Published by Elsevier
FEC 00090
Biological oxidation of hydrogen in soils
flushed with a mixture of
H2, CO2, 0 2
and
N 2
(Hydrogen-oxidising bacteria; landfill; hydrogen evolution; soil biofilters)L u c i a D u g n a n i b, Isabelle W y r s c h
a,b,
M a u r o G a n d o l l a b a n d Michel A r a g n o a,. a Laboratoire de Microbiologie, Institut de Botanique, Universitk de Neuchatel, Chantemerle 22, 2000 Neuch~tel 7, Switzerland,and b Consorzio per l'Eliminazione dei Rifiuti del Luganese, Bioggio, Canton Tessin, Switzerland
Received 1 August 1986 Revision received 23 September 1986
Accepted 26 September 1986
1. SUMMARY
A stainless steel cylinder filled with soil was flushed upstream with a H 2 / C O 2 / a i r mixture. The consequence was a strong enrichment of the aerobic, autotrophic hydrogen-oxidising micro- flora, which reached densities enabling them to oxidize 84.5 ml H 2 •dm -2- h -x in the first 25-cm layer. H 2 concentration profiles, hydrogen uptake activity and cell numbers correlated well with each other. Most of the organisms isolated were di- nitrogen fixers. Thus, soils containing hydrogen- oxidising bacteria may act as a biological shield between H2-rich environments and air, and may be utilized as biofilters, e.g., in the waste- processing industry.
2. I N T R O D U C T I O N
The autotrophic, aerobic hydrogen-oxidising bacteria are able to utilise the reducing power and oxidation energy of molecular hydrogen to per- form carbon dioxide fixation [1,2]. Most of them
* To whom correspondence should be addressed.
also utilise organic compounds as carbon and energy sources. Hydrogen is taken up only above a threshold concentration [3]. This threshold lies in the range of 1-10 nM for most of the strains studied. It is higher than both the H 2 concentra- tion in equilibrium with the atmosphere and the concentration generally found in steady-state sys- tems such as soil and fresh water [4,5]. Thus, ecological niches where these bacteria may express their H2-oxidising capabilities are restricted to oxic environments with hydrogen concentrations above this threshold.
A strong hydrogen production may arise from dumping grounds where organic matter is de- posited under conditions of restricted oxygen dif- fusion [6-8]. Under such conditions, microbiologi- cally catalysed events occur in a certain sequence (Fig. 1) beginning with oxygen exhaustion and climaxing with the full operation of the methano- genic syntrophy. Between these two events, a transitory fermentative phase takes place, accom- panied by a first peak of gas evolution, the main components being H 2 (up to 30%) and CO 2. Then the gas production decreases dramatically, and H 2 is progressively replaced by methane.
This H2-evolving phase is accompanied by the production of volatile organic compounds, and is 0168-6496/86/$03.50 © 1986 Federation of European Microbiological Societies
1(]0 G a s [ % Vol 1 gO , / " ... " ... " " • -. ... .,, B 0 / " " " " ' ' " 70 ~ " BO ~ / .... "'"" 5(] ~ / ".v t 40 A : / '"--.2-"""-"-, ... ...: ~ ,-" % Vol C02 ... 3() ..: i ~ ~ , - /---~, , . _ , ~ y " ;" % Vo] CH 4 ... 2 0 ~ ~I "\ Z Vol H2 . . . . lD ~ / ' ~ \ , , f - _ , f . . . . , , , , ~ - ~ , Z Vo] N2--- O \~ ; "" _ _" ... ---r---"" . T ~ - - - 4 ~ . . . % Vol 02 , , "Gas "Flow [ 1 / d a y ] pH [] 60 ... . ... ~ ~ ... " 40 pH ... 0 " < " ' , " ~ - - ' , ~ , I ' - • ' ' ' ' ' ' ~ - ~ ' ' l (] 20 30 4~ 60 80 l 0(] 120 14 (] 160 pH 8 6 4 TIME [ days ] ]80
Fig. 1. Evolution of gases from a mixture of organic wastes deposited under anoxic conditions. A 1600-1 hard polyethylene vessel without external air access was filled with a mixture of 443 kg urban waste (65$ dry matter content) ground and sorted at diameter < 8 cm and 200 1 digested sludge (5$ dry matter content). The percolating liquid was recirculated from the bottom to the surface. The temperature was maintained at 33-35°C. Total gas production was recorded continuously and gas composition was analysed at regular intervals.
