HAL Id: hal-02796488
https://hal.inrae.fr/hal-02796488
Submitted on 5 Jun 2020HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Effets de la nature de la ration et du niveau alimentaire
sur la valeur nutritionnelle des rations et la composition
de la carcasse chez des jeunes bovins
Yunlong Huo
To cite this version:
Yunlong Huo. Effets de la nature de la ration et du niveau alimentaire sur la valeur nutritionnelle des rations et la composition de la carcasse chez des jeunes bovins. Sciences du Vivant [q-bio]. 2014. �hal-02796488�
INRA-UMRH-Equipe AMUVI
Yunlong HUO
Janvier – Juin 2014
Faculté de Médecine 28 Place Henri Dunant 63001 Clermont Ferrand Cedex 1
89 Avenue de l’Europe 63370 Lempdes
INRA-UMRH Theix 63122 Saint Genès Champanelle 28 Place Henri Dunant
63000 Clermont Ferrand Faculté de Médecine
28 Place Henri Dunant BP38 63001 Clermont Ferrand Cedex 1
Effets de la nature de la ration et du niveau alimentaire sur la valeur
nutritionnelle des rations et la composition de la carcasse chez des
jeunes bovins
Master Sciences, Technologies, Santé – mention Nutrition, Santé, Aliments
Spécialité Nutrition Animale et Elevage – 2ème année
Rapport de Stage
Responsable de stage
Isabelle ORTIGUES-MARTY
Mjhgg21214545MKFJKJLM
Responsable Master
Fabienne BLANC
Remerciements
Je tiens à remercier sincèrement Isabelle ORTIGUES-MARTY, ma maître de stage, pour son accueil, son aide et ses conseils, ses corrections tout au long du stage, mille mercis à Isabelle puisque j’ai beaucoup appris pendant ce stage et j’en très apprécie.
Un très grand merci à Jacques AGABRIEL pour m’avoir initié au modèle MECSIC, ses conseils pour la rédaction de rapport et ses corrections.
Je tiens à remercier Vincent LARGEAU pour m’avoir accompagné pendant les analyses chimiques, qui ne sont pas toujours évidentes, et aussi pour ses conseils et corrections pour le rapport.
Je voudrais aussi remercier Gonzalo CANTALAPIEDRA-HIJAR pour son aide aux traitements de données et pour sa patience d’avoir répondu mes milliers de questions.
Je tiens à remercier monsieur Bernard SEPCHAT pour son aide sur les résultats.
Je tiens à remercier Jean VERNET, dont sa gentillesse m’a touché profondément, pour son aide des calculs et son encouragement.
Ce stage a été effectué au Centre de Recherche de l’INRA de Clermont-Ferrand/Theix, j’ai été rattaché à l’équipe Animal Muscle Viande (AMUVI). Je tiens donc à remercier l’ensemble du personnel de cette équipe. Grâce à eux j’ai passé 6 mois très agréables, et partagé de nombreux moments conviviaux avec eux.
