Modélisation des effets de la configuration spatiale des
interventions de coupes sur la mortalité du caribou des
bois en forêt boréale
Mémoire
Virginie Vanlandeghem
Maîtrise en biologie - avec mémoire
Maître ès sciences (M. Sc.)
Modélisation des effets de la configuration spatiale des
interventions de coupes sur la mortalité du caribou des bois
en forêt boréale
Mémoire
Maîtrise en biologie
Maître ès sciences (M.Sc.)
Virginie Vanlandeghem
Sous la direction de :
Daniel Fortin, directeur de recherche
Pierre Drapeau, codirecteur de recherche
Résumé
Ce projet vise à évaluer l’effet de la configuration spatiale des coupes forestières et des routes sur la mortalité des adultes du caribou des bois (Ranfiger tarandus), écotype forestier, dans des paysages habités par le loup gris (Canis lupus) et une proie alternative, l’orignal (Alces alces). Nous avons utilisé un modèle centré sur l’individu pour modéliser les déplacements d’agents, représentant des individus fictifs de chaque espèce, dans des environnements virtuels. Les agents se déplaçaient selon des règles de déplacement établies à partir de l’analyse des données de suivis télémétriques de caribous, d’orignaux et de loups. Les environnements virtuels étaient façonnés par différents aménagements forestiers se distinguant par leur niveau de perturbations et par la configuration spatiale des coupes et du réseau routier. Le nombre de caribous morts par la prédation augmentait avec le niveau de perturbations des scénarios d’aménagement forestier. De plus, pour un même niveau de perturbations, la mortalité des caribous différait selon la configuration spatiale des activités forestières. Le risque de prédation des caribous était plus élevé lorsque la configuration des coupes dans le paysage engendrait un dense et vaste réseau routier et une fragmentation importante de l’habitat du caribou. Ainsi, en plus du niveau de perturbations, les plans de gestion de l’habitat de l’écotype forestier du caribou des bois devraient prendre en compte l’effet de la configuration spatiale des coupes et des routes sur la mortalité du caribou afin d’atténuer l’impact de la prédation sur ces populations dû à l’extraction des ressources.
Abstract
The aim of this project was to assess the impact of the spatial configuration of cuts and roads on adult mortality of the boreal ecotype of woodland caribou (Ranfiger tarandus) in landscapes inhabited by the gray wolf (Canis lupus) and an alternative prey, the moose (Alces alces). We used an individual-based model to simulate movements of agents, which are virtual individuals of each species, moving in virtual landscapes. Agents moved according to movement rules determined from field studies conducted on radio-collared caribou, moose and wolves. Virtual landscapes were shaped by various forest managements, differing in disturbance levels and spatial configuration of cuts and roads. The number of caribou killed by wolves increased with increasing disturbance levels. Also, for a given level of disturbance, caribou mortality varied with the spatial distribution of forest activities. Predation risk of caribou was higher in landscapes with extensive road networks and high degree of forest fragmentation. Thus, habitat management plans for boreal caribou should not only focus on assessing habitat disturbance thresholds but also consider the effect of the spatial configuration of cuts and roads on caribou mortality in order to reduce the impact of predation due to natural resource extraction on the ecotype’s populations.
Table des matières
RÉSUMÉ ...II ABSTRACT ... III TABLE DES MATIÈRES ... IV LISTE DES FIGURES ... VI LISTES DES TABLEAUX ... VII REMERCIEMENTS ... IX AVANT-PROPOS ... XI
INTRODUCTION ... 1
Les modèles centrés sur l’individu ... 5
Le caribou forestier comme modèle d’étude... 6
L’influence des activités humaines sur le caribou des bois ... 8
Le système caribou-loup-orignal ... 9
L’aménagement de l’habitat du caribou des bois jusqu’à maintenant ... 9
Objectif de l’étude ... 10
Approche empirique ... 11
CHAPITRE PRINCIPAL. MANAGEMENT-MEDIATED PREDATION RATE IN THE CARIBOU-MOOSE-WOLF SYSTEM: SPATIAL DESIGN OF LOGGING ACTIVITIES MATTERS... 13
Résumé... 14
Abstract ... 16
Introduction ... 17
Materials and methods ... 19
Forest management scenarios ... 19
Movement rules derived from radio-tracking caribou, moose and wolves ... 23
Statistical analysis of empirical movement rules ... 25
Individual-based model ... 26
Analysis of IBM’s outputs ... 29
Results ... 30
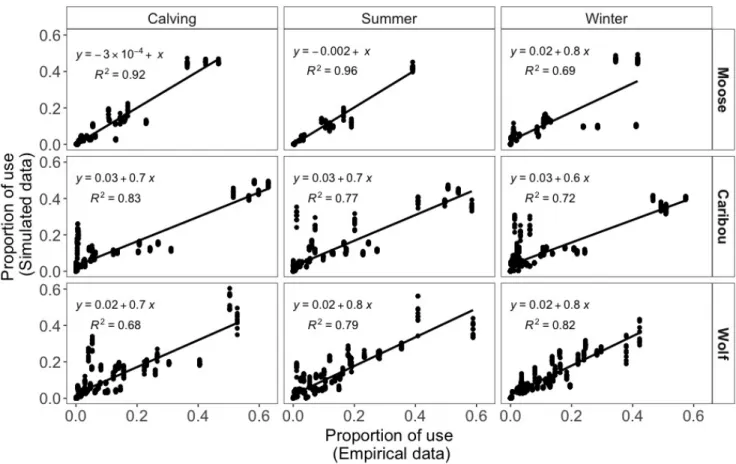
Movements rules from radio-collared boreal caribou, moose, wolves ... 30
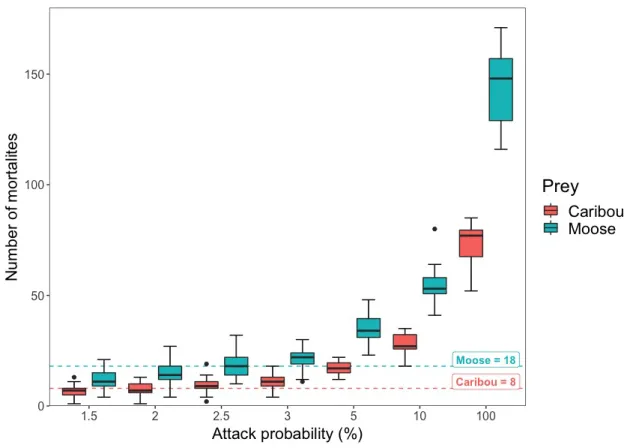
Wolf predation on caribou and moose under different management scenarios ... 33 Discussion ... 35 Acknowledgements ... 39 References ... 40 CONCLUSION ... 46 Perspectives ... 51 BIBLIOGRAPHIE GÉNÉRALE ... 53
ANNEXE 1. DATA PARTITIONING ... 61
ANNEXE 2. CALIBRATION OF THE PROBABILITY THAT WOLVES ATTACK A DETECTED PREY ... 61
ANNEXE 3. VALIDATION OF INDIVIDUAL-LEVEL BEHAVIORS OF AGENTS .... 63
Liste des figures
Figure 1. Representation of the landscape (5065 km2) in which forest management
scenarios were simulated for assessing the influence of the spatial configuration of cuts and roads on caribou mortality. Enlargements (404 km2) show the spatial
distribution of cuts and roads according to the three management strategies: a. mosaic, b. small agglomeration cutting and c. large agglomeration cutting. Aggregation of cuts directly affects the total area disturbed (without the 500-m buffer: mosaic = 62.4 km2, small agglomeration = 60.0 km2, large agglomeration =
57.5 km2; with the 500-m buffer: mosaic = 256 km2, small agglomeration = 155
km2, large agglomeration= 147 km2). Other colors represent land cover types: red =
burned area, blue = water body, dark green = closed-canopy conifer forest, green = open conifer forest without lichen, light green = open conifer with lichen, orange = mixed and deciduous and purple = other. ... 22 Figure 2. Relationship between road (a.), edge densities (i.e., perimeter of 500-m buffer around cuts and roads) (b.) and proportion of cuts (c.), as a function of the proportion of total disturbance (burned areas and cuts and roads with 500-m buffer) for each of the three forest management strategies (i.e., mosaic, small-agglomeration and large-small-agglomeration cutting). Overall relationships are represented by the dashed line. ... 23 Figure 3. General scheduling of the processes called during the execution of the
