Étiologie génétique-environnementale de la relation
entre le contrôle maternel et la flexibilité cognitive de
l'enfant à l'âge préscolaire
Mémoire doctoral
Samuel Dallaire
Doctorat en psychologie
Docteur en psychologie (D. Psy.)
Étiologie génétique-environnementale de la relation
entre le contrôle maternel et la flexibilité cognitive de
l’enfant à l’âge préscolaire
Mémoire doctoral
Samuel Dallaire
Sous la direction de :
Résumé
Les fonctions exécutives (FE) jouent un rôle important dans plusieurs sphères du développement de l’enfant. Il s’avère donc crucial de mieux comprendre leur étiologie. À l’aide d’un devis génétiquement informatif, cette étude examine l’étiologie génétique-environnementale d’une composante importante des FE, soit la flexibilité cognitive, et les associations phénotypiques et étiologiques entre celle-ci et le contrôle maternel à l’âge préscolaire. Des jumeaux (400 monozygotes et 534 dizygotes) provenant de l’Étude des jumeaux nouveau-nés du Québec ont été évalués à l’âge de 5 ans. La flexibilité cognitive a été mesurée à l’aide d’une tâche informatisée et le contrôle maternel à l’aide d’un questionnaire autorapporté. Un modèle génétique univarié démontre que les différences individuelles dans la flexibilité cognitive sont entièrement expliquées par des facteurs environnementaux (incluant l’erreur de mesure). Des corrélations démontrent ensuite que les jumeaux exposés à plus de contrôle maternel performent moins bien à la tâche mesurant la flexibilité cognitive (r = -.13, p < 0.001). Un modèle génétique bivarié, permettant de décomposer les contributions génétiques et environnementales sous-jacentes à la relation entre le contrôle maternel et la flexibilité cognitive, démontre que cette relation est essentiellement expliquée par des facteurs issus de l’environnement unique (Re = -0.13, p < .05). Cela suggère que les mécanismes qui relient la flexibilité cognitive aux pratiques parentales sont environnementaux et opèrent indépendamment des facteurs génétiques transmis par le parent à l’enfant. Cette étude représente un premier pas vers une meilleure compréhension des mécanismes qui sous-tendent le développement de la flexibilité cognitive à l’enfance et la relation entre elle et les comportements parentaux.
Table des matières
Résumé ... ii
Table des matières ... iii
Liste des tableaux... iv
Liste des figures ... v
Remerciements ... vi
Introduction ... 1
Les comportements parentaux en tant que prédicteur du développement des FE ... 2
Mécanismes génétiques ou environnementaux ... 3
Études de jumeaux... 5
Étiologie des FE ... 6
Étiologie des comportements parentaux ... 10
Modèles génétiques bivariés ... 11
Le contrôle parental et le développement des FE ... 12
Objectifs ... 16 Chapitre 1 : Méthodologie ... 17 Participants et procédure... 17 Mesures ... 18 Plan d’analyse ... 20 Chapitre 2 : Résultats ... 22 Analyses préliminaires ... 22
Modèles génétiques univariés ... 22
Modèles génétiques multivariés ... 23
Chapitre 3 : Discussion ... 25
Interprétation des résultats ... 25
Conclusion ... 35
Bibliographie ... 36
Liste des tableaux
Tableau 1 - Statistiques descriptives et corrélations intraclasses pour la flexibilité cognitive et le contrôle maternel ... 47 Tableau 2 - Modèles génétiques univariés : décomposition des contributions génétiques (A) et de l’environnement partagé (C) et unique (E) à la flexibilité cognitive et au contrôle maternel ... 48 Tableau 3 - Modèles génétiques bivariés : décomposition de la variance génétique (A) et environnementale (C et E) commune à la flexibilité cognitive et au contrôle maternel ... 49 Tableau 4 - Analyses de régression pour prédire les scores de discordance entre jumeaux MZ sur le plan de la flexibilité cognitive à partir des scores de discordance sur le plan du contrôle maternel ... 50
Liste des figures
Figure 1. Modèles univariés décomposant les contributions génétiques (A) et de
l’environnement partagé (C) et unique (E) de la flexibilité cognitive et du contrôle maternel (séparément). ... 51
Figure 2. Modèle bivarié (Décomposition de cholesky) ACE de la flexibilité cognitive et du
Remerciements
Ce mémoire est l’aboutissement d’un parcours doctoral parsemé d’apprentissages et de défis, qui n’aurait pas été possible sans l’aide et le soutien de plusieurs personnes. Je tiens d’abord à remercier ma directrice de recherche, Célia Matte-Gagné, pour ton support et l’importance que notre réussite prend dans ton travail. Que ce soit pour les opportunités d’emplois, les encouragements, la bienveillance ou le support que tu m’as accordé, il n’y a aucun doute pour moi que je suis privilégié d’avoir pu travailler avec toi comme directrice.
Je remercie mon groupe de recherche (Groupe de recherche en inadaptation psychosociale – GRIP Ulaval). Merci Bei pour ton aide indispensable sur le plan statistique. Merci Michel, pour tes rétroactions constructives et ton implication tout au long de mon doctorat en tant que membre de mon comité. Merci Nadine pour ton aide et tes bons conseils qui m’ont permis de mieux comprendre les études en génétique. Je remercie aussi les familles participantes dans l’Étude des jumeaux nouveau-nés du Québec (EJNQ). Je remercie également les Fonds de Recherche du Québec – Société et Culture (FRQSC) qui m’ont offert des bourses d’études qui m’ont permis de me consacrer pleinement à mes études.
Enfin, je remercie mes amis pour tous les beaux moments et escapades que nous avons partagés au long de ce beau parcours. Merci à ma conjointe, Alexane, pour ton soutien, tes encouragements et ta solidarité dans toutes ces étapes de mon parcours doctoral. Merci à mes parents qui m’ont toujours encouragé à poursuivre mes études et de m’avoir fourni le soutien nécessaire afin de réussir mes études. Merci à Charlie et Mango, mes deux chiens sans qui les périodes de rédaction à la maison n’auraient jamais été remplies d’autant d’affection. J’ai tant appris durant cette expérience qui m’a permis de cheminer vers l’adulte que je suis aujourd’hui. À tous ceux qui ont été présents durant cette aventure, je vous en suis reconnaissant.
Introduction
Depuis quelques années, la psychologie du développement porte un intérêt grandissant au développement des fonctions cognitives supérieures impliquées dans le contrôle volontaire des pensées, émotions et actions, soit les fonctions exécutives (FE; Diamond, 2013; Garon, Bryson, & Smith, 2008). Les FE regroupent plusieurs habiletés cognitives qui sont cruciales pour l’adaptation sociale, comportementale et scolaire de l’enfant telles que la mémoire de travail, la flexibilité cognitive et l’inhibition (Blair, 2002; Diamond, 2006; Miyake et al., 2000). L’inhibition fait référence à la capacité d’inhiber une réponse prédominante qui n’est pas nécessaire à l’atteinte d’un but visé (Carlson, Moses, & Breton, 2002). La mémoire de travail réfère à la capacité de retenir et manipuler mentalement de l’information (Carlson et al., 2002) tandis que la flexibilité cognitive constitue la capacité de sélectionner de manière adaptative une nouvelle réponse en fonction d’un changement dans la situation (Chevalier & Blaye, 2006; Miyake et al., 2000). Comme on a longtemps cru que ces fonctions cognitives complexes ne se développaient qu’à l’adolescence (Chevalier, 2010), l’étude des FE chez les jeunes enfants est récente (Carlson, Mandell, & Williams, 2004; Hughes & Ensor, 2005). Ainsi, la compréhension des facteurs et mécanismes qui mènent à leur développement demeure limitée. La poursuite de la recherche dans ce domaine est importante puisque les FE jouent un rôle déterminant dans la prédiction de plusieurs aspects du développement de l’enfant (Zelazo, 2015).