Table des abréviations
ADF : Acide Detergent Fibre
ADFI : Acide Detergent Fibre Ingéré
CEM : Concentration en Energie métabolisable
DA5Q : Dépôt Adipeux dans le Cinquième Quartier
DACA : Dépôt Adipeux dans la Carcasse
DAT : Dépôt Adipeux Totaux
dE : Digestibilité de l’Energie
dMO : Digestibilité de la Matière Organique
dr : Digestibilité Intestinale des Protéines Alimentaires non Dégradées
DT : Dégradabilité Théorique des Matières Azotées dans le Rumen
EB : Energie Brute
EBI : Energie Brute Ingérée
ED : Energie Digestible
EDI : Energie Digestible Ingérée
EM : Energie Métabolisable
EMI : Energie Métabolisable Ingérée
EN : Energie Nette
ERH : Traitement d’Enrubannage d’herbe avec un haut niveau énergétique
ERB : Traitement d’Enrubannage d’herbe avec un bas niveau énergétique
ESM : Erreur Standard de la Moyenne
GEB : Groupe Economie du Bétail
GMQ : Gain Moyen Quotidien
GPV : Gain de Poids Vif
IMF : Intramuscular Fat
INRA : Institut National de la Recherche Agronomique
ISO :
International Organization for Standardization
j : jour
JB : Jeune Bovin
kg : kilogramme
LipC : Lipides dans la Carcasse
LipNC : Lipides dans le cinquième quartier
MAT : Matières Azotées Totales
MD : Masse Délipidée
MECSIC : modèle MECaniste pour la Simulation de la Croissance
MJ : Mégajoule
MM : Matière Minérale
MO : Matière Organique
MOI : Matière Organique Ingérée
MOF : Matière Organique Fermentescible
MOFI : Matière Organique Fermentescible ingérée
MPEEH : Traitement d’Ensilage de maïs avec un haut niveau énergétique
MPEEB : Traitement d’Ensilage de maïs avec un bas niveau énergétique
MS : Matière Sèche
MSI : Matière Sèche Ingérée
N : Azote
NF: Norme Française
NI :Azote ingéré
NDF : Neutral Detergent Fibre
NDF : Neutral Detergent Fibre Ingéré
PDI : Protéines Digestibles dans l’Intestin
PDIA : PDI d’origine alimentaire
PDIE : PDI permises par l’Energie
PDIM : PDI d’origine microbienne
PDIME : PDIM permises par l’Energie
PDIMN : PDIM permises par l’azote
PDIN : PDI permises par l’azote
ProtC : Protéines dans la Carcasse
ProtNC : Protéines dans le cinquième quartier
PV : Poids Vif
PVV : Poids Vif Vide
QI : Quantités Ingérées
UFL : Unité Fourragère « Lait »
UFV : Unité Fourragère « Viande »
SOMMAIRE
Introduction ... 1
Synthèse bibliographique ... 2
1.
EVOLUTION DE LA COMPOSITION CORPORELLE ET DE LA QUALITÉ DES CARCASSES CHEZ LES JEUNES BOVINS EN CROISSANCE... 2
2.
EVALUATION DE LA VALEUR NUTRITIONNELLE DES RATIONS... 3
3.
INFLUENCE DU NIVEAU ALIMENTAIRE SUR LES DÉPÔTS DE LA CARCASSE... 4
4.
INFLUENCE DE LA NATURE DES RATIONS SUR LES DÉPÔTS DE LA CARCASSE... 5
5.
MODÉLISATION DE LA CROISSANCE CHEZ LES BOVINS ET MODÈLE DE L’INRA
MECSIC ... 7
Matériels et Méthodes ... 8
E
XPERIMENTATIOND'
« E
NGRAISSEMENT»
... 8
E
XPERIMENTATIONDE
« D
IGESTIBILITÉ» ... 8
1. Animaux et logements ... 8
2. Aliments et Rations ... 9
3. Prélèvements et mesures ... 10
4. Calculs et analyses statistiques des résultats ... 10
5. Ajustement du modèle MECSIC ... 11
Résultats ... 13
1.
EXPERIMENTATION DE« D
IGESTIBILITÉ» ... 13
2.
EXPERIMENTATION D’« E
NGRAISSEMENT»
... 14
3.OPTIMISATIONDESPARAMÈTRESDUMODÈLEMECSIC………..
15
Discussion ... 16
1.
EXPERIMENTATION DE« D
IGESTIBILITÉ» ... 16
2.EXPERIMENTATIOND’« ENGRAISSEMENT »……….17
3. DYNAMIQUE DES DEPOTS ET AJUSTEMENT DU MODELE MECSIC……….19
Conclusion et perspective ... 20
Tableau 1 : Evolution de la production de Jeunes Bovins en France. Source : Estimations GEB-Institut de l’Elevage
Figure 1 : Origine de la viande produite et consommée en France en 2010 (Estimation GEB -Institut de l’Elevage - % du tonnage de Gros Bovins).
Introduction
D’après les données de l’Institut de l’Elevage (2011), en 2010, ce sont 903 000 jeunes bovins (mâles non castrés âgés de 12 à 24 mois) qui ont été abattus en France, soit un effectif en constante augmentation par rapport à 2000 (Tableau 1) (Economie de l’Elevage, 2011). La production de jeunes bovins, représentant environ un tiers de la production de viande bovine en France (Figure 1), se caractérise par des animaux ayant une croissance continue la plus proche possible de leur vitesse maximale de croissance, et une alimentation fortement énergétique à l’auge (INRA, 2007). Elle représente le système le plus intensif dans la production de la viande bovine.