individual-based model. Agents move one by one according to specific movement rules. For wolves, the mode in which they are will influence the buffer radius within random points are drawn, but their movement rule remaining unchanged. Once all agents have moved, wolves in hunting mode can move to prey’s location (i.e., they can move twice) and kill a prey depending on the success of the three stages (i.e., detection, attack and kill). The cycle then starts over until the maximum number of steps is reached (i.e., 7 320 steps). ... 27 Figure 4. Number of caribou and moose agents killed by wolves during calving, summer and winter (left to right, respectively) as a function of the proportion of total disturbance (i.e., burnt areas, cuts and roads with 500-m buffer) in virtual landscapes harvested either by mosaic (blue), small- (yellow) or large-agglomeration (grey) cutting. Predictions of mortality number (represented by lines) were based on model coefficients (Table 2) with increasing level of disturbance. Average mortalities (represented by points) and their standard errors of simulations (n = 10) are also represented for each species and spatial configuration of cuts. The caribou population included 95 individuals at the beginning of all simulations, whereas the initial number of moose increased from 200 to 250 individuals with increasing level of disturbance... 34
Listes des tableaux
Table 1. Parameter estimates (b) and their robust standard errors (SE) estimated from step selection function in boreal caribou, moose, and wolves, for three biological seasons. The !" of k-fold cross validation is also given for each of the model... 32 Table 2. Coefficients (b ) and robust standard errors (SE) of generalised models relating the number of prey (i.e., caribou and moose) killed by wolves, 1) as a function of the proportion of disturbance in the virtual landscape and its interaction with categorical covariates representing forest management strategies (small-agglomeration cutting was the reference category), or 2) as a function of the proportion of disturbance and residual values of road and edge densities. For each species and each season, the analysis was fitted based on 130 simulations (10 replicates × 13 scenarios). Akaike’s Information Criterion (AIC) is provided to assess the relative support of the two models. ... 35
«When the snows fall and the white winds blow, the lone wolf dies but the pack survives»
Remerciements
Je tiens à remercier toutes les personnes qui ont été impliquées de près ou de loin dans ce projet et à sa réussite. Je n’aurais pu achever tout ce travail sans l’appui et le soutien de vous tous.
Tout d’abord, je voudrais remercier mon directeur de recherche Daniel Fortin qui m’a donné l’opportunité de prendre part à ce projet. Tu as toujours été là pour m’aider dans ce projet complexe, m’orienter lorsque je déviais du bon chemin et m’encourager à travers les épreuves rencontrées. Je suis reconnaissante d’avoir pu faire partie de cette aventure et de contribuer à l’apport d’informations pour la conservation du caribou forestier. J’aimerais aussi te remercier de m’avoir offert la possibilité d’être l’auxiliaire d’enseignement dans le cadre du cours d’Écologie des populations animales.
Je remercie également Pierre Drapeau, mon codirecteur. J’ai beaucoup apprécié les discussions que nous avons eues, tes encouragements et ton soutien m’ont été d’une grande aide. Merci aussi de prendre le temps de commenter et corriger de mes travaux.
J’aimerais remercier Jean-Pierre Tremblay d’avoir accepté de faire partie de mon comité d’encadrement et du jury de mon mémoire. Tes commentaires et tes idées sur le proposé et suite à mon séminaire de présentation m’ont permis de me questionner davantage sur mon sujet. Merci également à Yann Boulanger pour ses commentaires suite à l’évaluation de mon mémoire.
Un gros merci aux collaborateurs qui ont pris part à ce projet. Martin Côté et Frédéric Bujold sans qui les scénarios d’aménagement forestier n’auraient pu se faire. Je vous remercie du temps que vous avez pris pour les rencontres et la réalisation des scénarios. Je tiens également à remercier Jérôme Rioux pour les idées concernant l’élaboration des différentes configurations spatiales des coupes. Pierre Racine pour la construction des chemins forestiers des scénarios, tes idées et ton aide ont grandement été appréciées. Merci également à Maxime Boissonneault de Calcul Québec qui a répondu à toutes mes questions concernant le modèle centré sur l’individu et qui a pris le temps de
faire les modifications nécessaires aux codes afin que je puisse le paramétrer selon mes besoins.
Mon expérience à la maîtrise n’aurait pas été la même sans les conseils et les moments partagés avec mes collègues de laboratoire. Je remercie Florian Barnier pour les premiers échanges sur l’IBM et son fonctionnement, Marie-Caroline Prima pour toute l’aide apportée au commencement de mon projet avec l’analyse des données et pour se familiariser avec R, Ricardo Simon qui m’a aidé à surmonter les problèmes reliés à l’IBM et grâce à qui je me sentais moins seule dans ce tourbillon de programmation. Finalement à Guillemette Labadie et à Alexandre Terrigeol, mes compagnons de bureau, sur qui je peux toujours compter pour m’encourager, me divertir et me servir une tasse de thé dans les moments de stress. Merci également à tous mes collègues pour vos commentaires et votre appui dans mes travaux de rédaction, j’en suis énormément reconnaissante.
J’aimerais également remercier mes amis de biologie avec lesquels j’ai pu partager mon expérience de maîtrise. Merci aussi à tous mes coéquipiers et coéquipières au soccer, au CrossFit et au ultimate frisbee avec qui j’ai pu libérer mon corps de son énergie et du stress après de longues journées passées devant l’ordinateur.
Je dois remercier ma famille, mes parents, mon frère, mes sœurs et ma cousine, qui ont toujours été là pour me soutenir et m’encourager. Cela n’a pas toujours été facile, mais avec votre appui, j’ai pu trouver la force nécessaire dans les moments difficiles pour accomplir ce que j’avais entrepris.
Finalement, je remercie la personne qui me côtoie tous les jours et qui est toujours prête à m’écouter, mon copain Mathieu. Pendant ces trois ans d’étude, tu as été mon pilier à travers les épreuves. Je ne pourrai te remercier suffisamment d’avoir été là, de me faire rire et de me permettre de m’accomplir tous les jours. Maintenant, c’est le temps de relaxer et de profiter du soleil.
Avant-propos
Ce mémoire de maîtrise vise à évaluer l’effet de la configuration spatiale des coupes et des routes sur la mortalité des adultes du caribou forestier. Ce mémoire contient trois parties : une introduction générale, un chapitre présenté sous la forme d’un article scientifique rédigé en anglais et une conclusion générale. En tant que première responsable du projet et contributrice (développement des objectifs, analyses des données, réalisation des simulations, écriture du mémoire et de l’article), je serai première auteure de l’article. Les chercheurs qui sont coauteurs de l’article sont :
- Daniel Fortin, directeur de maîtrise et professeur au département de biologie de l’Université Laval. Il a contribué à l’élaboration du projet, au développement de la méthode statistique, des scénarios d’aménagement forestier et du modèle statistique et à la rédaction et à la révision de l’article.