m a i n l y r e s p o n s i b l e for t h e o d o u r s e v o l v i n g f r o m l a n d f i l l d i s p o s a l s a n d g a r b a g e t a n k s .
C o v e r i n g a c o m p a c t e d l a n d f i l l w i t h 3 0 - 5 0 c m o f soil a l m o s t c o m p l e t e l y e l i m i n a t e s the e m i s s i o n o f o d o u r s [9]. T h e soil acts h e r e as a s u p p o r t for b a c t e r i a l g r o w t h at the i n t e r f a c e b e t w e e n the a n a e r o b i c g a r b a g e a n d air. T h e r e , a n a e r o b i c b a c t e r i a l f l o r a c o u l d o x i d i s e t h e gases a n d o t h e r v o l a t i l e c o m p o u n d s e v o l v i n g f r o m t h e d e c o m p o s - i n g w a s t e mass. A s i m i l a r p r o c e s s c o u l d o c c u r in soil o r c o m p o s t b i o f i l t e r s (Fig. 2) u s e d to d e p u r a t e
soil biof l i t e r garbage tank
Fig. 2. Scheme of the possible use of a soil biofilter to depurate the gases evolving from a waste container. 1, waste gas pump; 2, air pump.
t h e m i x t u r e o f air a n d gases e v o l v i n g in a n u r b a n w a s t e c o n t a i n e r , e.g., in a n i n c i n e r a t i o n p l a n t .
T h e a i m o f the r e s e a r c h r e p o r t e d h e r e was to test the effect o f flushing a soil s i m i l a r to t h a t c o v e r i n g a d u m p i n g g r o u n d w i t h a n a i r : H 2 : C O 2 m i x t u r e , a n d to f o l l o w t h e k i n e t i c s a n d d i s t r i b u - t i o n o f h y d r o g e n - o x i d i s i n g a c t i v i t y a n d the d i s t r i - b u t i o n a n d c h a r a c t e r i s t i c s o f t h e h y d r o g e n - o x i d i s - i n g b a c t e r i a in such a n e n v i r o n m e n t . 3. M A T E R I A L S A N D M E T H O D S 3 . 1 . S o i l c o l u m n
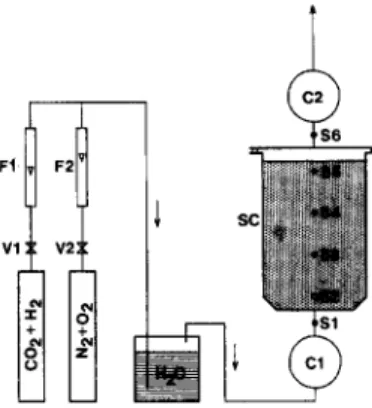
T h e a p p a r a t u s u s e d was a 30-1 stainless steel c o l u m n , 28 c m in d i a m e t e r (Fig. 3). It was filled w i t h a n a l l u v i a l siliceous s a n d w i t h a m o i s t u r e c o n t e n t o f a b o u t 2 0 $ . T h e c o l u m n was f l u s h e d u p w a r d s w i t h a m i x t u r e o f 1 / 3 a r t i f i c i a l air (20% 0 2 + 80% N 2 ) a n d 2 / 3 o f a 20% H 2 + 80% C O 2 m i x t u r e . T h e s e p r o p o r t i o n s a l l o w e d c o m p l e t e o x i d a t i o n o f h y d r o g e n to occur. T h e H 2 a n d C O 2 c o n t e n t s c o r r e s p o n d e d to the m e a n c o m p o s i t i o n
sample (1 ml inoculum, 5 tubes/dilution). The tubes were incubated with slight agitation (80 r e v . / m i n ) under a H 2 / O 2 / C O 2 mixture (85 : 5 : 10) at 27°C for 14 days.