Le système INRA d’alimentation des ruminants, largement utilisé dans la pratique, a pour objectif d’estimer la valeur alimentaire des aliments et les besoins des animaux afin d’ajuster les rations au potentiel de production des animaux. Dans bovins, leur engraissement en France repose sur l’utilisation de régimes très énergétiques à forte proportion d’ensilage de maïs ou de céréales, dont les prix fluctuent de façon importante. Avec la contrainte économique forte, les exigences des éleveurs en termes de la rentabilité économique n’ont jamais été aussi grandes. Les éleveurs de jeunes bovins s’intéressent à minimiser le coût des intrants tout en produisant une bonne qualité de carcasse, qui dépend à la fois de critères quantitatifs (poids), mais aussi de critères qualitatifs liés à la composition (protéines et lipides) de la carcasse et la répartition des dépôts. Il est donc intéressant de connaître précisément l’impact des apports nutritionnels des rations sur le développement musculaire et adipeux afin d’optimiser l’engraissement et l’abattage de la façon la plus rentable possible.
Mon stage s’inscrit dans le cadre du Programme Transversal UMRH Recommandations Alimentaires des Bovins Viandes et plus particulièrement d’une étude « CRoissance, Alimentation et DEpôts des taurillons CHArolais » (CRADECHA) dont l’objectif est de quantifier l’impact de la nature de la ration ingérée et du niveau alimentaire sur l’orientation des dépôts protéiques et lipidiques au cours de la phase d’engraissement chez des taurillons Charolais. Cette étude s’appuie sur deux expérimentations : « Engraissement » et « Digestibilité ». La première permet d’étudier la croissance et les dépôts de lipides et de protéines ainsi que leur répartition, et la deuxième de déterminer la valeur nutritionnelle des rations.
Figure 2 : Evolution de la composition corporelle d’un jeune bovin laitier durant la période de croissance et d’engraissement (200-550 kg de poids vif) (Micol et al., 1993).
Mon travail de stage, contribuant à ces objectifs, portera sur différents points :
Détermination de la valeur nutritionnelle des rations.
Interprétation des résultats de dépôts adipeux et protéiques dans la carcasse selon les valeurs nutritionnelles mesurées.
Paramétrage du modèle MECSIC (MECaniste pour la SImulation de la Croissance) selon la valeur nutritionnelle des rations et les résultats de dépôts adipeux et protéiques dans la carcasse
Synthèse bibliographique
1. Evolution de la composition corporelle et de la qualité des
carcasses chez les jeunes bovins en croissance
On peut définir la croissance d’un animal comme l’accroissement de sa taille et de son poids ainsi que ses modifications de forme et de composition corporelle (INRA, 1986). Selon Micol et al. (1993), la croissance pondérale résulte du développement en masse de chacun des éléments constitutifs de son corps. La composition anatomique correspond à la part des différents tissus de la masse corporelle (muscle, dépôts adipeux, os) et des éléments du cinquième quartier (organes, peau, tractus digestif) (Figure 2), tandis que la composition chimique exprime l’importance relative des différents éléments chimiques (eau, protéines, lipides, minéraux…) contenus dans l’ensemble des tissus (Figure 3). La composition corporelle se modifie donc selon le développement différentiel de chacun des éléments.
D’une manière générale, chez les jeunes bovins, pour lesquels la croissance n’est pas terminée, la quantité de muscles augmente avec l’âge et le poids (Robelin, 1986). On peut donc séparer la période d’engraissement en deux phases distinctes : une phase de croissance et une de finition. Comme montré dans la Figure 2, au cours de la croissance puis de l’engraissement, la part du tissu adipeux dans le gain de poids augmente, et peut donc être supérieure à celle du tissu musculaire dans la phase de finition (Micol et al., 1993). Selon l’étude de Robelin (1986) visible dans la Figure 4, le poids des tissus adipeux varie de 6 à 30%, le poids des muscles de 30 à 50% en pourcentage du gain de poids vif, alors que les variations de poids de squelette restent quant à elles beaucoup moins importantes (8 à 13%).