- Pierre Drapeau, codirecteur de maîtrise et professeur au département des sciences biologiques de l’Université du Québec à Montréal. Il a contribué à l’élaboration du projet, au développement des scénarios d’aménagement forestier et la rédaction et la révision de l’article.
- Marie-Caroline Prima, docteure en biologie. Elle a contribué aux analyses statiques et à la révision de l’article.
- Martin-Hugues St-Laurent, professeur titulaire en écologie animale au département de biologie, chimie et géographie à l’Université du Québec à Rimouski. Il a contribué au développement des scénarios d’aménagement forestier et à la révision de l’article.
Le chapitre principal servira de base à un article qui sera éventuellement publié dans une revue scientifique internationale en anglais.
Introduction
La perte d’habitat est l’une des principales causes du déclin des populations et de l’extinction des espèces (Myers et al. 2000; Fahrig 2003). L’expansion des activités humaines, telles que l’extraction de ressources naturelles et la conversion de l’habitat naturel en terres agricoles, ont grandement contribué à la perte d’habitat nécessaire à la survie de certaines espèces (Saunders et al. 1991; Harrison et Bruna 1999). Par exemple, la récolte du bois dans les vieilles forêts de l’ouest des États-Unis est l’un des principaux facteurs ayant causé le déclin historique des populations de la chouette tachetée du Nord (Strix occidentalis caurina; Noon et Blakesley 2006). Également, la conversion des milieux naturels en terres agricoles ou en plantations de palmiers à l’huile sur l’île de Sumatra a grandement contribué à la réduction de l’habitat profitable aux tigres de Sumatra (Panthera
tigris sumatrae) et à ses proies contribuant ainsi à son déclin (Wibisono et Pusparini 2010;
Imron et al. 2011). Un défi majeur en biologie de la conservation consiste donc à concilier le maintien des espèces animales dans un environnement sujet à des perturbations anthropiques multiples, continues et largement réparties dans le paysage (Margules et Pressey 2000; Kareiva et Marvier 2015). Cela est particulièrement critique dans le cas des espèces désignées menacées, dont les plans de gestion reposent principalement sur l’identification et la désignation des habitats essentiels, en lien avec leur quantité, leur structure (c.-à-d., configuration spatiale et connectivité) et leur composition, dans le but de promouvoir la viabilité des populations. De surcroît, les perturbations anthropiques induisent des changements de l’environnement qui influencent la façon dont les individus utilisent l’espace (Koper et al. 2009; Garmendia et al. 2013; Beauchesne et al. 2014) et s’y déplacent (Johnson et al. 1992; Fahrig 2007; Beauchesne et al. 2013; Dickie et al. 2017). Ces modifications du comportement de déplacement des individus peuvent affecter directement leur survie et, ultimement, la dynamique des populations (Fahrig 2007). Il importe donc d’évaluer les impacts des perturbations anthropiques sur les individus afin d’anticiper les changements futurs de leur utilisation de l’environnement et d’établir des plans de gestions efficaces pour en atténuer les effets sur les populations (Fahrig 2001; Doherty et Driscoll 2018).
Face aux changements de leur environnement, les individus peuvent modifier leur déplacement et leur utilisation des milieux propices à leur survie. Par exemple, suite à la perte d’habitat causée par la coupe forestière, et donc à la diminution des ressources disponibles pour les espèces associées à ces forêts, certains individus agrandissent leur domaine vital et augmentent leurs déplacements entre les parcelles de ressource (Fisher et Wilkinson 2005; Faille et al. 2010) pour subvenir à leurs besoins. De plus, les perturbations anthropiques peuvent engendrer la perte fonctionnelle de l’habitat, via la fragmentation du paysage, en modifiant certaines caractéristiques des parcelles. En effet, les changements de la taille, de la connectivité avec les autres parcelles d’habitat propice à une espèce ainsi que la quantité de bordures avec des habitats non propices peuvent mener à l’inutilisation d’une parcelle d’habitat par les individus d’une espèce, et ce, malgré que cette dernière représente un habitat essentiel (Fahrig 2003). La matrice environnante peut aussi influencer l’utilisation des parcelles (Garmendia et al. 2013; Lesmerises et al. 2013), notamment par l’effet de bordure (Saunders et al. 1991; Fahrig 2003). La juxtaposition des coupes forestières ou des terres agricoles et des milieux naturels crée souvent une bordure franche, c’est-à-dire une transition abrupte, verticale et linéaire entre deux milieux (Matlack et Litvaitis 1999). Ces bordures peuvent modifier de façon importante la répartition, l’abondance et le comportement des individus (Murcia 1995). Citons en exemple le cas du déclin des populations de Pipit de Sprague (Anthus spragueii), un passereau nicheur des prairies en Alberta, chez qui l’effet de bordure a considérablement amplifié les effets négatifs de la perte d’habitat sur la population. En effet, les oiseaux évitaient, non seulement les terres cultivées, mais également les habitats de bordure des parcelles de prairies adjacentes; la réduction de l’habitat disponible par l’effet de bordure fut donc beaucoup plus importante que celle attribuée seulement à la conversion de l’habitat en terres agricoles (Koper et al. 2009). Pour toutes ces raisons, les perturbations anthropiques affectent à la fois directement et indirectement l’utilisation de l’espace par les individus. Ainsi, dans une optique de conservation, la quantité et la qualité de l’habitat doivent être prises en compte. Cependant, l’effet de la configuration spatiale de l’environnement (c.-à-d., la répartition des perturbations anthropiques et du milieu à préserver dans l’environnement) doit également faire partie intégrante des plans de gestion (Fahrig 2001; Nabe-Nielsen et al. 2010).
Par ailleurs, de plus en plus d’études démontrent qu’un changement des interactions trophiques peut causer le déclin des populations animales (Davis 2003; Sinclair et Byrom 2006; Bryant et Page 2005; DeCesare et al. 2010). Les perturbations anthropiques peuvent modifier ces interactions trophiques au sein d’un écosystème (Schneider 2001; Swihart et
al. 2001; Ryall et Fahrig 2006), puisque, notamment, les espèces impliquées ne réagissent
pas de la même façon face aux modifications de leur environnement (Kareiva 1987; Ryall et Fahrig 2006; Fortin et al. 2015; DeMars et Boutin 2018). Par exemple, la conversion des milieux naturels en terres agricoles peut modifier la communauté d’espèces de cet environnement. D’un côté, il y a les espèces qui perdent une partie de leur habitat (Koper et
al. 2009), et de l’autre, celles qui colonisent et exploitent le nouveau milieu (Umetsu et
Pardini 2007). De ce fait, la conversion du milieu peut être avantageuse pour les prédateurs généralistes (c.-à-d., se nourrissant de diverses proies) puisqu’ils peuvent exploiter davantage de ressources (Swihart et al. 2001; Gehring et Swihart 2003), autant les proies déjà présentes que les nouvelles. Ce changement de la communauté des prédateurs et de l’assemblage de proies dans l’environnement, ainsi perturbé, entraînerait donc en une intensification de la prédation dans le paysage pour certaines des espèces (Chalfoun et al. 2002; Batary et Balbi 2004; Okada et al. 2019). De plus, l’augmentation de la prédation d’une espèce peut résulter de l’interaction indirecte avec une autre espèce de proie chassée par le même prédateur (DeCesare et al. 2010). En effet, deux espèces sont en compétition apparente lorsque la présence et l’abondance de l’une des deux proies affectent négativement la population de la seconde, en induisant une réponse numérique du prédateur qui, en retour, intensifie la pression de prédation (Holt 1977). Il existe plusieurs exemples de ce type d’interaction impliquant une espèce menacée, et dont les changements de l’environnement induits par les humains ont augmenté les ressources disponibles pour la proie principale du prédateur, amplifiant ainsi la prédation et menant au déclin de l’espèce menacée (DeCesare et al. 2010). Par exemple, Bryant et Page (2005) ont démontré que la compétition apparente entre la marmotte de l’île de Vancouver (Marmota vancouverensis), une espèce en voie d’extinction, et le cerf à queue noir (Odocoileus hemionus hemionus) était influencée par la coupe forestière. La régénération des coupes a offert de nouvelles ressources pour le cerf, induisant une augmentation de sa population et celles de ses prédateurs, le loup (Canis lupus L.) et le cougar (Puma concolor), avec l’effet inverse sur la
marmotte. Par contre, les relations prédateur-proie sont également influencées par l’hétérogénéité spatiale de l’environnement (Kauffman et al. 2007; Gorini et al. 2012); la coexistence des espèces est même parfois possible grâce à un environnement hétérogène. En effet, certaines caractéristiques de l’environnement favorisent le succès du prédateur tandis que d’autres fournissent des refuges pour les proies (Huffaker 1958; Holt 1984; Kareiva 1987; Schneider 2001). La présence des refuges introduit une différence entre la densité totale de proies et la densité réellement disponible pour le prédateur (Gorini et al. 2012). Dans un système avec compétition apparente, le risque encouru par la proie dépendra à la fois de sa répartition spatiale, de celle de la proie alternative et des déplacements du prédateur (Fortin et al. 2015).