The procedures for isolation and identification of hydrogen-oxidising bacteria have been de- scribed elsewhere [1].
Fig. 3. Scheme of the experimental soil column. V1, V2, precision valves; F1, F2, flow meters; C1, C2, gas counters; SC, soil column; $1-$6, gas sampling points.
of the gases produced during the hydrogen-evol- ving phase of anaerobic waste decomposition.
The system was fitted with 6 gas-sampling points at different levels, 2 flow meters with preci- sion valves to control the gas admission at the input of the fermenter, 2 gas meters to measure the input and output gas flows, and 3 temperature probes inside the soil column. It was run at am- bient temperature.
3.2. Gas analysis
The gas was analysed using a two-column gas chromatograph (GOW-MAC) fitted with a molec- ular sieve (5A, 60-80 mesh) and Carbosieve B (120-140 mesh) columns (Supelco, Bellefonte, PA, U.S.A.) and with a thermal conductivity detector. This set-up allowed us to separate and detect all the components in the gas mixture.
3.3. Hydrogen uptake measurements
Hydrogen uptake activity was measured in a Warburg respirometer, with oxygen as terminal electron acceptor, as described by Aragno and Schlegel [1].
3.4. Culture conditions
A basal mineral medium for hydrogen-oxidis- ing bacteria [1] was used throughout this study. Most probable number (MPN) determinations were performed by enrichment culture in 18-ram- diameter tubes containing 4 ml of mineral medium, inoculated by a series of decimal dilutions of the
4. RESULTS
The gas mixture was first provided at a rate of 285 ml- d m - 2 . h-1. The kinetics of gas composi- tion measured at the output are shown in Fig. 4. After 450 h, H 2 completely disappeared while flowing through the column, provided that 02 was presentl in excess. The input and output gas com- positions are shown in Table 1. The subsequent reappearance of hydrogen was due to an acciden- tal decrease in 02 concentration in the input gas mixture. This confirms that oxygen was the main electron acceptor for hydrogen uptake as expected for an aerobic HE-lithotrophic respiration.
Then, the total gas flow was increased to 640 ml. dm -2. h - t . This flow was of the same order of magnitude as the maximal flow expected from the anaerobic decomposition of a 1-m layer of garbage. Increasing the gas flow up to this value did not result in the reappearance of hydrogen at the output, indicating that the bacterial flora was undersaturated with respect to the amount of H E provided. Gas sampling at different levels in the
15- 10- Vol % ~ \ H 2 \ 0 0 \ \ \ / , , , ', 200 400 600 800 Time (hr)
Fig. 4. Amounts of H 2 and 0 2 in the gas at the output of the column with time.
Table 1
Composition of the gas mixture at the input and output of the soil column after 767 h of flushing
Gas Input Output
(%) (%)
H 2 13.3 0
02 6.7 1.7
CO 2 53.3 65
N 2 26.7 33.3
cylinder p r o v i d e d profiles like the typical one s h o w n in Fig. 5.