Durant l’enfance, les différences individuelles dans les FE sont notamment associées au rendement académique (Best, Miller, & Jones, 2009; Biederman et al., 2004; Diamond, 2013; Monette, Bigras, & Guay, 2011; van der Stel & Veenman, 2014; Willoughby, Wylie, & Little, 2019), à la compétence sociale et morale (Camerota, Willoughby, & Blair, 2019; Clark, Prior, & Kinsella, 2002; Kochanska, Murray, & Harlan, 2000), à la compréhension des émotions (Martins, Osório, Veríssimo, & Martins, 2016), à la théorie de l’esprit (Cassetta, Pexman, & Goghari, 2018; Devine & Hughes, 2014; Poulin-Dubois, & Yott, 2014), l’agressivité (Baker, Jensen, & Tisak, 2019) et la régulation émotionnelle (Andres, Castaneiras, Stelzer, Juric, & Introzzi, 2016; Carlson & Wang, 2007; Simonds, Kieras, Rueda, & Rothbart, 2007). De nombreuses études laissent même entendre que la performance
de l’enfant a des tâches mesurant différentes FE serait un meilleur prédicteur de la réussite scolaire que le quotient intellectuel (QI; Alloway & Alloway, 2010; Blair, 2002; Blair & Razza, 2007; Borella, Carretti, & Pelegrina, 2010). D’autre part, des déficits sur le plan des FE ont été associés à plusieurs difficultés psychologiques et développementales à l’âge scolaire, notamment l’agressivité (Baker, Jensen, & Tisak, 2019; Rohlf, Holl, Kirsch, Krahé, & Elsner, 2018), les symptômes d’inattention et d’hyperactivité (Berenguer, Rosello, Colomer, Baixauli, & Miranda, 2018; Clark et al., 2002; Sasser, Beekman, & Bierman, 2015) et l’autisme (Kenny, Cribb, & Pellicano, 2019; Pennington & Ozonoff, 1996). À l’âge adulte, des déficits dans les FE sont reliés à des difficultés interpersonnelles, à de l’agressivité, de l’instabilité émotionnelle, à un taux plus élevé de criminalité (Denson, Pedersen, Friese, Hahm, & Roberts, 2011; Moffitt et al., 2011; Wallinius, Nordholm, Wagnstrom, & Billstedt, 2019; Winstok, 2009) et une moins bonne qualité de vie (Davis, Marra, Najafzadeh, & Liu-Ambrose, 2010). Compte tenu du rôle important que jouent les FE dans plusieurs sphères du développement, il s’avère crucial de mieux comprendre leurs prédicteurs.
Les comportements parentaux en tant que prédicteur du développement
des FE
Il est maintenant largement accepté que les expériences relationnelles précoces jouent un rôle dans le développement cognitif de l’enfant (Belsky & De Haan, 2011; Kraybill & Bell, 2013). Cela serait attribuable au fait que l’environnement, surtout les expériences relationnelles, interagit avec le bagage génétique pour marquer de façon durable le développement du cerveau pendant les premières années de vie (Kolb et al., 2012). Il est d’ailleurs démontré que la trajectoire développementale des FE se fait en parallèle avec le développement du cortex préfrontal (Garon et al., 2008) qui est lui-même associé à plusieurs comportements parentaux (Bernier, Calkins, & Bell, 2016; Whittle et al., 2014). Il est donc important de tenir compte des facteurs environnementaux, tels que les comportements parentaux, dans l’étude des prédicteurs du développement des FE. Un nombre croissant d’études rapportent que les comportements parentaux contribueraient à l’explication des différences individuelles dans les FE (pour une recension des écrits, voir Fay-Stammbach, Hawes, & Meredith, 2014). Une récente méta-analyse regroupant 42 études suggère que la
relation entre le développement des FE à l’enfance et les comportements parentaux serait faible, mais significative (Valcan, Davis, & Pino-Pasternak, 2017; rm = .22). Cette relation
est significative peu importe les FE et les comportements parentaux ciblés. Les mécanismes qui expliquent cette relation demeurent toutefois méconnus.
Mécanismes génétiques ou environnementaux
Lorsqu'on tente d'identifier des facteurs propres aux parents, tels que les comportements parentaux, qui favorisent le développement de l’enfant, un des principaux défis est de déterminer dans quelle mesure les associations observées entre ces facteurs et le développement de l’enfant sont le reflet d’influences environnementales ou génétiques. Dans la littérature, la relation entre les comportements parentaux et les FE est généralement attribuée à des effets de l’environnement social. Toutefois, étant donné que les enfants et les parents partagent un bagage génétique (transmis à l’enfant par le parent), cette relation pourrait aussi être le reflet d’influences génétiques communes. Plusieurs études, qui seront détaillées plus bas, suggèrent d’ailleurs que les FE sont hautement héritables (Friedman et al., 2008; Friedman et al., 2016). En observant une association positive et significative entre les FE des parents et celles des enfants (Cuevas et al., 2014; Deater-Deckard, 2014), des études suggèrent aussi une transmission intergénérationnelle des FE. Des facteurs environnementaux tels que les comportements parentaux pourraient jouer un rôle dans cette transmission intergénérationnelle, de même que des facteurs génétiques transmis à l’enfant par le parent (Deater-Deckard, 2014). Ainsi, il serait crucial de départager les influences environnementales et génétiques qui sous-tendent la relation entre les FE de l’enfant et les comportements parentaux afin de mieux comprendre les mécanismes susceptibles d’expliquer cette relation.
Dans les écrits, il est généralement admis que le développement humain est à la fois influencé par des facteurs et mécanismes génétiques et environnementaux qui n'agissent pas indépendamment les uns des autres (Petrill, 2002). Plus précisément, on distingue généralement trois sources d’influences sur le développement humain, soit les influences génétiques, celles de l'environnement partagé et celles de l'environnement unique (Scarr,
1992). Les influences génétiques correspondent aux effets directs ou additifs d’un grand nombre de gènes qui opèrent ensemble (Friedman et al., 2008). L’environnement partagé regroupe les facteurs environnementaux auxquels les enfants d’une même fratrie sont exposés (par exemple, climat familial) et qui augmentent leur similarité (au-delà de leur similarité génétique). L’environnement unique regroupe les expériences et facteurs qui rendent les enfants d’une même famille différents, indépendamment de leur ressemblance génétique, et donc qui affectent différemment chaque enfant (p. ex., les conduites parentales différentielles). Les parents ne se comportent pas toujours de la même façon envers tous leurs enfants (Asbury, Dunn, Pike, & Plomin, 2003; Caspi et al., 2004; Forget-Dubois et al., 2007) et ces pratiques parentales différentielles pourraient entraîner des différences individuelles chez les enfants d’une même fratrie même si ceux-ci partagent un même bagage génétique (Boyle et al., 2004), et donc se traduire du côté de l’environnement unique. Ainsi, des divergences dans les pratiques parentales pourraient induire des différences de fonctionnement exécutif entre les enfants d’une même famille. Cela appuierait alors l’idée que les mécanismes qui sous-tendent la relation entre les FE de l’enfant et les comportements parentaux sont en partie de nature environnementale. Cela n’a toutefois jamais été examiné.
Des corrélations gènes-environnement (CorrGE) de type passive ou réactive pourraient aussi jouer un rôle dans l’explication de la relation entre les comportements parentaux et le développement des FE. Une CorrGE passive survient lorsque l'environnement parental est corrélé avec des facteurs génétiques partagés par le parent et l’enfant. Une CorrGE réactive pourrait aussi être à l’œuvre. Celle-ci se produit lorsque des caractéristiques héritables de l'enfant évoquent ou provoquent certaines réponses de la part de l’environnement (Scarr & McCartney, 1983). Une mère pourrait, par exemple, se montrer plus contrôlante envers un enfant qui est génétiquement prédisposé à des difficultés de régulation sur le plan comportemental. Seul un devis génétiquement informatif permettrait de départager les influences environnementales et génétiques susceptibles d’expliquer la relation documentée entre les comportementaux parentaux et le développement des FE de l’enfant.