En somme, les répercussions des perturbations anthropiques sur les populations des espèces menacées sont multiples et influencent autant les ressources disponibles que les interactions trophiques. Dans ce contexte, l’organisation spatiale de l’environnement joue un rôle déterminant sur l’utilisation de l’espace des individus, sur le risque de prédation qui en résulte, et donc sur la dynamique des populations de la proie. Pour anticiper les conséquences des changements environnementaux et en minimiser les impacts sur les populations, il importe de se doter de moyens pouvant prévoir comment le système écologique spécifique sera affecté (Evans 2012). Or, les modèles traditionnellement utilisés à cette fin sont limités quant aux prévisions possibles des effets d’un nouvel environnement, car les taux démographiques estimés sont valides uniquement dans les conditions environnementales sous lesquelles ils ont été observés (Evans 2012). Pour pallier cette lacune, il est intéressant d’utiliser une approche spatialement explicite, qui représente la dynamique des populations par l’émergence des comportements des individus interagissant à la fois avec l’environnement et entre eux (McLane et al. 2011; Stillman et
al. 2015).
Les modèles centrés sur l’individu, « individual-based model, ou IBM », sont de plus en plus utilisés pour clarifier les incertitudes des changements futurs de l’environnement sur les populations animales. Par exemple, Imron et al. (2011) ont développé un IBM afin d’évaluer les conséquences de différentes utilisations des terres entourant le Parc national de Tesso Nilo sur la persistance de la population des tigres de
Sumatra (Panthera tigris sumatrae). Les impacts potentiels de parcs éoliens sur la masse, la productivité et la mortalité du Fou de Bassan (Morus bassanus) ont aussi été testés par IBM (Warwick-Evans et al. 2018). De plus, l’influence des modifications de l’environnement par les changements climatiques sur la durée des arrêts des canards colverts (Anas
platyrhynchos) aux haltes migratoires printanières (Beatty et al. 2017) et les variations de la
taille des parcelles, de la configuration et de la composition du paysage sur la persistance à long terme de quatre espèces (Nabe-Nielsen et al. 2010) ont également été traitées en simulant différents scénarios. Les IBMs permettent d’intégrer les comportements des animaux en réaction à leur environnement et donc, de mieux évaluer l’influence potentielle d’un changement de cet environnement sur leur répartition et leur abondance (McLane et
al. 2011).
Les modèles centrés sur l’individu
Les IBMs se basent sur une approche du bas vers le haut (« bottom-up ») pour comprendre la dynamique des populations selon différentes conditions environnementales. Les effets résultants sur la population émergent des actions cumulées des individus autonomes interagissant les uns avec les autres et avec leur environnement virtuel dans les simulations (Grimm et Railsback 2005). La modélisation du comportement des individus à fine échelle et l’utilisation d’environnements virtuels spatialement explicites offrent la possibilité d’estimer des propriétés émergentes difficilement prévisibles en observant uniquement les individus, et ainsi, de mieux guider la planification de la conservation de l’habitat des espèces menacées et leur gestion (Pressey et al. 2007). Les règles de déplacements établies pour reproduire le comportement des individus (McLane et al. 2011) peuvent, entre autres, inclure les motivations reliées à la quête de nourriture (Tardy et al. 2018), au risque de prédation (Latombe 2013) et à la compétition (Imron et al. 2011). Les individus simulés interagissent simultanément les uns avec les autres ainsi qu’avec leur environnement, simulant la dynamique complexe des systèmes écologiques (McLane et al. 2011).
Cette approche permet d’étudier des phénomènes ponctuels et temporels difficiles à mesurer dans le système réel ou à quantifier de manière empirique (DeAngelis et Moooij 2005). En considérant les individus de manière discrète, la variabilité spatiale des
interactions locales peut être prise en compte, ce qui peut ultimement affecter la dynamique de la population (Durrett et Levin 1994; Murrell 2005). Dans le cas de la dynamique prédateur-proie, par exemple, les composantes spatiales et temporelles de la réponse fonctionnelle des prédateurs (c.-à-d., les rencontres entre les individus et le temps passé par proie) peuvent être modélisées en évènement unique et être influencées par la non-uniformité locale des populations dans le paysage (Murrell 2005). De plus, l’interaction de l’individu avec son environnement permet de considérer certaines différences entre les individus. Par exemple, deux individus peuvent montrer des comportements différents seulement dus aux contraintes locales de leur environnement (Mason et Fortin 2017). Par conséquent, les IBMs permettent de tester différents scénarios de plans de gestion des espèces en péril, et de prévoir l’effet des modifications et de la configuration de l’environnement sur les populations, pour ainsi cibler les actions potentielles de gestion qui permettront d’atteindre les objectifs de protection.
L’utilisation des IBMs est particulièrement pertinente dans des contextes où la conservation doit concilier la protection et l’exploitation des ressources naturelles. Dans ces situations, l’utilisation d’expériences in situ pour tester des scénarios d’aménagement est difficile, voire impossible, compte tenu de la complexité du système et des hauts risques que peuvent subir les populations qui sont déjà dans un état de précarité démographique (Grimm et Railsback 2012; Latombe et al. 2014). Il existe de surcroît, une urgence d’agir avant que les connaissances essentielles à la protection d’une espèce menacée ne soient totalement rassemblées (Primack et al. 2012).
Le caribou forestier comme modèle d’étude
Le caribou des bois, écotype forestier (Rangifer tarandus caribou), est un animal menacé au Canada (COSEPAC, 2011) et vulnérable au Québec (MFFP, 2019), dont les populations sont au centre d’enjeux socioécologiques impliquant les industries d’exploitation des ressources ligneuses et la protection de son habitat. La perte, la fragmentation et la modification de l’habitat du caribou par les activités humaines constituent les principales causes de son déclin actuel (Environnement Canada 2011). La dynamique des populations de caribous est également affectée par la prédation (Seip 1992) des adultes par le loup gris (Canis lupus; Rettie et Messier 1998; Wittmer et al. 2005;
Équipe de rétablissement du caribou forestier du Québec 2013), et par celle des faons par le loup et l’ours noir (Basille et al. 2011; Pinard et al. 2012), qui sont intensifiées dans les paysages aménagés. Parallèlement, la régénération feuillue des jeunes peuplements après les coupes forestières offre un milieu de haute qualité pour l’orignal (Alces alces; Potvin et
al. 2005), favorisant la compétition apparente avec le caribou (Wittmer et al. 2007).