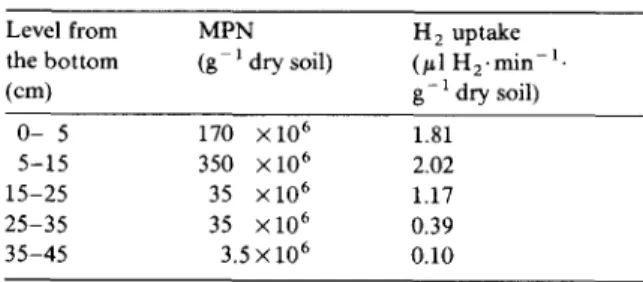
A t the end of the experiment, after 767 h, H : h a d completely d i s a p p e a r e d in the first 25 c m of soil. Samples were taken at different depths, respi- rometric h y d r o g e n u p t a k e activity was measured, a n d estimates of the cell n u m b e r s of aerobic h y d - rogen-oxidising bacteria were m a d e (Table 2). T h e distributions of h y d r o g e n - u p t a k e activity a n d cell n u m b e r s correlate well with each other and with the gas c o n c e n t r a t i o n profiles. Based on an as- s u m e d d r y weight o f 0.2 /xg per 106 cells a n d a protein c o n t e n t of 60% of the cell d r y matter, the specific h y d r o g e n u p t a k e activity in the 5 - 1 5 c m level would be 2860 /~1 H : . h - 1 . m g -1 protein, which is in the same order as specific activities m e a s u r e d in pure a u t o t r o p h i c cultures of m o s t h y d r o g e n - o x i d i s i n g bacteria. 5o [cm] 40 cm s o i l column I0 I [Vol %]
Fig. 5. A typical profile of H 2 and 0 2 contents in the soil
column. Time after the beginning of the experiment, 767 h; total gas flow, 640 ml- din- 2. h- t.
Table 2
Numbers of hydrogen-oxidising bacteria (determined by the most probable number method) and specific H2-uptake activ- ity of the soil column after 767 h of flushing
Total gas flow = 640 ml.dm 2.h 1
Level from MPN H 2 uptake
the bottom (g 1 dry soil) (/d H2.min -1.
(cm) g- 1 dry soil) O- 5 170 X 106 1.81 5-15 350 X 106 2.02 15-25 35 x l O 6 1.17 25-35 35 × 1 0 6 0.39 35-45 3.5 × 106 0.10
F r o m the last positive dilutions in enrichment cultures, we isolated different types of h y d r o g e n - oxidising bacteria. T h e m o s t frequent ones were
strains related to
Pseudomonas pseudoflava
a n d toXanthobacter autotrophicus,
as well as a third, n o tyet identified type. A t a x o n o m i c study of these different types is at present being u n d e r t a k e n in o u r laboratory. T h e bacteria share two properties of peculiar interest: (a) they are versatile, faculta- tire heterotrophs, able n o t o n l y to grow auto- trophically with molecular h y d r o g e n , b u t also to utilise a large variety of organic volatile c o m - p o u n d s . Thus, such organisms, although t h e y m a i n l y g r o w lithotrophically, would be able to r e m o v e o d o r o u s volatile organic c o m p o u n d s as well. (b) m o s t of our isolates are dinitrogen fixers. This allows t h e m to o v e r c o m e a limitation b y shortage of c o m b i n e d nitrogen in the soil. This explains w h y cell n u m b e r s as high as 350 × 106 g 1 were reached, i.e., one order of m a g n i t u d e higher than a n o r m a l ' t o t a l ' soil flora. The distri- b u t i o n of dinitrogen fixing activity in similar ex- perimental systems is at present u n d e r study in o u r laboratories.
5. D I S C U S S I O N
These results show that a u t o t r o p h i c h y d r o g e n - oxidising bacteria m a y play a m a j o r role in the aerobic oxidation of H 2 evolving f r o m a landfill or o t h e r m a c r o v o l u m e s of anaerobically decaying organic matter. T h e question arises whether such a p h e n o m e n o n , also a similar H 2 evolution f r o m
decaying organic m a t t e r a n d its oxidation at the o x i c - a n o x i c interface, occurs in microenviron- ments. C o n r a d a n d coworkers have p o i n t e d out that the kinetic characteristics of H 2 oxidation in n o r m a l soil [4] a n d in water bodies [3,5] do n o t meet those of 'classical' bacteria in pure cultures:
the a p p a r e n t K m was m u c h lower, a n d n o
threshold was noticed within the detection limits o f their analytical system (0.1 nM), thus allowing H 2 u p t a k e to occur even at atmospheric con- centrations. This activity was supposed to be due to soil enzymes rather than to soil microorganisms [10]. Thus, H2-oxidising bacteria would have a m o r e indirect effect on the atmospheric h y d r o g e n budget, preventing H 2 f r o m reaching the a t m o - sphere b y f o r m i n g a 'shield' between hydrogen-rich e n v i r o n m e n t s a n d air. W h e t h e r significant H 2 con- centrations occur at the o x i c - a n o x i c interface of freshwater sediments should be investigated fur- ther. We f o u n d relatively large n u m b e r s of h y d r o - gen-oxidising bacteria in the thin, oxidised sup- erficial layer of eutrophic freshwater sediments [11] (Aragno, unpublished results), b u t such an a c c u m u l a t i o n could also be the consequence of bacterial sedimentation f r o m the water c o l u m n (B. Schink, personal c o m m u n i c a t i o n ) .