Études de jumeaux
Les études de jumeaux offrent la possibilité de départager les contributions génétiques de celles qui sont environnementales (Plomin, DeFries, McClearn, & McGuffin, 2008; Plomin, DeFries, Knopik, & Neiderhiser, 2013). La logique est basée sur la comparaison de paires d’individus qui varient systématiquement quant à leur relation génétique, soit les jumeaux monozygotes (MZ) et dizygotes (DZ). Les jumeaux MZ ou identiques partagent 100 % de leur bagage génétique tandis que les jumeaux DZ ou fraternels partagent en moyenne 50 % de leur bagage génétique (DiLalla, Mullineaux, & Elam, 2009). Les devis de jumeaux permettent de déterminer dans quelle mesure les individus les plus semblables sur le plan génétique le sont aussi sur le plan phénotypique, peu importe le trait observable et mesurable. La quantification de la ressemblance entre jumeaux est donc centrale à cette approche. La ressemblance entre les jumeaux MZ et DZ sur un trait observé peut être quantifiée par des taux de concordance, soit par le calcul de corrélations intra-classes (Plomin et al., 2008). La comparaison des ressemblances entre les MZ et les DZ permet d’estimer dans quelle proportion les différences individuelles sur un trait sont liées à des facteurs génétiques et environnementaux. Si la ressemblance est plus grande pour les jumeaux MZ que DZ, cela indique que des contributions génétiques sont en cause. Si les jumeaux sont très semblables, qu’ils soient MZ ou DZ, cela indique que des facteurs environnementaux qui rendent semblables les enfants d’une même famille (issus de l’environnement partagé) sont en cause. Enfin, toute dissemblance entre jumeaux MZ est indicatrice d’une source de variation provenant de facteurs environnementaux propres à chaque enfant (issus de l’environnement unique) ou à l’erreur de mesure.
En génétique comportementale, l’héritabilité (h2 ou A) est le construit qui réfère à la proportion de variance phénotypique (par exemple, la variation entre individus au niveau de la performance à une tâche de FE) qui est attribuable à des facteurs génétiques. L’héritabilité permet de quantifier l’effet des facteurs génétiques sur les variations phénotypiques sans observer directement le génome. Dans une étude de jumeaux, l’héritabilité peut s’estimer grossièrement en soustrayant la ressemblance (i. e. corrélation ou concordance) entre les jumeaux DZ sur un trait/phénotype de la ressemblance entre les MZ sur ce même trait et en
doublant cette différence (DiLalla et al., 2009). Une approche génétiquement informative et l’utilisation de modélisation par équation structurelle permettent, en plus d’estimer l’héritabilité d’un phénotype, d’estimer la variance dans le phénotype qui peut s’expliquer par des facteurs issus de l’environnement partagé ou commun (c2 ou C) et par des facteurs issus de l’environnement unique (e2 ou E). La méthode des jumeaux peut être utilisée pour examiner les contributions environnementales et génétiques qui sous-tendent n’importe quel phénotype mesurable, incluant les FE et les comportements des parents. Dans ce mémoire, la méthode des jumeaux sera utilisée pour examiner l’étiologie génétique et environnementale des FE et des comportements parentaux de contrôle.
Étiologie des FE
Les études de jumeaux qui ont examiné l’héritabilité des FE durant l’enfance se font plutôt rares, mais sont de plus en plus nombreuses. Dresser un portrait cohérent des résultats de ces études est toutefois difficile, car plusieurs divergences méthodologiques les caractérisent. Les tâches utilisées pour mesurer les FE, de même que l’âge des enfants varient d’une étude à l’autre. Tandis que certaines études regroupent la performance des enfants dans plusieurs tâches qui mesurent différentes dimensions des FE, d’autres se sont penchées sur la performance à des tâches prises individuellement. Une recension des écrits récente (Li & Roberts, 2017) rapporte une héritabilité faible à modérée, variant de .09 à .64, pour les dimensions des FE mesurées séparément, mais une héritabilité plus élevée (autour de .99) pour un score factoriel regroupant la performance à plusieurs tâches qui mesurent différentes FE, ce qui souligne que les différentes FE ont une forte base génétique commune. Les auteurs présentent aux pages 107-109 de leur recension des écrits, un tableau synthèse des résultats des études réalisées dans ce domaine. Au total, sept études de jumeaux se sont intéressées à l’étiologie des FE mesurées par des tâches standardisées durant la période de l’enfance. Celles-ci seront détaillées ci-dessous.
Une seule étude a été réalisée durant l’âge préscolaire (Wang & Saudino, 2013). Cette étude porte sur l’étiologie de la performance à une tâche mesurant la mémoire de travail (Visually Cued Recall Task) dans un échantillon de 304 paires de jumeaux (140 MZ, 164 DZ)
âgés de 3 ans. Les résultats de celle-ci révèlent que 29% des variations individuelles dans la mémoire de travail s’expliquent par des facteurs génétiques (A), tandis que 32% s’expliquent par des facteurs issus de l’environnement partagé (C) et 39% par des facteurs issus de l’environnement unique (E).
Polderman et ses collaborateurs (2006) ont examiné l’étiologie de la mémoire de travail mesurée par les tâches neuropsychologiques d’Amsterdam au sein d’un échantillon de 237 paires de jumeaux (125 MZ, 112 DZ) évalués à deux reprises, soit à l’âge de 5 et 12 ans. Cette étude révèle qu’à 5 ans, 55% des différences individuelles dans la mémoire de travail s’expliquent par des facteurs génétiques (A) et que 45% s’expliquent par des facteurs de l’environnement unique (E). Tandis qu’à 12 ans, 73% des variations individuelles dans la mémoire de travail s’expliquent par des facteurs génétiques (A) et 27% s’expliquent par des facteurs de l’environnement unique (E). Les résultats de cette étude suggèrent que l’héritabilité de la mémoire de travail augmenterait au fil du temps. Toutefois, il est aussi possible que cette augmentation soit le résultat d’une diminution de l’erreur de mesure chez les enfants plus âgés.
Groot, De Sonneville, Stins et Boomsma (2004) ont examiné l’héritabilité de l’inhibition mesurée à l’aide d’une tâche standardisée (Go/no-go) à l’âge de 6 ans au sein d’un échantillon de 237 paires de jumeaux (125 MZ, 112 DZ). Deux modèles étiologiques différents ont été retenus sur la base d’un bon indice d’ajustement : le modèle AE et CE. L’absence de différence significative entre les deux modèles n’a pas permis aux auteurs de l’étude d’appuyer un modèle plus que l’autre. Les auteurs se sont intéressés de façon séparée aux garçons et aux filles, ce qui donne quatre modèles différents. Pour les filles, le modèle AE obtenu suggère que 36% des variations individuelles dans l’inhibition s’expliquent par des facteurs génétiques (A) et que 64% s’expliquent par des facteurs de l’environnement unique (E), alors que le second modèle CE suggère que 29% des variations individuelles dans l’inhibition s’expliquent par des facteurs de l’environnement partagé (C) et que 71% s’expliquent par des facteurs de l’environnement unique (E). Pour les garçons, le modèle AE obtenu dans l’étude suggère que 53% des variations individuelles dans l’inhibition s’expliquent par des facteurs génétiques (A) et que 57% s’expliquent par des facteurs de
l’environnement unique (E), alors que le second modèle CE suggère que 51% des variations individuelles dans l’inhibition s’expliquent par des facteurs de l’environnement partagé (C) et que 49% s’expliquent par des facteurs de l’environnement unique (E). Les résultats de cette étude appuient la présence d’influences issues de l’environnement unique, mais ne sont pas concluants en ce qui concerne les influences génétiques.
Wang, Deater-Deckard, Cutting, Thompson et Petrill (2012) ont utilisé un échantillon de 202 paires de jumeaux (90 MZ, 112 DZ) âgés entre 6 et 8 ans pour examiner l’étiologie de la mémoire de travail mesurée à l’aide d’une tâche administrée en laboratoire (Digit span
task). Les résultats de cette étude suggèrent que 21% de la variance dans la mémoire de travail
s’explique par des facteurs génétiques (A) tandis que 21% s’explique par des facteurs de l’environnement partagé (C) et 58% par des facteurs de l’environnement unique (E).
Kuntsi et ses collaborateurs (2006) examinent l’étiologie des FE entre 7 et 9 ans au sein d’un échantillon de 400 paires de jumeaux (156 MZ, 244 DZ). Les FE ont été mesurées à l’aide de deux tâches (Go/no-go task et Fast task) sollicitant l’inhibition et la mémoire de travail. Les résultats de cette étude suggèrent que 36% des variations individuelles dans la mémoire de travail s’expliquent par des facteurs génétiques (A) et que 64% s’expliquent par des facteurs de l’environnement unique (E). Tandis que 45% des variations individuelles dans l’inhibition s’expliquent par des facteurs génétiques (A) et que 55% s’expliquent par des facteurs de l’environnement unique (E).