L’orignal étant la proie principale du loup (Bergerud et Elliot 1986), l’augmentation de son abondance dans l’environnement induit une réponse numérique du prédateur et, par ricochet, une pression accrue de prédation sur les caribous (Seip 1992; Wittmer et al. 2007). La conservation et la gestion des populations du caribou des bois dans la forêt commerciale aménagée représentent un défi considérable, impliquant le maintien d’une espèce en péril dans un environnement perpétuellement modifié par les activités industrielles de récoltes forestières (Parrott et al. 2012) dont les conséquences affectent l’habitat essentiel et les interactions trophiques des populations locales du caribou forestier.
La ligne directrice la plus influente pour la protection et la conservation du caribou est basée sur un modèle non spatial (voir Environnement Canada 2011). Le modèle perturbation-croissance de la population utilise des valeurs de recrutement propres au niveau de perturbation et un taux annuel moyen de survie des femelles adultes. Sommairement, pour qu’il soit probable (> 60 %) qu’une population soit autosuffisante (l³ stable) son aire de répartition devrait être composée au minimum de 65 % d’habitat non perturbé. Or, plusieurs études montrent que la structure spatiale de l’habitat affectant la sélection d’habitat par les individus (Fortin et al. 2008, 2013; Lesmerises et al. 2013) de même que leur vulnérabilité à la prédation (Courtois et al. 2007; Courbin et al. 2013; Fortin
et al. 2015; DeMars et Boutin 2018) influencent la survie des individus (Losier et al. 2015).
La présente étude vise à évaluer l’influence de l’organisation spatiale des coupes et des chemins forestiers selon divers niveaux de perturbation afin d’éclairer la gestion et la conservation de l’écotype forestier du caribou des bois et de son habitat essentiel. Nous avons utilisé le système « caribou-orignal-loup » pour analyser et modéliser dans un cadre prospectif et prévisionnel l’importance des effets spatiaux de l’environnement sur les interactions prédateur-proie.
L’influence des activités humaines sur le caribou des bois
Les modifications de l’environnement par les activités humaines influencent les déplacements et l’utilisation de l’espace par le caribou (Courtois et al. 2007; Courbin et al. 2009; Lesmerises et al. 2013). L’augmentation des perturbations affecte la fidélité aux sites des femelles caribous (Faille et al. 2010; Lafontaine et al. 2017), le taux de déplacements des individus et la taille de leur domaine vital (Smith et al. 2000). L’utilisation des parcelles de forêts résiduelles dans un paysage perturbé par la coupe est influencée par sa composition, sa taille et le milieu environnant (Lesmerises et al. 2013). Notamment, on observe un effet de bordure, créé par la juxtaposition des coupes et des routes aux forêts non perturbées, pouvant s’étendre à l’intérieur de la forêt jusqu’à 4 km de la zone de coupe, ce qui représente une grande proportion du milieu et affecte la répartition des individus (Fortin et al. 2013). Par conséquent, l’utilisation des habitats jugés essentiels pour les caribous n’est pas uniforme dans l’espace, et la densité et la répartition spatiale des individus détermineront alors la probabilité de rencontre avec leur prédateur dans un environnement donné.
Contrairement au caribou, les coupes et les routes avantagent le loup. Les routes sont fréquemment utilisées par les loups, car elles facilitent leurs déplacements (James et Stuart-Smith 2000; Kittle et al. 2017). Notamment, les loups se déplacent de deux à trois fois plus vite sur les routes qu’en milieu forestier (Dickie et al. 2017), augmentant de fait leur efficacité de chasse. Également, DeMars et Boutin (2018) ont montré que la présence des routes à travers des habitats refuges du caribou augmente la sélection de ce milieu par les loups; le risque de prédation des caribous est alors plus élevé à proximité des routes (James et Stuart-Smith 2000; Latham et al. 2011; Whittington et al. 2011; Fortin et al. 2015). Conséquemment, la disposition des coupes et des routes joue un rôle fondamental dans la probabilité de rencontre entre le caribou et le loup dans un paysage perturbé. Cependant, la dynamique prédateur-proie de ce système dépend également de la présence de l’orignal, la proie principale du loup.
Le système caribou-loup-orignal
La présence de l’orignal doit être prise en compte pour bien comprendre la prédation du loup sur le caribou pour trois raisons principales. Tout d’abord, l’augmentation des populations de loups dans le paysage est essentiellement reliée à la hausse du nombre d’orignaux (Messier et Crête 1985). Par conséquent, l’augmentation de la pression de prédation sur le caribou découle de la réponse numérique de la population d’orignaux suite aux changements de l’environnement. Ensuite, le temps passé par le loup à manipuler et à consommer une proie diminue le temps consacré à une seconde proie (McLellan et al. 2010). Ainsi, la prédation des caribous peut être différente de celle estimée sans la présence de l’orignal dans le système. Enfin, l’hétérogénéité du paysage influence le patron spatial des interactions entre les trois espèces (Courbin et al. 2009, 2013; Fortin et
al. 2015). La présence des coupes et des routes influence différemment l’utilisation du
paysage par les trois espèces (Fortin et al. 2013, 2015; Gagné et al. 2016). Par exemple, la probabilité relative d’occurrence des orignaux augmente graduellement jusqu’à atteindre un plateau à environ 3 km de la route (c.-à-d., à 3 km ou à 8 km de la route la probabilité d’occurrence d’un orignal est la même), alors que la probabilité relative d’occurrence des caribous augmente pour atteindre une valeur maximale à environ 4 km, avant de redescendre à de plus grandes distances (Fortin et al. 2015). Ces différences entre l’orignal et le caribou sont responsables de la hausse des interactions à distance des coupes et des routes entre le caribou et le loup (Fortin et al. 2015). La présence de l’orignal dans les modèles prévisionnels permettra donc de mieux comprendre les interactions prédateur-proie de ce système complexe.
L’aménagement de l’habitat du caribou des bois jusqu’à maintenant
Afin de maintenir les populations de caribous, des lignes directrices pour l’aménagement de son habitat ont été suggérées autant à l’échelle globale (pour l’ensemble de populations au Canada) qu’à l’échelle régionale (spécifique aux provinces ou aux populations locales). À ce titre, Environnement Canada (2011) suggère de ne pas dépasser un seuil critique de 35 % de perturbation totale dans l’habitat du caribou : ce niveau de perturbations permettant une probabilité de persistance de 60 % des populations. Dans leur étude, les zones perturbées comprennent les perturbations naturelles, c.-à-d., les feux de
moins de 40 ans et les perturbations anthropiques, notamment les coupes âgées de moins 50 ans et les routes, avec une zone tampon de 500 m. Cependant, la répartition spatiale des perturbations dans les territoires aménagés n’est pas spécifiquement prise en compte. Au Québec, Courtois et al. (2004) ont recommandé une approche écosystémique en concentrant les coupes forestières pour imiter les perturbations par le feu, et en protégeant de grands massifs forestiers interconnectés pour le caribou. Le déploiement de cette approche dans la région de la Côte-Nord a toutefois montré que les massifs entourés de coupes étaient moins intensivement fréquentés par les caribous que ceux adjacents à la forêt non perturbée (Courtois et al. 2008). En plus de la qualité intrinsèque des massifs (c.-à-d., la taille et le type de peuplement), la connectivité entre les massifs serait un facteur important affectant l’utilisation de l’habitat et la répartition spatiale du caribou (O’Brien et
al. 2006; Courbin et al. 2014). Cette stratégie d’agrégation des coupes permettrait
l’extraction d’un niveau relativement élevé de bois par les industries forestières sans compromettre la persistance du caribou boréal (Cyr et al. 2017), en plus de réduire la construction des routes et l’étendue de l’empreinte humaine sur l’environnement (Tittler et
al. 2012). Or, l’impact de la perte de l’habitat sur la survie des populations, dans ces études,
suppose généralement des interactions prédateur-proie uniformes à travers les milieux perturbés, pourtant, la configuration spatiale du paysage peut influencer ces interactions. Même si le seuil critique de l’habitat essentiel à préserver recommandé par Environnement Canada (2011) est respecté, la variabilité de la prédation au sein du paysage peut mener à des résultats inattendus diminuant l’efficacité des plans de gestion (p. ex. voir Mahoney et
al. 2018).