This confirms the general a s s u m p t i o n that mic- r o o r g a n i s m s are globally m o r e i m p o r t a n t in re- m o v i n g volatile c o m p o u n d s p r o d u c e d in the bio- sphere before they reach the atmosphere, than in r e m o v i n g trace volatile c o m p o u n d s f r o m the a t m o s p h e r e at p p b or p p t c o n c e n t r a t i o n ranges. T h e use of hydrogen-oxidising bacteria in soil biofilters appears promising for the treatment of gases evolving f r o m decaying wastes.
A C K N O W L E D G E M E N T S
This w o r k was s u p p o r t e d b y grant 3.517-83 f r o m the Swiss N a t i o n a l Science F o u n d a t i o n . T h e
c o - o p e r a t i o n of the C E R , Bioggio, a n d valuable technical help b y G. Muschi were greatly appreci- ated.
R E F E R E N C E S
[1] Aragno, M. and Schlegel, H.G. (1981) The hydrogen- oxidizing bacteria. In The Prokaryotes (Starr, M.P., Stolp, H., Triiper, H.G., Balows, A. and Schlegel, H.G., Eds.) Vol. 1, pp. 865-893. Springer, Berlin.
[2] Bowien, B. and Schlegel, H.G. (1981) Physiology and biochemistry of aerobic, hydrogen-oxidizing bacteria. Annu. Rev. Microbiol. 35, 405-452.
[3] Conrad, R., Aragno, M. and Seiler, W. (1983) The inabil- ity of hydrogen bacteria to utilize atmospheric hydrogen is due to threshold and affinity for hydrogen. FEMS Mi- crobiol. Lett. 18, 207-210.
[4] Conrad, R. and Seller, W. (1979) The role of hydrogen bacteria during the decomposition of hydrogen by soil. FEMS Microbiol. Lett. 6, 143-145.
[5] Conrad, R., Aragno, M. and Seiler, W. (1983) Production and consumption of hydrogen in a eutrophic lake. Appl. Environ. Microbiol. 45, 502-510.
[6] Gandolla, M., Grabner, E. and Leoni, R. (1982) Ergeb- nisse yon Lysimetern auf der Deponie Croglio, Schweiz. VerSffentl. Inst. Stadtbauwesen Techn. Univ. Braun- schweig 33,163-182.
[7] Gandolla, M. and Grabner, E. (1983) Proposte per future ottimizzazioni di discariche controllate. Wasser, Energie, Luft 75, 241-244.
[8] Glauser, M., Gandolla, M. and Aragno, M. (1986) Anaerobic digestion of urban wastes: sewage sludge and organic fraction of garbage. In Bioenvironmental Systems (Wise, D., Ed.) Vol. 3. CRC Press, Boca Raton, FL, in press.
[9] Gandolla, M., Dugnani, L., Leoni, R. (1986) Valutazione dei processi chimici e biologici in uno scarico controllato per un corretto approccio alia progettazione e gestione. Ingegneria Ambientale, in press.
[10] Conrad, R. (1987) The biogeochemistry and ecophysio- logy of the atmospheric CO and H2 cycles. Adv. Microb. Ecol., in press.
[11] Schweizer, C. and Aragno, M. (1975) Etude des hydro- genobact~ries dans un petit lac (Le Loclat ou Lac de Saint-Blaise). Bull. Soc. Neuch$tel. Sci. Nat. 98, 79-87.