L’étude de Schachar, Forget-Dubois, Dionne, Boivin et Robaey (2011) examine l’étiologie de l’inhibition mesurée à l’aide d’une tâche standardisée (Stop Signal) au sein d’un échantillon constitué de 131 paires de jumeaux (55 MZ, 76 DZ) âgés de 8 ans. Les résultats de cette étude suggèrent que 27% des variations individuelles dans l’inhibition s’expliquent par des facteurs génétiques (A) et que 73% s’expliquent par des facteurs de l’environnement unique (E).
Engelhardt, Briley, Mann, Harden et Tucker-Drob (2015) ont administré 12 tâches à 272 paires de jumeaux (84 MZ, 188 DZ) âgés entre 7 et 15 ans afin de mesurer trois
dimensions des FE, soit la mémoire de travail (Symmetry Span, Listening Recall et Digit Span
Back), l’inhibition (Animal Stroop, Mickey et Stop Signal) et la flexibilité cognitive (Trail Making, Plus Minus et Local Global). Les performances des enfants aux différentes tâches
ont été combinées afin de créer un score global de fonctionnement exécutif. Les résultats de cette étude suggèrent que des influences génétiques expliqueraient la totalité de la variance commune entre les différentes FE (A = 1.00; E = .00). Ainsi, les corrélations entre les FE sont entièrement attribuables à une base génétique commune. Par ailleurs, les résultats montrent que d’autres influences génétiques et environnementales sont uniques à chacune des FE et que l’étiologie des FE varie d’une dimension à l’autre et d’une tâche à l’autre. Même lorsque les tâches mesurent une même dimension du fonctionnement exécutif, l’étiologie des différences individuelles dans la performance à ces tâches varie. Les auteurs soulignent toutefois le peu de variance attribuable à des facteurs issus de l’environnement partagé : la variance dans les FE qui n’est pas expliquée par les gènes est majoritairement expliquée par l’environnement unique. Selon les résultats obtenus, l’héritabilité des différences individuelles dans la performance aux tâches mesurant la flexibilité cognitive varie entre 0 et 59 % et la variance résiduelle est majoritairement attribuable à des facteurs issus de l’environnement unique (E entre .15 et .95). En ce qui concerne la mémoire de travail, l’héritabilité dans la performance aux tâches varie entre 29 et 73% et la variance résiduelle est aussi majoritairement attribuable à des facteurs issus de l’environnement unique (E entre .39 et .64). En ce qui concerne l’inhibition, l’héritabilité des différences individuelles dans la performance aux tâches varie entre 27 et 55% et l’influence de l’environnement unique varie de .55 à .77.
En résumé, les estimations d’héritabilité varient d’une étude à l’autre notamment en fonction des FE examinées et des tâches utilisées pour les mesurer. Les études réalisées dans le domaine suggèrent essentiellement que les associations entre les différentes FE (variance commune) s’expliquent par des mécanismes génétiques (Engelhardt et al., 2015), tandis que la variance dans les FE prises individuellement s’expliquerait par des facteurs génétiques et des facteurs issus de l’environnement unique. L’environnement partagé jouerait un rôle minime dans l’explication des différences individuelles dans les FE. Selon les études réalisées, l’héritabilité de la mémoire de travail varierait entre 21% et 55% et celle de
l’inhibition entre 27% et 53%. En ce qui concerne l’héritabilité de la flexibilité cognitive, celle-ci a fait l’objet d’une seule étude qui suggère qu’elle varie entre 0% et 59% dépendamment de la tâche utilisée pour la mesurer (Engelhardt et al., 2015). Cette étude a mesuré la flexibilité auprès d’enfants âgés entre 7 et 15 ans. L’héritabilité de la flexibilité cognitive avant l’âge de 7 ans demeure ainsi inconnue. Elle sera examinée pour la première fois dans le cadre de ce mémoire. Peu d’études se sont d’ailleurs penchées sur l’héritabilité des FE durant la période préscolaire même si cette période est caractérisée par un développement rapide dans ces habiletés (Diamond, 2016). La seule étude réalisée chez les enfants avant l’entrée à l’école suggère que la variance dans les FE qui n’est pas expliquée par la génétique est essentiellement expliquée par l’environnement unique (Wang & Saudino, 2013). Ainsi, des facteurs issus de l’environnement unique tels que les comportements différentiels des parents envers chacun de leur enfant pourraient expliquer une portion importante de la variance dans la performance des enfants à différentes tâches de FE durant l’enfance. Cela souligne la pertinence de poursuivre les efforts pour identifier ces facteurs.
Étiologie des comportements parentaux
Selon une première méta-analyse réalisée sur 12 études ayant examiné l’étiologie des comportements parentaux, ceux-ci seraient légèrement héritables (h2 = 27%) (Kendler &
Baker, 2007). Selon une méta-analyse plus récente regroupant 32 études (Avinun & Knafo, 2014), 23% des variations individuelles dans les comportements parentaux s’expliqueraient par des facteurs génétiques de l’enfant (A), 43% par des facteurs issus de l’environnement partagé (C) et 34% par des facteurs de l’environnement unique (E). Ainsi, il est possible que certains gènes de l’enfant (possiblement impliqués dans un mécanisme de corrélation GE) permettent d’expliquer la relation documentée entre les comportements parentaux et les FE. Toutefois, cela n’a jamais été examiné. Bien que certaines études génétiquement informatives aient traité les FE et les comportements parentaux séparément et démontrent qu’une portion de la variance dans ces phénotypes s’explique par des facteurs génétiques, les contributions sous-jacentes à leur relation restent encore un mystère.
Modèles génétiques bivariés
Les modèles univariés ne permettent pas de déterminer si des processus génétiques sont à l’origine de la relation entre les FE et les comportements parentaux. Pour clarifier ces processus, l’utilisation d’un modèle d’analyse génétique bivarié est nécessaire. L’approche génétiquement informative bivariée est particulièrement pertinente lorsqu’il s’agit d’examiner les mécanismes qui sous-tendent la relation entre les comportements parentaux et le développement de l’enfant (Deater-Deckard, 2000). Celle-ci permet de documenter les facteurs génétiques susceptibles de sous-tendre cette relation afin notamment de confirmer ou d’infirmer la contribution environnementale des pratiques parentales dans l’explication des différences individuelles observées chez les enfants. Cette approche peut être utilisée afin de déterminer si l’effet des comportements parentaux opère par le biais de mécanismes environnementaux qui sont distincts des mécanismes génétiques qui sont susceptibles d’être aussi à l’œuvre.
Les modèles génétiques bivariés permettent l’estimation des sources environnementales et génétiques de covariance entre deux phénotypes (Pike, McGuire, Hetherington, Reiss, & Plomin, 1996; Plomin et al., 2013). Ils sont essentiellement basés sur une décomposition des corrélations croisées dites « cross-trait, cross-twin » (CTCT). Par exemple, la corrélation entre les FE du jumeau 1 et le comportement du parent envers le jumeau 2. Ils permettent de dire dans quelle mesure la covariance entre deux phénotypes (par exemple, les FE de l’enfant et les comportements parentaux) s’explique par des facteurs génétiques que les jumeaux ont en commun (plus grande corrélation CTCT des jumeaux MZ que DZ; il s’agit d’une corrélation génétique) ou par des facteurs environnementaux en commun (corrélations similaires non nulles CTCT des jumeaux MZ et DZ) ou environnementaux uniques (corrélations CTCT des MZ qui dévient de 1) (Posthuma, 2009). Cette approche permet essentiellement de vérifier la présence d’une corrélation gènes-environnement. Par exemple, si les parents agissent de façon plus similaire envers les jumeaux MZ que les jumeaux DZ (i.e., lorsque les enfants sont plus similaires génétiquement), cela suggère que le bagage génétique de l’enfant influence le comportement
du parent (corrélation gène-environnement) par le biais de son association avec un phénotype de l’enfant.