Objectif de l’étude
L’objectif de cette étude est d’évaluer l’impact du niveau de perturbation totale et de la répartition spatiale des coupes et des routes sur la mortalité du caribou des bois coexistant avec le loup gris et l’orignal. Pour ce faire, des scénarios d’aménagement forestier virtuels ont été développés variant selon deux attributs principaux : 1) le niveau de perturbation totale (entre 10 % et 75 % du paysage perturbé) et 2) la configuration spatiale des parterres de coupes, soit en mosaïque, en petites ou grandes agglomérations. Plus spécifiquement, nous avons vérifié si pour un même niveau de perturbation totale, la
répartition spatiale des coupes et des routes engendrera des différences du nombre de caribous et d’orignaux tués par le loup et cela durant trois saisons distinctes, la mise bas, la fin de l’été et l’hiver. Également, nous avons évalué quels attributs des aménagements forestiers, parmi la densité de bordures (zones de contact entre les perturbations et les peuplements non perturbés) et la densité de routes, affectaient davantage la mortalité saisonnière des proies.
Approche empirique
Étant donné la complexité et la dynamique du système, un modèle centré sur l’individu a été utilisé pour tester divers scénarios d’aménagement forestier. Le modèle a donc été paramétré et validé avant d’en faire usage pour générer les données nécessaires afin de répondre à nos objectifs. Pour que les agents se déplacent dans le paysage virtuel tel que le feraient les individus des espèces à l’étude, nous avons paramétré l’IBM à l’aide de règles de déplacements établies à partir des données de suivis GPS faits sur 60 caribous, 15 orignaux et sept loups au Québec. Nous avons développé des fonctions de sélection de pas (Fortin et al. 2005) caractérisant les déplacements de chaque espèce en termes de distances parcourues, d’angles de direction et de réactions aux divers types de couverts forestiers et aux routes. Cette information a ensuite été utilisée dans l’IBM pour recréer les déplacements des individus dans un environnement hétérogène. La prédation par le loup a été simulée en décomposant les évènements de prédation en quatre étapes : 1) la détection, 2) l’attaque, 3) l’abattage de la proie et 4) la consommation et le repos du prédateur suite à la chasse. La distance de détection de la proie, le succès d’abattage et la période stationnaire (c.-à-d., le temps avant de recommencer à chasser) ont été tirés de la littérature (Hayes et al. 2000; Mech et Boitani 2003; Wikenros et al. 2009; Latombe 2013) tandis que la probabilité d’attaque suite à la détection de la proie a été calibrée en ajustant la valeur par itération jusqu’à ce que le nombre de mortalités durant les simulations corresponde aux valeurs souhaitées du nombre de mortalité. Par la suite, nous avons validé notre modèle en comparant les sorties de 150 simulations avec les données empiriques de la région pour laquelle nous avions un suivi des trois espèces. La validation du modèle a permis de s’assurer que les individus simulés reproduisaient adéquatement le comportement (c.-à-d., l’utilisation des différents types de couverts et la distance parcourue entre deux points) des
individus suivis par télémétrie, et ce, sous les mêmes conditions de paysage. Pour les simulations des scénarios d’aménagement forestier, nous avons ajusté le nombre d’orignaux et de loups virtuels en fonction du taux de perturbation, afin de reproduire la réponse numérique de ces populations suivant l’enfeuillement des parterres de coupes. Finalement, suite aux simulations des scénarios d’aménagement forestier, nous avons évalué le nombre d’agents de chaque espèce de proies tués en fonction du niveau de perturbation totale, du type de configuration spatiale des coupes et de la densité de bordures et de routes.
Chapitre principal. Management-mediated predation
rate in the caribou-moose-wolf system: spatial design of
logging activities matters
VIRGINIE VANLANDEGHEM1, PIERRE DRAPEAU2, MARIE-CAROLINE PRIMA1,
MARTIN-HUGUES ST-LAURENT3 AND DANIEL FORTIN1
1Département de biologie, Université Laval, Québec, Qc, Canada
2Département de biologie, Université du Québec à Montréal, Montréal, Qc, Canada
Résumé
La structure complexe du paysage peut dicter la dynamique des populations des proies et des prédateurs interagissant. Pourtant, les plans de gestion des espèces sont généralement développés à partir de modèles de prévision non spatiaux. Omettre cet aspect peut mener à des résultats inattendus et même compromettre la possibilité d’effectuer des interventions spatiales susceptibles d’accroître l’efficacité des plans. La gestion de la population boréale du caribou des bois (Rangifer tarandus caribou), un animal menacé au Canada, basée sur une approche non spatiale en est un exemple. Présentement, la recommandation de gestion ayant le plus d’influence est de maintenir au minimum 65 % de forêt non perturbée dans les aires occupées par les populations de caribous, et ce, peu importe la configuration spatiale des interventions forestières. En utilisant un modèle spatialement explicite basé sur l’individu (IBM), nous avons évalué l’effet de la configuration spatiale des coupes et des routes sur la mortalité du caribou forestier vivant en interaction avec le loup gris (Canis lupus) et l’orignal (Alces alces), son compétiteur apparent. En se basant sur un paysage forestier réel, nous avons appliqué des scénarios d’aménagement forestier variant selon des configurations spécifiques de coupes (c.-à-d., en mosaïque, en petite ou en grande agglomération) avec des niveaux de perturbations croissants. L’IBM simulait des agents, représentant les individus de chaque espèce, se déplaçant selon des règles déterminées empiriquement à partir des données d’individus suivis par télémétrie. Nos résultats montrent que le comportement de déplacement des individus porteurs d’un collier GPS était influencé par les types de couvert et la distance à la route, et différait entre les espèces et les saisons. Par exemple, les caribous et les orignaux évitaient les endroits près des routes alors que les loups les utilisaient. Ces différences ont eu un effet direct sur la mortalité des agents caribous, laquelle ne dépendait pas uniquement du niveau de perturbations, mais également de la configuration spatiale des coupes et des routes. Après avoir contrôlé le niveau de perturbations, l’analyse indique que les loups tuaient plus de proies lorsque la configuration de l’aménagement forestier créait un réseau routier dense et étendu et augmentait la fragmentation du paysage. Ainsi, la mortalité des caribous était plus faible dans les paysages pour lesquels la densité de routes et de bordures créées par les perturbations étaient faibles. Notre étude indique que même si le niveau de perturbations de l’habitat respecte le niveau maximal de perturbation
recommandé, les populations de proies pourraient continuer à décliner simplement parce que l’hétérogénéité du paysage créée par la gestion de l’habitat fait en sorte que les rencontres prédateur-proie y sont plus fréquentes. Notre étude démontre comment la gestion du paysage peut être utilisée pour manipuler les interactions entre espèces dans le but d’augmenter ou de diminuer le taux de prédation de populations cibles selon les objectifs de gestion.
