses d'association (Grattapaglia & Resende, 2011; Iwata et al., 2011). Dans l’approche SG, l’analyse se fait sur un très grand nombre d’individus (plusieurs centaines voire des milliers) qui constitue la population test. À partir des jeux de données génotypiques et phénotypiques, un modèle de prédiction est construit pour estimer la valeur génomique estimée, aussi connue sous le nom de «Genomic Estimated Breeding Value» (GEBV), des individus de la population. La précision de la prédiction de la valeur du GEBV est établie par le biais d’une validation réalisée sur une sous-population n’ayant pas servi au développement du modèle prédictif. Les résultats indiquent alors si il est possible de sélectionner efficacement à partir de l’information génotypique multi-locus sans avoir recours aux données phénotypiques (Heffner et al., 2009; Iwata et al., 2011; Lorenz et al., 2011).
La SG repose sur 4 paramètres : 1) l’étendue de la LD entre les marqueurs et les QTL, ce qui dépend de la taille effective de la population qui doit être petite (entre 20 et 50 parents croisés entre eux) ainsi que le nombre de marqueurs utilisés ou couverture génomique; 2) le nombre d’individus dans la population utilisée pour construire le modèle de prédiction; 3) l’héritabilité du caractère en question; 4) la distribution des effets des QTLs (Grattapaglia
& Resende, 2011). Cette approche est bien développée chez les animaux domestiques (Hayes et al., 2009; Hayes & Goddard, 2010). Elle est aussi efficace chez les plantes cultivées (Bernardo & Yu, 2007; Heffner et al., 2009; Jannink et al., 2010) et les plantes pérennes (Wong & Bernardo, 2008). Chez les arbres forestiers, l’utilisation de la SG pour l’amélioration des caractères d’intérêt est d’une grande utilité pour améliorer la précision de la sélection des caractères à faible héritabilité. En outre, la SG est très prometteuse pour raccourcir les cycles d’amélioration car elle permettrait de réduire le recours au phénotypage des descendants et les délais associés à ce phénotypage. Cet optimisme est appuyé par des résultats récents encourageants chez l’eucalyptus (Resende et al., 2012a); le pin à encens (Zapata-Valenzuela et al., 2012; Resende et al., 2012b), l'épinette blanche (Beaulieu et al., 2014a,b) et l’épinette noire (Picea mariana [Mill.] B.S.P.) (Lenz et al., 2017).
Toutefois, la contre-partie de la SG est que lorsque les modèles sont appliqués à des populations n’ayant aucune parenté avec la population test avec laquelle les modèles de prédiction ont été développés, les précisions obtenues de prédiction sont nulles (Beaulieu et al., 2014a,b; Lenz et al., 2017). La SG est donc une stratégie « sur mesure » de prédiction par la génomique qu’il faut refaire lorsqu’une nouvelle population cible se présente. Également, puisque la SG se base sur l’ensemble de l’information multi-locus des profils génomiques de façon simultanée, elle n’est pas adaptée à l’élucidation des causes génétiques moléculaires des variations phénotypiques observées. Dans de telles conditions, l’information génotypique à chaque locus n’est pas traitée de façon statistiquement indépendante.
1.6 Fonctions, rôles biologiques et expression des gènes identifiés par les
études d’associations
Dans le cadre d’études d’association par l’approche des gènes candidats, étudier l’expression des gènes en plus de tenir compte de la fonction prédite pourrait permettre la sélection de gènes candidats les plus pertinents pour un caractère d’intérêt; toutefois, aucune étude n’a testée cette hypothèse chez les arbres forestiers. Pour la majorité des gènes des conifères, la fonction est attribuée d’après la similarité de séquence à des gènes et des protéines d’organismes modèles qui ont été étudiés au niveau moléculaire et
biochimique. C’est pourquoi, les fonctions moléculaires sont généralement prédites et non démontrées. À titre d’exemple, des études ont permis de mettre en évidence certains gènes dans des voies de biosynthèses intrinsèques à la formation du bois chez les arbres (Plomion et al., 2001) dont les voies de biosynthèse des lignines (Boerjan et al., 2003) et de la cellulose (Doblin et al., 2002). Ces études ont montré que les processus cellulaires de la formation du bois mettent en jeu des centaines de gènes impliqués à chaque étape. Ceci n’est pas surprenant puisque la formation du bois se déroule suivant au moins cinq étapes : 1) division cellulaire, 2) élongation, 3) mise en place d’une paroi secondaire, 4) mort cellulaire programmée, 5) formation du bois de cœur (cette dernière étape se produit quelques années après les étapes 1 à 4) (Plomion et al., 2001).
Certaines études d’association sur le bois ont montré l’influence d’un ou de quelques gènes impliqués notamment dans la formation de la paroi cellulaire (Beaulieu et al., 2011); toutefois, aucune étude préalable du bois n’a entrepris une analyse systématique d’un grand nombre de gènes tel que présenté dans le chapitre 2 de la présente thèse. L’analyse qui y est présentée est l’une des premières visant à explorer la relation entre la diversité génétique affectant le phénotype, l’expression et les fonctions prédites des gènes. Des études d’expression basées sur la comparaison des profils des transcrits d’ARN dans plusieurs tissus ont montré qu’il existe une grande variété de profils allant de l’expression ubiquitaire à l’expression très préférentielle à un ou des tissus particuliers, parfois au sein d’une même famille de gènes (Raherison et al., 2012). Au niveau de la formation du bois, Pavy et al. (2008) ont mis en évidence un groupe de 360 gènes spécifiques au xylème en croissance chez l’épinette blanche, dont plusieurs étaient aussi spécifiques au xylème chez le peuplier et Arabidopsis thaliana. L’étude de Raherison et al. (2012) montre que certains gènes sont exprimés préférentiellement dans le bois juvénile comparé au bois mature et vice versa. Chez le pin maritime (Pinus pinaster Ait.), l’expression des gènes a aussi été étudiée en comparant le xylème à différents stades physiologiques et développementaux (Paiva et al., 2008).