Plusieurs études ont utilisé une approche génétiquement informative pour examiner les mécanismes qui sous-tendent la relation entre les comportements parentaux et certaines sphères du développement de l’enfant (Boivin et al., 2005; Deater-Deckard, 2000, 2014; Forget-Dubois et al., 2007; Guimond et al., 2012; Micalizzi, Wang, & Saudino, 2017; Muftić & Updegrove, 2017). Plusieurs de ces études suggèrent que les comportements parentaux opèrent par le biais de mécanismes environnementaux et de CorrGE pour influencer le développement de l’enfant. Aucune étude génétique ne s’est toutefois intéressée à la relation entre les comportements parentaux et les FE. Un devis génétiquement informatif de l’enfant sera donc utilisé dans le cadre de ce mémoire pour mieux comprendre les mécanismes qui sous-tendent la relation entre le contrôle parental et le développement des FE.
Le contrôle parental et le développement des FE
Tel que mentionné précédemment, de plus en plus d’études s’intéressent à la relation entre les comportements parentaux et le développement des FE (Distefano, Galinsky, McClelland, Zelazo, & Carlson, 2018; Hughes & Devine, 2019; Lee, Baker, & Whitebread, 2018; Zeytinoglu, Calkins, & Leerkes, 2019). Malgré cet intérêt grandissant, la majorité des études dans le domaine se sont penchées sur les comportements parentaux positifs visant à soutenir le développement des FE de l’enfant tels que la sensibilité, le soutien à l’autonomie, l’étayage et la stimulation (Blair et al., 2011; Clark et al., 2013; Distefano et al., 2018; Hughes & Ensor, 2009; Matte-Gagne & Bernier, 2011), en négligeant les comportements négatifs susceptibles de nuire au développement des FE, tels que le contrôle parental (Valcan et al., 2017). Parmi les études recensées dans la méta-analyse réalisée par Valcan et ses collègues, 15 études portaient sur les pratiques parentales dites négatives (échantillon total de 4322 dyades parent-enfant). Les pratiques parentales négatives regroupaient les comportements intrusifs ou sévères (intrusive or harsh parenting), l’expression d’émotions négatives envers l’enfant, le détachement, l’hostilité, le contrôle, les comportements autoritaires visant à prendre le pouvoir (power assertion) et le rejet. Les résultats obtenus dans le cadre de cette
aux FE (r = -.22, p <.01) entre l’âge de 18 mois et de 8 ans. Parmi les comportements négatifs qui ont été démontrés comme étant significativement associés aux FE, on retrouve le contrôle parental (Hertyas, Hendrawan, Arbiyah, & Nurbatari, 2018; Hughes & Devine, 2019; Meuwissen & Carlson, 2015; Roskam, Stieyenart, Meunier, & Noël, 2014).
Le contrôle parental a été conceptualisé de différentes façons dans les écrits scientifiques et au sein même des études portant sur les relations entre ce comportement et le développement des FE. Selon la théorie de l’autodétermination (TDA), le contrôle parental regroupe l’ensemble des techniques éducatives qui exercent une pression sur l’enfant pour qu’il pense, se sente ou se comporte d’une façon qui va à l’encontre de ses besoins psychologiques (Grolnick & Ryan, 1989). Le contrôle parental peut prendre différentes formes et se manifester par des conduites parentales intrusives ou contraignantes (p. ex., empêcher l’enfant de s’exprimer, dire à l’enfant quoi faire durant une période de jeux, interférer avec ses activités et brimer son autonomie) et coercitives ou punitives (p. ex., des critiques, l’utilisation de la force et de punitions physiques excessives). Il peut aussi prendre une forme plus implicite et psychologique telle qu’insister pour que son enfant se dégêne ou pour qu’il aille de l’avant et fonce alors qu’il hésite ou lui manifester de l’indifférence ou menacer de lui retirer de l’affection. Lorsque le parent fait preuve de contrôle (psychologique, coercitif ou intrusif), le développement des capacités de régulation de l’enfant serait entravé (Grolnick & Pomerantz, 2009). En effet, l’enfant aurait moins d’occasions de développer des stratégies lui permettant d’apprendre à contrôler par lui-même ses pensées, émotions et comportements et cela pourrait engendrer des déficits sur le plan des FE (Roskam et al., 2014). Plusieurs études appuient d’ailleurs l’existence d’une relation négative entre les comportements parentaux de contrôle, tels que la coercition, la punition et l’intrusion, et des construits s’apparentant aux FE tel que l’effortful control (Karreman, van Tujil, van Aken, & Deković, 2009; Olson, Chloe, & Sameroff, 2017; Tiberio et al., 2016). Le contrôle parental intrusif, punitif ou coercitif est d’ailleurs considéré comme étant un facteur de risque important dans le développement de l’enfant (Bindman, Hindman, Bowles, & Morrison, 2013; Pomerantz et Wang, 2009). Il est notamment associé à des troubles de comportement (Pinquart, 2017), des difficultés d’autorégulation (Bridgett et al., 2018), de l’anxiété (Van der Bruggen, Stams, & Bögels, 2008), ainsi qu’à des difficultés académiques (Pinquart,
2016). Peu d’études ont toutefois examiné la relation entre le contrôle parental (peu importe sa forme) et le développement des FE (Roskam et al., 2014).
Dans l’étude de Roskam et ses collègues (2014), la capacité d’inhibition a été mesurée à trois reprises entre l’âge de 2 et 8 ans auprès d’un échantillon de 421 enfants à l’aide de six tâches standardisées. La performance des enfants aux six tâches a ensuite été regroupée pour former un score global de capacité d’inhibition. Afin de mesurer le contrôle, les parents (348 mères et 342 pères) devaient indiquer sur une échelle Likert en 5 points allant de « Jamais » à « Toujours », à quel point ils avaient tendance à punir (physiquement ou non) l’enfant lorsqu’il se comportait mal (contrôle punitif) et à l’ignorer (contrôle psychologique) jusqu’à ce qu’il se comporte mieux. Les résultats de l’étude suggèrent une relation négative entre le contrôle parental de la part de la mère et l’inhibition (r = -.18, p < 0.01).
Meuwissen et Carlson (2015) ont observé et mesuré le contrôle parental de 110 pères en laboratoire lorsque l’enfant était âgé de 3 ans. Le père et l’enfant devaient compléter un casse-tête trop difficile pour l’enfant. Le père était considéré contrôlant lorsqu’il intervenait trop tôt ou excessivement, utilisait un ton sévère ou ne tolérait pas que l’enfant dévie de la tâche. Les FE ont été mesurées durant la même rencontre à l’aide de quatre tâches impliquant la mémoire de travail, l’inhibition et la flexibilité cognitive. Un score global de FE a été créé en combinant les scores de performance obtenus aux tâches. Les résultats de cette étude indiquent une relation négative et modérée entre le contrôle paternel et les FE (r = -.30, p <.01).
Dans l’étude de Hertyas et ses collaborateurs (2018), le contrôle a été mesuré en laboratoire auprès de 118 dyades parent-enfant lorsque l’enfant était âgé entre 48 et 72 mois. Le parent et l’enfant devaient compléter deux tâches difficiles pour l’enfant, soit répliquer un modèle à l’aide de blocs et compléter un casse-tête. Le parent était considéré contrôlant lorsqu’il intervenait trop tôt ou excessivement, utilisait un ton sévère ou ne tolérait pas que l’enfant dévie de la tâche. Trois tâches de FE ont été administrées afin de mesurer l’inhibition (Grass/Snow task), la mémoire de travail (Backward Span task) et la flexibilité cognitive (Dimensional Card Sorting task) pour ensuite créer un score global des FE. Les résultats de
cette étude révèlent une relation négative et modérée entre le contrôle maternel et les FE (r = -0.33, p < 0.05).