Abstract
Landscape complexity can determine the population dynamics of interacting predators and prey. Yet, management plans are commonly developed from aspatial predictive models. This oversight may result in unexpected outcomes or in the loss of opportunities to make spatial interventions that would increase the plan’s effectiveness. The management of the threatened woodland caribou (Rangifer tarandus caribou), boreal population, provides a recent example of such an aspatial approach. Currently, the most influential management recommendation is to maintain at least 65 % of undisturbed forests in areas occupied by caribou populations, regardless of the spatial configuration of the forest cover. Using a spatially explicit individual-based model, we evaluated the effects of the spatial configuration of cuts and roads on the mortality of boreal caribou living in sympatry with wolves (Canis lupus) and moose (Alces alces), an apparent competitor. Starting with a real forest landscape, we created forest management scenarios of specific spatial distribution of cuts (mosaic, small- or large agglomeration) with increasing levels of disturbance. The IBM simulated agents, representing individuals of the three species, moving according to movement rules determined from radio-collared individuals. Our results indicated that movement responses to land cover types and roads differed among species. For example, caribou and moose generally avoided areas close to roads, contrary to wolves. Those differences influenced the mortality of caribou agents which did not only depend on the levels of disturbance, but also on the spatial distribution of cuts and roads. After controlling for the levels of disturbance, wolves were more successful when forest management required an extensive road network resulting in relatively high habitat fragmentation. Accordingly, caribou agents experienced lower mortality in landscapes with low densities of road and disturbance-related edges. Our study indicates that even if the currently prescribed level of habitat disturbance is not exceeded, prey populations could still decline simply because the landscape configuration created by habitat management produces relatively high encounter rates between predators and prey. Our study demonstrated how landscape management could be used to manipulate species interactions, with the intent of either increasing or decreasing predation rates on specific populations, depending on management goals.
Introduction
The movement response of individuals to landscape features often varies among species, a difference sufficient to determine interaction rates between opponents and, ultimately, population and community dynamics (Holt 1984; Johnson et al. 1992; Morales et al. 2010; DeMars and Boutin 2018). Spatial heterogeneity in landscape attributes thus can govern food web interactions (Murrell 2005; Kauffman et al. 2007; Gorini et al. 2012). The key role of spatial heterogeneity has become increasingly recognized following Huffaker's (1958) landmark experiment. His results showed that landscape complexity could stabilize predator-prey dynamics by reducing the rate of interactions among opponents and promote species co-existence (Huffaker 1958), under conditions where extinction would be expected in homogeneous environments (Gause et al. 1936). Landscape composition and structure thus should be considered when studying population dynamics (Wiegand et al. 1999), but spatial patterns in vital rates remain largely overlooked in population studies.
Spatial heterogeneity has relevance beyond fundamental studies of population and community ecology. Wildlife management and conservation relies on the accurate forecast of biological system (Wiegand et al. 1999; Fryxell et al. 2005; Sibly et al. 2009; Kareiva and Marvier 2015; Warwick-Evans et al. 2018) a challenge that may require spatially explicit approaches (Evans 2012; Stillman et al. 2015). For example, the spatial model of Mahoney et al. (2018) for coyote (Canis latrans) lethal control in a mule deer (Odocoileus
hemionus) system demonstrated that predictions could be more reliable and management
more effective by considering specific attributes of landscape heterogeneity. Implementing management plans developed from non-spatial models can have unexpected ¾ even undesirable ¾ consequences on the system because the result of management actions may vary with landscape composition and physiognomy (Wiegand et al. 1999; Mahoney et al. 2018).
A persisting challenge of conservation planning lies in the preservation of organisms despite ongoing habitat changes due to the exploitation of natural resources (Margules and Pressey 2000; Kareiva and Marvier 2015). Decision-makers can rely on
scenario planning, which explores the uncertainty surrounding the future consequences of a decision by contrasting scenarios of a plausible future (Peterson et al. 2003). Scenarios can be tested with individual-based models (IBMs) that account for detailed knowledge of animal behaviors (e.g., movement, foraging, memory) in spatially explicit and dynamic environments (Grimm and Railsback 2005; McLane et al. 2011). IBMs are particularly useful when in situ experiments are difficult or impossible to conduct because of the complexity, size, and slow dynamics of ecological systems (Grimm and Railsback 2012). This situation represents well the biological system involving the boreal population of woodland caribou (Rangifer tarandus caribou) in managed boreal forests.
The boreal population of woodland caribou (hereafter boreal caribou) is threatened in Canada (COSEWIC 2014). Boreal caribou are under top-down control (Seip 1992), and predation by gray wolves (Canis lupus) is considered the main driver of population decline (Rettie and Messier 1998; Wittmer et al. 2005; Équipe de rétablissement du caribou forestier du Québec 2013). Anthropogenic activities increase habitat loss and fragmentation, which tends to intensify predation rate on caribou. Indeed, roads facilitate the movement of wolves across caribou ranges (James and Stuart-Smith 2000; DeMars and Boutin 2018) and enhance hunting success (Whittington et al. 2011; Dickie et al. 2017). Moreover, deciduous vegetation invades cutovers few years after logging (Potvin et al. 2005; Gagné et al. 2016), which is the primary food source for moose (Alces alces; Crête 1989). This increase in vegetation can trigger an increase in moose abundance, followed by a numerical response of the wolf population that can exacerbate predation risk for boreal caribou (Seip 1992; Wittmer et al. 2007). Because of such negative impact of human-induced activities, Environment Canada (2011) recommended that critical habitat for boreal caribou should contain less than 35 % of total disturbance, as defined by anthropogenic disturbances (plus a 500-m buffer) and by fires. With this threshold, a local population has a 60 % probability of being self-sustaining (l ≥ 1), given national averages in vital rates. However, the demographic consequence of maintaining at least 65 % of undisturbed habitat varies among populations (Environment Canada 2012; Fortin et al. 2017; Rudolph et al. 2017), in part because spatial patterns of landscape composition are likely to influence the spatial game of interactions between caribou, wolves and moose differently (Fortin et al. 2015). The three species do not react similarly to cuts, roads and other landscapes features
(Fortin et al. 2013, 2015; Gagné et al. 2016), and those species-specific responses generate spatial patterns in predator-prey co-occurrence (Courbin et al. 2009, 2013) and in prey mortality (Wittmer et al. 2007; Fortin et al. 2015; Losier et al. 2015; Serrouya et al. 2019). The caribou-moose-wolf system thus provides an opportunity to use IBMs to inform conservation planning by revealing how different spatial patterns of human disturbances could influence predation rates.
In this study, we used a spatially explicit IBM to evaluate the effects of landscape configuration on caribou mortality for a population living in sympatry with wolves and moose in managed boreal forests. Specifically, we used realistic management scenarios differing in terms of spatial distribution of cuts and roads and levels of total disturbance to evaluate the rate of caribou mortality and forecast consequences on local populations. Caribou, moose, and wolf agents moved in the virtual landscapes following rules that were empirically determined from the movement of GPS-collared individuals of each species. Our assessment of mortality rates of caribou and moose agents demonstrates the value of spatially explicit models to inform the decision-making process regarding wildlife conservation.