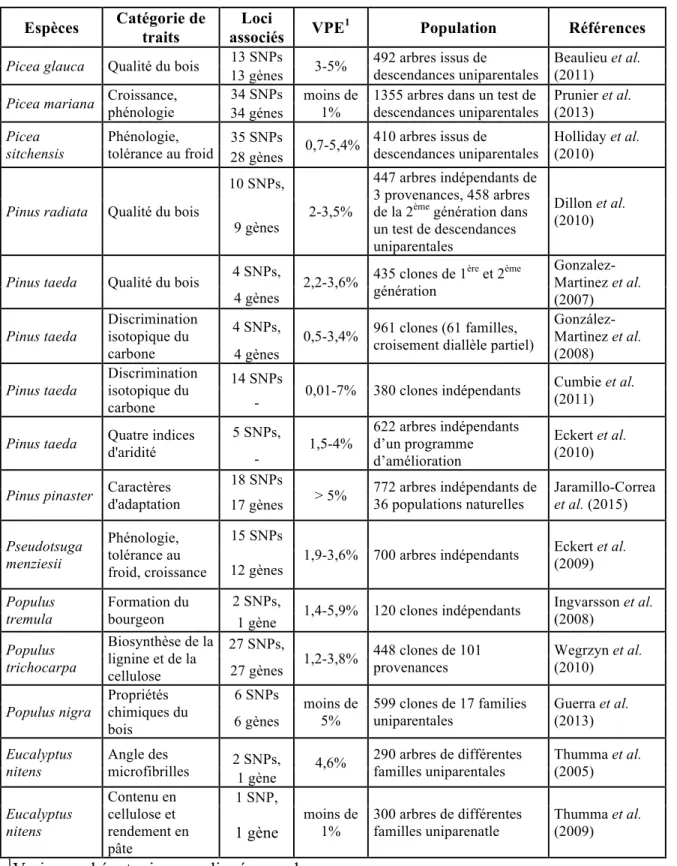
Tableau 1.2 Synthèse des résultats de quelques études d'association par l’approche de
gènes candidats chez les arbres forestiers.
Espèces Catégorie de traits associés Loci VPE1 Population Références
Picea glauca Qualité du bois 13 SNPs 3-5% 492 arbres issus de descendances uniparentales Beaulieu et al. (2011) 13 gènes
Picea mariana Croissance, phénologie
34 SNPs moins de 1%
1355 arbres dans un test de descendances uniparentales Prunier et al. (2013) 34 génes Picea sitchensis Phénologie,
tolérance au froid 35 SNPs 0,7-5,4% 410 arbres issus de descendances uniparentales
Holliday et al. (2010) 28 gènes
Pinus radiata Qualité du bois
10 SNPs,
2-3,5%
447 arbres indépendants de 3 provenances, 458 arbres de la 2ème génération dans un test de descendances uniparentales
Dillon et al. (2010) 9 gènes
Pinus taeda Qualité du bois 4 SNPs, 2,2-3,6% 435 clones de 1génération ère et 2ème
Gonzalez-Martinez et al. (2007) 4 gènes Pinus taeda Discrimination isotopique du carbone 4 SNPs,
0,5-3,4% 961 clones (61 familles, croisement diallèle partiel)
González-Martìnez et al. (2008) 4 gènes Pinus taeda Discrimination isotopique du carbone 14 SNPs
0,01-7% 380 clones indépendants Cumbie et al. (2011) -
Pinus taeda Quatre indices d'aridité 5 SNPs, 1,5-4% 622 arbres indépendants d’un programme d’amélioration Eckert et al. (2010) -
Pinus pinaster Caractères d'adaptation 18 SNPs > 5% 772 arbres indépendants de 36 populations naturelles Jaramillo-Correa et al. (2015) 17 gènes Pseudotsuga menziesii Phénologie, tolérance au froid, croissance 15 SNPs
1,9-3,6% 700 arbres indépendants Eckert et al. (2009) 12 gènes Populus tremula Formation du bourgeon 2 SNPs,
1,4-5,9% 120 clones indépendants Ingvarsson et al. (2008) 1 gène Populus trichocarpa Biosynthèse de la lignine et de la cellulose 27 SNPs,
1,2-3,8% 448 clones de 101 provenances Wegrzyn et al. (2010) 27 gènes Populus nigra Propriétés chimiques du bois 6 SNPs moins de 5% 599 clones de 17 families uniparentales Guerra et al. (2013) 6 gènes Eucalyptus nitens Angle des microfibrilles 2 SNPs, 4,6% 290 arbres de différentes familles uniparentales Thumma et al. (2005) 1 gène Eucalyptus nitens Contenu en cellulose et rendement en pâte 1 SNP, moins de 1% 300 arbres de différentes familles uniparenatle Thumma et al. (2009) 1 gène