Certaines études se sont aussi intéressées à la relation entre les FE de l’enfant et les comportements intrusifs des parents (Blair et al., 2011; Cuevas et al., 2014). Dans l’étude de Blair et al. (2011), les FE ont été mesurées à l’âge de 36 mois auprès d’un échantillon de 1292 enfants à l’aide de trois tâches standardisées mesurant la flexibilité cognitive, la mémoire de travail et l’inhibition. Un score global de FE a été créé en combinant les scores de performance obtenus à toutes les tâches. L’intrusion maternelle était mesurée de façon observationnelle à 7, 15 et 24 mois et définie comme la tendance de la mère à interférer avec les tentatives de résolution de problèmes de l’enfant en intervenant plus que nécessaire pour que l’enfant réussisse. Les résultats de cette étude indiquent une relation négative et significative entre les FE et l’intrusion maternelle à 7, 15 et 24 mois (r = -.21, -.21, -.25; p <.01).
Dans l’étude de Cuevas et ses collaborateurs (2014), les FE de 63 enfants ont été mesurées à l’âge de 24, 36 et 48 mois à l’aide de sept tâches impliquant l’inhibition, la mémoire de travail et la flexibilité cognitive. Un score composite a été créé pour les FE en regroupant tous les temps de mesure et toutes les tâches. Les comportements intrusifs de la mère ont été mesurés de façon observationnelle à 10, 24 et 36 mois. L’intrusion de la mère était caractérisée par des comportements tels que l’expression d’émotions négatives, la recherche excessive de l’attention de l’enfant durant le jeu ou le fait de forcer l’enfant à jouer avec un jouet en particulier. Un score composite a été créé en combinant les trois temps de mesure de l’intrusion maternelle. Les résultats de cette étude suggèrent une relation négative entre l’intrusion maternelle et les FE (r = -.35, p <.01).
Même si la mesure et l’opérationnalisation du contrôle parental et des FE varient d’une étude à l’autre, les résultats des études réalisées dans ce domaine suggèrent que des déficits dans les FE seraient plus probables chez les enfants dont les parents adoptent des comportements contrôlants ou intrusifs. Bien qu’un nombre grandissant d’études appuient l’existence d’une association entre le contrôle parental et le développement des FE, les
mécanismes qui sous-tendent cette association demeurent méconnus, notamment le rôle d’une vulnérabilité génétique associée aux comportements chez les enfants qui pourraient influencer les conduites parentales.
Objectifs
À l’aide d’un devis génétiquement informatif, l’objectif de ce mémoire est de mieux comprendre les mécanismes génétiques et environnementaux chez l’enfant qui sous-tendent la relation entre le contrôle maternel et la flexibilité cognitive de l’enfant, une composante importante du fonctionnement exécutif. Dans un premier temps, l’étiologie de la flexibilité cognitive sera examinée à l’aide d’un modèle génétique univarié. En s’appuyant sur les études de jumeaux antérieures (Li & Roberts, 2017), une contribution significative, mais faible des facteurs génétiques, est attendue pour la flexibilité cognitive. Il est aussi attendu que la variance dans la flexibilité cognitive non expliquée par les facteurs génétiques soit essentiellement expliquée par des facteurs issus de l’environnement unique.
Dans un deuxième temps, la relation entre le contrôle maternel et la flexibilité cognitive de l’enfant sera examinée. Un modèle génétique bivarié sera utilisé afin de départager les mécanismes environnementaux ou génétiques qui sous-tendent cette relation. Comme aucune étude génétiquement informative n’a examiné la relation entre le contrôle parental et la flexibilité cognitive, il est difficile de formuler une hypothèse. Toutefois, sur la base des études génétiques antérieures réalisées sur d’autres sphères du développement de l’enfant, il est attendu que des mécanismes environnementaux soient essentiellement à l’origine de cette relation.
Chapitre 1 : Méthodologie
Participants et procédure
Les participants de cette étude proviennent de l’Étude des jumeaux nouveau-nés du Québec (EJNQ; Boivin et al., 2012). Il s’agit d’une vaste étude longitudinale menée auprès d’un échantillon de 1324 jumeaux nés entre 1995 et 1998 dans la grande région de Montréal. Le recrutement a été effectué via le registre des naissances du Bureau de la Statistique du Québec. Tous les parents de jumeaux nés sans complications majeures dans la grande région de Montréal (N = 989) ont été contactés soit par téléphone ou par lettre et 75% d’entre eux ont accepté de participer à l’étude. Les jumeaux et leurs parents ont d’abord participé à trois rencontres et entrevues à domicile lorsque les enfants étaient âgés de 5, 18 et 30 mois et ils ont ensuite été suivis une fois par année. À chacune des rencontres, les jumeaux, accompagnés de leur mère, ont été évalués individuellement par un assistant de recherche pour une période d’environ 20 minutes. Pendant que les jumeaux complétaient à tour de rôle des tâches mesurant différentes habiletés, leur mère complétait des questionnaires mesurant différents aspects du développement de l’enfant et de l’environnement familial. À la fin de la rencontre, la mère recevait aussi un questionnaire à compléter et retourner par la poste dans le mois suivant la rencontre.
L’échantillon de la présente étude est composé de 934 jumeaux (400 MZ et 534 DZ) ayant participé à la rencontre réalisée en laboratoire lorsque les enfants étaient âgés de 60 mois (M = 63.46; ET = 3.30). Le poids moyen de l’enfant à la naissance est de 2.47 kg (ET = .54). L’âge moyen de la mère à la naissance de l’enfant est de 30 ans (ET = 4.80). 49% des jumeaux sont des garçons. Le revenu familial annuel moyen se situe entre 30 000 et 40 000$ : 7% des familles ont un revenu inférieur à 20 000$, 7% entre 20 000 et 30 000$, 10% entre 30 000 et 40 000$, 23% entre 40 000 et 60 000$ et 53% ont un revenu de plus de 60 000$. Pour ce qui est de la scolarité maternelle, 14% des mères n’ont aucun diplôme, 54% ont un diplôme d’études secondaires ou collégiales et 32% des mères détiennent un diplôme universitaire.
Mesures
Contrôle maternel. Afin d’évaluer le contrôle maternel, un questionnaire
autorapporté (Boivin et al., 2005) a été administré à la mère séparément pour chaque jumeau lorsque les enfants étaient âgés de 60 mois. Cinq items ont été sélectionnés comme indicateurs du contrôle maternel (voir Annexe A pour une liste de ces items). Pour chaque item, la mère devait indiquer sur une échelle variant de 1 (jamais) à 5 (tout le temps) à quelle fréquence elle avait tendance à adopter les comportements décrits. Par la suite, chaque fréquence a été additionnée pour former un score qui reflète le degré de contrôle parental de la mère. Un score plus élevé indique un degré plus élevé de contrôle maternel. La cohérence interne du construit mesuré à l’aide des cinq items sélectionnés est acceptable (ɑ = 0,64). Lors de la rencontre effectuée en laboratoire, la mère devait compléter la première moitié du questionnaire pour le premier jumeau et la deuxième moitié pour le second jumeau. Quelques semaines plus tard, la mère devait terminer la complétion des deux autres moitiés du questionnaire à la maison et les renvoyer par la poste à l’équipe de recherche (Boivin et al., 2005). Cette complétion à des temps séparés a pour but d’éviter certains biais liés au fait de remplir deux fois le même questionnaire de façon rapprochée, ce qui peut entraîner notamment de l’ennui ou un effet de contagion (les réponses au premier questionnaire affectant les réponses au deuxième questionnaire qui concerne un enfant différent).