Materials and methods
FOREST MANAGEMENT SCENARIOS
We applied 12 virtual management scenarios to the undisturbed Montagne-Blanche region (50°33‘N to 51°18’N, 69°46’W to 70°59’W) included in the Saguenay-Lac-St-Jean and the Côte-Nord area of Québec (Canada), representing a typical boreal landscape. We used a 1:20,000 ecoforest map provided by the Ministère des Forêts, de la Faune et des Parcs (hereafter MFFP) as our base map, which among other information, characterized tree species composition and stand age of forested areas. Our scenarios (5065 km2, with a
25-m resolution) thus represented realistic landscape composition and configuration that individuals might encounter in nature. Furthermore, by using the same base map for all management scenarios, we were able to focus on the impact of different spatial distributions of cuts and roads on space use of caribou, moose, and wolves and on prey mortality. The total of 13 scenarios consisted of 12 management scenarios varying in terms of the levels of total disturbance and spatial distribution of cuts and roads, together with a
scenario of reference representing a landscape undisturbed by human activities (the original map of the region; Fig. 1). As per Environment Canada (2011), the levels of total disturbance were calculated as the percentage of the landscape of the non-overlapping surface of burns, roads, and cuts, the latter two buffered by 500-m. Burned areas covered 7 % in all 13 landscapes. Cuts and roads were simulated in the 12 disturbed landscapes in order to obtain ranges of total disturbance of approximatively 30 %, 45 %, 60 %, and 75 % of the landscape. Besides, cutblocks were organized in mosaic, small or large agglomerations. Mosaic cutting consisted of dispersed clear-cuts with at least a 200-m buffer of residual forest around them and a 100-m buffer when cutblocks were < 0.25 km2
(MFFP 2018). Mosaic cutting thus has a checkboard pattern of residual and logged forest patches of 85-100 ha each. Small-agglomeration cutting involved the grouping of adjacent cutblocks in sectors of 10-50 km2, whereas large-agglomeration cutting grouped cutblocks
in larger sectors of 40-150 km2 (Fig. 1). For a fixed level of disturbance, the total amount
of cuts and roads differed between the three management strategies due to differences in the level of overlap between the 500-m buffers and differences in the roads required to connect the cutblock network (Fig. 2).
Forest management scenarios were generated using Remsoft's Woodstock and
Stanley planning models 2017.11 (Walters et al. 1999; Remsoft Incorporated, Fredericton,
New Brunswick, Canada). The base map was split into spatial organization compartments (SOC) of 50–150 km2 generally used by the MFFP (2018) to create spatial patterns of
logging in line with current forest management harvesting guidelines. Each SOC was gradually available to harvest from south to north to consider the road network’s deployment. Cuts in an available SOC occurred over 10 years and only stands that reached the exploitability age were harvested (as determined by annual growth rate curve; Pothier and Savard 1988). The targeted level of harvest was reached after a 50 years-period in each simulated landscape. Simulations of the IBM were then run on these resulting landscapes. Mosaic and small-agglomeration cutting were simulated using the Stanley planning model, whereas large-agglomeration cuttings were created with the AAunit module of the
Woodstock planning model. Road networks were constructed to connect centroids of
harvested stands. First, we defined the same starting point for all scenarios, which was randomly selected within the south part of the landscape to create the first link of the road
network. Second, we delimited a polygon for each set of adjacent cuts. For each of those polygons, we identified the centroid. Third, we connected each centroid to the nearest connected centroid using least-cost-path algorithm to avoid crossing large rivers (i.e., water had high value of cost and the rest had low value). Finally, roads were transformed in 25-m wide polygons. ArcGIS 10.2.2 (ESRI 2014) and R software (R Development Core Team 2016) were used to create road networks and to incorporate them in forest management scenarios.
Figure 1. Representation of the landscape (5065 km2) in which forest management scenarios
were simulated for assessing the influence of the spatial configuration of cuts and roads on caribou mortality. Enlargements (404 km2) show the spatial distribution of cuts and roads according to the three
management strategies: a. mosaic, b. small agglomeration cutting and c. large agglomeration cutting. Aggregation of cuts directly affects the total area disturbed (without the 500-m buffer: mosaic = 62.4 km2, small agglomeration = 60.0 km2, large agglomeration = 57.5 km2; with the 500-m buffer: mosaic
= 256 km2, small agglomeration = 155 km2, large agglomeration= 147 km2). Other colors represent
land cover types: red = burned area, blue = water body, dark green = closed-canopy conifer forest, green = open conifer forest without lichen, light green = open conifer with lichen, orange = mixed and deciduous and purple = other.
MOVEMENT RULES DERIVED FROM RADIO-TRACKING CARIBOU, MOOSE AND WOLVES
To identify species-specific movement rules that were implemented in the IBM, we radio-tracked caribou, moose, and wolves in landscapes under different levels of forest management. Radio-collared caribou were followed in the study area (48˚N-52˚N 66˚W-80˚W) covering approximatively 380 000 km2 of the boreal forest in the Jamésie, Saguenay
and Côte-Nord regions of Québec, Canada, whereas radio-collared moose and wolves were followed over 11 500 km2 in the Côte-Nord region. Forest harvesting is the main industrial
activity occurring in those regions. Caribou densities vary between 1.6 and 2.5 individuals/100 km2 (mean density throughout the area is 1.5 caribous/100 km2; Équipe de
rétablissement du caribou forestier du Québec 2013) and moose occur at densities ranging Figure 2. Relationship between road (a.), edge densities (i.e., perimeter of 500-m buffer around cuts and roads) (b.) and proportion of cuts (c.), as a function of the proportion of total disturbance (burned areas and cuts and roads with 500-m buffer) for each of the three forest management strategies (i.e., mosaic, small-agglomeration and large-agglomeration cutting). Overall relationships are represented by the dashed line.
between 4.3 and 10.0 individuals/100 km2 (Fortin et al. 2008; Lefort and Massé 2015).
Although poorly documented, wolf density should range between 0.3 and 1.5 individuals/100 km2 with mean territory size of 1000 km2 (Larivière et al. 1998; Jolicoeur
and Hénault 2002).
A total of 60 female caribou were fitted with GPS collars (i.e., 2200 or 3300 model, Lotek Engineering Inc., Newmarket, Canada or TGW 3600 model, Telonics Inc., Mesa, AZ, USA or GPS-Plus model, Vectronic Aerospace, Berlin, Germany; Argos/GPS collars, TGW 3680 or TGW 4680 model, Telonics Inc.; and GlobalStar/GPS collars, Track M model, Lotek Engineering Inc., Newmarket, ON, Canada). Caribou were relocated every 1-h for an average of 15 months (range 1-48 months) between 2005 and 2014. Fifteen female moose were followed with GPS collars or Argos/GPS collars from March 2005 to March 2009 with relocations taken every hour. Seven GPS-collared (GPS 3300SW) wolves from four different packs were also relocated every hour, two days a week, from 2005 to 2010. Individual moose and wolves were monitored for an average of 17 and 19 months, respectively. Following Basille et al. (2013), we defined three periods by merging caribou and moose biological seasons: pre-calving/calving season (mid-May through late July, which covers the calving season of both boreal caribou and moose), late-summer (early August to the end of September) and winter (October to mid-May). Wolf locations were also separated according to the same periods.
We characterized the study area using Landsat Thematic Mapper image taken in 2000 with a 25-m resolution grid (Natural Resources Canada, Canadian Forest Service). The initial 48 land cover categories were regrouped into seven classes: closed-canopy conifer forest, open conifer forest with lichen, open conifer forest without lichen, mixed and deciduous forest, lichen-heath community, water body and other (including heath without lichen, open area, wetland and unclassified areas, following Courbin et al. 2009). Land cover maps were updated every year by adding recent cuts (0-20 years old), old cuts (21-50 years old), roads, and fires (0-50 years), which resulted in a total of 11 land cover types.