Flexibilité cognitive. Pour mesurer les FE, une version française du Dimensional Change Card Sort (DCCS) a été administrée en laboratoire à chaque jumeau à l’âge de 60
mois (Zelazo, 2006). Le Tri de cartes consiste à trier des cartes par couleur ou par forme en fonction d’une règle qui change. Il comporte 59 essais, dont 18 essais de pratique et 1 essai de démonstration. Le score total varie de 0 à 40 et correspond au nombre de cartes correctement classées par l’enfant lors des 40 essais expérimentaux. Le Tri de cartes est considéré comme l’une des tâches expérimentales les plus fiables et valides pour mesurer les différences individuelles sur le plan des FE à l’âge préscolaire et scolaire (Bauer & Zelazo, 2014; Carlson, 2005; Zelazo, 2006, 2015). Cette tâche a d’ailleurs été utilisée dans plus de 150 études pour mesurer les FE durant l’enfance et elle est démontrée comme étant
particulièrement sensible au développement des FE entre l’âge de 2 et 7 ans (Doebel & Zelazo, 2015; Carlson & Zelazo, 2014). De plus, différentes versions plus exigeantes ont aussi été utilisées chez des enfants plus âgés, de même que chez des adolescents et des adultes (Diamond & Kirkham, 2005; Morton, Bosma, & Ansari, 2009; Zelazo, Craik, & Booth, 2004; Zelazo, 2006). Une méta-analyse portant sur le DCCS durant l’enfance suggère une amélioration significative de la performance à cette tâche entre l’âge de 3 et 5 ans (Doebel & Zelazo, 2015) ce qui soulève la pertinence d’utiliser cet outil à 5 ans pour mesurer les différences individuelles dans les FE. Le DCCS possède une bonne validité convergente avec d’autres mesures des FE (Zelazo, 2006) et la performance dans cette tâche est associée de façon cohérente à d’autres sphères du développement cognitif de l’enfant telles que le langage (Carlson & Meltzoff, 2008), la théorie de l’esprit (Kloo & Perner, 2003; Carlson & Moses, 2001) et le développement fonctionnel du cortex préfrontal latéral (Ezekiel, Bosma, & Morton, 2013).
Le Tri de cartes comporte deux blocs, soit un bloc de pratique et un bloc expérimental. Le bloc de pratique comporte 18 essais. L’assistant de recherche présente à l’enfant six cartes sur lesquelles sont représentés des bateaux ou des fleurs de couleur bleue ou verte. L’assistant explique à l’enfant qu’il doit d’abord classer les cartes par forme : les fleurs avec les fleurs et les bateaux avec les bateaux. L’enfant s’exécute et l’assistant de recherche le corrige au besoin. Lorsque les six cartes sont classées correctement par forme, l’assistant de recherche modifie la règle et demande à l’enfant de classer les mêmes cartes par couleur : les cartes bleues avec les cartes bleues et les cartes vertes avec les cartes vertes. Par la suite, une composante supplémentaire, soit la présence ou l’absence d’un point noir situé sur la carte, est introduite. L’enfant reçoit la consigne de classer les cartes par couleur lorsqu’un point noir est présent sur les cartes et de les classer par forme lorsqu’il n’y a pas de point noir. Deux essais de pratique sont ensuite effectués par l’assistant de recherche. Ce dernier répète les consignes et invite l’enfant à classer quatre cartes pour se familiariser avec la tâche à effectuer, tout en corrigeant les erreurs de l’enfant. Enfin, le bloc expérimental de 40 essais débute. Pour tous les essais qui sont comptabilisés, l’assistant de recherche rappelle à l’enfant de classer les cartes par couleur s’il y a un point et de les classer par forme s’il n’y a pas de
point. À chaque carte, l’assistant de recherche mentionne si un point est présent ou non sur la carte.
Plan d’analyse
Les analyses préliminaires ont été réalisées à l’aide du logiciel SPSS (Version 24.0). Les corrélations intraclasses et les modèles génétiques univariés et bivariés ont été estimés à l’aide du logiciel Mplus 5.21 (Muthén & Muthén, 2007). Un modèle univarié ACE a d’abord été testé pour chaque phénotype, soit le contrôle maternel et la flexibilité cognitive. La variance du contrôle maternel et celle de la flexibilité cognitive ont été décomposées en trois variables latentes, représentant les sources d’influences génétiques (A), de l’environnement partagé (C) et de l’environnement unique (E). Le modèle ACE est ajusté aux données en un modèle à deux groupes où la covariance génétique est fixée à 1.0 pour les MZ et à 0.5 pour les DZ. Les covariances de l’environnement commun (facteurs C) sont fixées à 1.0 pour les jumeaux MZ et DZ, tandis que les covariances attribuables à l’environnement unique (facteurs E) sont fixées à 0. Le modèle ACE complet a ensuite été comparé à d’autres modèles génétiques univariés plus simples afin de déterminer si ceux-ci représentaient mieux les données (le modèle AE (C = 0), le modèle CE (A = 0) et le modèle E). Cette démarche itérative permet de déterminer si on peut retirer un paramètre sans détériorer de façon significative l’ajustement du modèle. Le modèle le plus parcimonieux a été retenu sur la base du khi carré (différence non significative) et du Akaike’s information criterion (AIC) le plus bas.
Un modèle génétique bivarié a ensuite été estimé pour examiner l’association entre le contrôle maternel et la flexibilité cognitive et décomposer les contributions génétiques et environnementales qui la sous-tendent. Les sources partagées de variance entre le contrôle maternel et les fonctions exécutives ont été estimées sous forme de corrélations entre les facteurs génétiques (RG), de l’environnement commun (RC) et de l’environnement unique
(RE). Les modèles nichés supprimant le A ou le C de chacun des phénotypes ou supprimant
les associations entre les A, C ou E des deux phénotypes ont également été testés et comparés avec le modèle complet afin de conserver le modèle le plus parcimonieux. Il est à noter que
des covariables ont été ajoutées aux modèles telles que le sexe, le niveau d’éducation des mères, le statut socioéconomique et le poids à la naissance de l’enfant. Par souci de parcimonie, étant donné que les paramètres estimés étaient similaires, les modèles sans variable confondante ont été retenus.
Chapitre 2 : Résultats
Analyses préliminaires
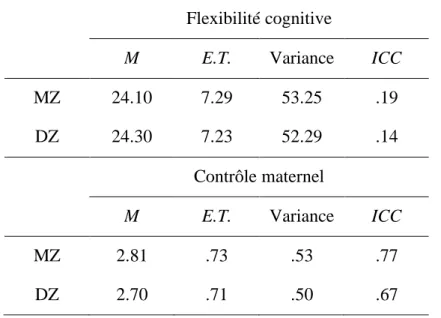
Des analyses descriptives ont d’abord été effectuées sur les variables d’intérêt et elles sont présentées dans le Tableau 1. Celles-ci démontrent notamment que les moyennes et variances de la flexibilité cognitive et du contrôle maternel des jumeaux MZ et DZ sont similaires. Un test d’égalité des variances a été effectué et révèle que celles-ci ne se distinguent pas de façon significative. Ainsi, aucune transformation n’a été nécessaire afin de pouvoir estimer les modèles génétiques. Les corrélations effectuées dans l’échantillon total révèlent que le contrôle maternel est faiblement, mais significativement et négativement associé à la flexibilité cognitive de l’enfant (r = -.13, p < 0.001). Cela justifie de poursuivre les analyses.
Modèles génétiques univariés
Un modèle univarié ACE a été testé pour chaque phénotype, soit le contrôle maternel et la flexibilité cognitive. Des corrélations intra-paires ont d’abord été effectuées pour les jumeaux MZ et DZ séparément afin d’avoir un aperçu de l’héritabilité de ces variables. Les résultats de ces analyses démontrent une corrélation intra-paire pour les jumeaux MZ légèrement supérieure (r = 0.77) à celle des jumeaux DZ (r = 0.67) pour le contrôle maternel. Cela suggère la présence d’influences génétiques chez l’enfant (gènes des jumeaux). Les résultats démontrent par ailleurs une corrélation intra-paire similaire pour les jumeaux MZ (r = 0.19) et DZ (r = 0.14) en ce qui concerne la flexibilité cognitive. Cela suggère l’absence de contribution génétique et l’intervention prédominante de facteurs environnementaux dans l’explication des différences individuelles dans la flexibilité cognitive.
Les résultats des analyses génétiques univariées effectuées pour la flexibilité cognitive sont présentés dans le Tableau 2. Le modèle ACE testé (2 (6) = 3.28, p = .77) suggère que la variance expliquée par les facteurs génétiques n’est pas significativement différente de 0.
Les chi-carrés des modèles testés suggèrent que le modèle CE est le plus parcimonieux (2 (7) = 3.15, p = .87). Ce modèle suggère que la contribution de l’environnement partagé à l’explication de la variance dans la flexibilité cognitive est significative et de 18%, tandis que la contribution de l’environnement unique est de 82%. Le chi-carré du modèle E (2 (8) =
13.76, p = .09) suggère qu’il vaut mieux retenir le modèle contenant plus de paramètres, soit le modèle CE. Ce modèle a donc été retenu dans les analyses multivariées subséquentes. La comparaison des différents modèles génétiques univariés réalisés sur le contrôle maternel a révélé que le modèle ACE était préférable (2 (6) = .01, p = 1.00). Ce modèle est présenté
dans le Tableau 2. Ce dernier indique que les facteurs génétiques (A) expliquent 21% de la variance dans le contrôle maternel, tandis que la contribution de l’environnement partagé est de 56% et celle de l’environnement unique est de 23%. Tous les paramètres du modèle ACE sont significatifs et donc retenus dans les analyses subséquentes.
Modèles génétiques multivariés
Les résultats des analyses génétiques multivariées sont présentés dans le Tableau 3 et illustrés dans la Figure 2. Le modèle bivarié ACE complet (2 (17) = 4.69, p = .99) a d’abord
été testé, mais rejeté. Le modèle complet ne s’ajustait pas mieux aux données que le modèle excluant l’étiologie génétique (facteur A) de la flexibilité cognitive (2 (19) = 4.70, ns). Le
pourcentage de variance dans la flexibilité cognitive expliqué par la génétique était non significatif (1%, ns) et la corrélation génétique entre la flexibilité et le contrôle maternel était non significative (Ra = 0.02, ns). Le modèle CE a donc été retenu pour la flexibilité cognitive, excluant, par le fait même, la covariance génétique entre le contrôle maternel et la flexibilité cognitive de l’enfant. Le modèle final retenu suggère donc que la corrélation phénotypique entre le contrôle maternel et la flexibilité cognitive de l’enfant ne s’explique pas par des facteurs génétiques communs. Les paramètres estimés du modèle retenu suggèrent aussi que cette corrélation ne s’explique pas par des facteurs issus de l’environnement partagé bien que la valeur du p approche le seuil de signification (Rc = -.21, p = .08). Les résultats suggèrent toutefois une corrélation de l’environnement unique significative (Re = -0.13, p < .05). Bien que faible, cette corrélation Re indique qu’une portion majeure de la corrélation phénotypique entre le contrôle et la flexibilité s’explique par des facteurs issus de l’environnement unique
(la corrélation phénotypique étant de -.13 dans l’échantillon global). Cela suggère que les associations entre le contrôle maternel et la flexibilité cognitive de l’enfant s’opèrent par des mécanismes environnementaux qui rendent les enfants d’une même fratrie différents.
Des analyses complémentaires de régression linéaire multiple ont ensuite été effectuées en utilisant la méthode de discordance entre jumeaux MZ. Les résultats de ces analyses sont présentés dans le Tableau 4. Des covariables pertinentes ont d’abord été entrées dans le premier bloc de la régression en tant que prédicteurs des discordances entre jumeaux MZ sur le plan de la flexibilité cognitive, soit le nombre d’années de scolarité et le revenu de la mère, le nombre de semaines de gestation de l’enfant, son âge exact lors de la passation du Tri de cartes et son poids à la naissance. Les scores de discordance sur le plan du contrôle maternel ont ensuite été ajoutés dans le second bloc de la régression pour prédire les scores de discordance sur le plan de la flexibilité cognitive. Les résultats révèlent que le contrôle maternel permet de prédire 2% de la variance dans la flexibilité cognitive à 60 mois (B = -0.16, p < .05), en contrôlant pour la génétique et l’environnement partagé à l’aide de la méthode de discordance entre jumeaux MZ. Ainsi, les jumeaux de 60 mois dont les mères se perçoivent comme étant plus contrôlantes performent moins bien à la tâche du Tri de cartes mesurant la flexibilité cognitive.
Chapitre 3 : Discussion
L’objectif du mémoire était de mieux comprendre les mécanismes génétiques et environnementaux qui sous-tendent la relation entre la flexibilité cognitive et le contrôle maternel durant l’âge préscolaire en utilisant un devis génétiquement informatif. L’étiologie de la flexibilité cognitive a d’abord été examinée à l’aide d’un modèle génétique univarié. Ensuite, la relation entre le contrôle maternel et la flexibilité cognitive a été examinée. Les contributions génétiques et environnementales sous-jacentes à cette relation ont finalement été départagées à l’aide d’un modèle génétique bivarié.
Interprétation des résultats
Étiologie de la flexibilité cognitive. Les résultats du modèle génétique univarié retenu
pour la flexibilité cognitive indiquent que l’héritabilité de la flexibilité cognitive est nulle et que les différences individuelles au niveau de la flexibilité cognitive s’expliquent par des facteurs issus de l’environnement (incluant l’erreur de mesure). Les résultats montrent plus spécifiquement une prédominance de l’environnement unique (82%), incluant l’erreur de mesure, comme source de variance avec une légère contribution de l’environnement partagé (18%). Cette étude est la première à examiner l’étiologie de la flexibilité cognitive durant l’âge préscolaire. Les résultats obtenus sont cohérents avec ceux de la seule autre étude ayant examiné l’étiologie de la flexibilité cognitive durant l’enfance, mais auprès d’enfants plus âgés, soit âgés entre 7 et 15 ans (Engelhardt et al., 2015). Bien que l’héritabilité de la flexibilité cognitive semble varier en fonction de la tâche utilisée pour la mesurer (Engelhardt et al., 2015), la contribution des facteurs génétiques demeure plutôt faible par rapport à la contribution de l’environnement. Ainsi, les mécanismes qui sous-tendent les différences individuelles dans la flexibilité cognitive seraient essentiellement environnementaux, mais les études faites à ce jour sont insuffisantes pour en arriver à une conclusion. Ce constat est cohérent avec ceux d’études réalisées durant l’adolescence et à l’âge adulte (Taylor, 2007; Chou, Kuo, Lin, & Chen, 2010). En effet, les études de jumeaux qui ont utilisé la version adulte du DCCS (le Wisconsin Card Sorting Test; WCST) suggèrent que la performance à
cette tâche ne serait pas sujette à des influences d’ordre génétique (Taylor, 2007; Chou et al., 2010). Les corrélations MZ pour la majorité des scores obtenus au WCST sont près de zéro au sein d’une population adulte (Taylor, 2007). Quant à la performance au WCST au sein d’une population adolescente, la variance attribuable aux gènes est presque inexistante tandis que celle attribuable aux facteurs issus de l’environnement unique est prédominante (A = .07, C = .29, E = .65; Chou et al., 2010). Les différences individuelles dans la flexibilité cognitive ne semblent donc pas s’expliquer par de la variance génétique, peu importe le stade de développement.
Même si les résultats des différentes études convergent, l’absence de variance génétique peut soulever des questions, notamment à la lumière des études qui démontrent que certaines FE, telles que la mémoire de travail, sont modérément ou hautement héritables (Li & Roberts, 2017). Les études qui se sont intéressées au point d’émergence des différentes FE suggèrent que la flexibilité cognitive émergerait plus tard dans le développement comparativement à la mémoire de travail (Cepeda, Kramer, & Gonzalez de Sather, 2001; Davidson, Amso, Anderson, & Diamond, 2006; Garon et al., 2008). La mémoire de travail se développerait durant les premiers mois de vie (Diamond, 1985), tandis que la flexibilité cognitive émergerait seulement vers l’âge de 3 ans (Marcovitch & Zelazo, 2009). Ce délai nécessaire avant que la flexibilité ne se développe pourrait être le signe que cette habileté se développe avec la pratique et donc par le biais de mécanismes environnementaux. Certaines études expérimentales démontrent d’ailleurs que l’on peut entraîner la flexibilité cognitive en pratiquant des activités qui la sollicitent (Howard, Powell, Vasseleu, Johnstone, & Melhuish, 2017). Il est toutefois aussi possible que ce délai (le développement plus tardif de la flexibilité) s’explique par des facteurs génétiques qui interviennent plus tard dans le développement. C’est pourquoi davantage d’études incluant des mesures répétées à plus long terme de la flexibilité cognitive sont nécessaires pour clarifier les mécanismes qui sous-tendent le développement de cette habileté.
La capacité à prendre en compte différentes perspectives simultanément, à s’adapter au changement et naviguer de façon flexible entre différentes tâches et demandes environnementales, pourrait être moins sensible aux influences génétiques et se développer