Figure 3 : Evolution de la composition chimique du corps entier chez les bovins Pie-Noirs (Robelin et Daenicke, 1980 ; cité par INRA, 1986)
Figure 4 : Evolution de la composition du gain de poids vif et des quantités de muscles, de dépôts adipeux et de squelette déposées par jour (Robelin, 1986)
Pour décrire la croissance relative des différents tissus ou organes, des équations d’allométrie sont utilisées par Robelin (1974) :
ln Y = b ln X + a
Dans cette relation, Y est le poids de la région, de l’organe ou du tissu considéré, X est le poids de l’ensemble de référence, « a »est une constante, « b » est le coefficient d’allométrie de Y par rapport à X. Si « b » < 1, ce serait l’allométrie minorante, Si « b » > 1, l’allométrie est majorante, si « b » = 1, on parle d’isométrie.
La croissance et le développement des bovins sont déterminés par le type génétique et l’âge physiologique, mais ils sont également fortement dépendants des apports nutritionnels quantitatifs et qualitatifs. On peut ainsi faire varier la composition du croît et celle de la carcasse en jouant sur les apports alimentaires.
2. Evaluation de la valeur nutritionnelle des rations
Rationner l’alimentation des ruminants nécessite une bonne connaissance de la composition chimique et de la valeur alimentaire des fourrages et des autres matières premières. La composition chimique est obtenue à partir d’analyses chimiques (ex : MS, EB, azote, NDF, ADF etc…), elle est une caractéristique des aliments et est indépendante de l’animal. Par contre la valeur nutritionnelle dépend aussi de l’animal et nécessite des mesures de digestibilité et d’utilisation ou de pertes métaboliques. Elle peut être prédite à partir d’équations issues de résultats expérimentaux et rassemblées dans les systèmes d’alimentation. Nous rappelons ici les principes des calculs de la teneur en énergie nette en termes d’UFL et UFV et la valeur azotée des aliments en termes de PDIN et PDIE (Annexe 1) tels qu’ils sont décrits dans le système INRA (2007).
Pour calculer la teneur en énergie nette d’un aliment, il faut la teneur en énergie brute (EB), la digestibilité de l’énergie (dE) et le rapport EM/ED. La teneur en énergie brute d’un aliment se mesure par bombe adiabatique, elle peut aussi être prédite à partir de sa composition chimique. La digestibilité de l’énergie dépend fortement du niveau alimentaire, auquel sont faites les mesures pour construire les tables INRA. En plus, elle est très étroitement corrélée à celle de la matière organique (dMO). Elle peut donc être estimée directement selon dMO. Mais il faut tenir en compte que la quantité
Figure 5 : Poids des tissus déposés au cours de l’engraissement d’un jeune bovin laitier selon la vitesse de croissance moyenne retenue (0,8 et 1,0 kg/j) Micol et al. (1993).
Figure 6 : Influence du gain de poids journalier sur la composition du croît d’un jeune bovin laitier de 500 kg de poids vif Micol et al.., (1993).
d’énergie digestible (ED) d’une ration n’est pas égale à la somme des quantités d’ED de chaque aliment puisqu’il existe des interactions digestives entre des aliments. Le rapport EM/ED rend compte des pertes d’énergie métabolique dans les urines et d’énergie digestive sous forme de gaz (CH4). Ces pertes dépendent de la composition chimique de l’aliment et aussi du niveau alimentaire auquel a été réalisée la mesure de digestibilité.
La valeur azotée d’un aliment est exprimée en protéines digestibles dans l’intestin (PDI). Les tables donnent deux valeurs parallèles : PDIN et PDIE. La valeur PDIN représente la valeur PDI de l’aliment lorsque la synthèse de protéines microbiennes est limitée par l’apport en azote dégradable, et la valeur PDIE représente la valeur PDI lorsque la synthèse de protéines microbiennes est limitée par l’énergie disponible dans le rumen. Les valeurs PDIN et PDIE sont la somme des PDIA qui sont les protéines d’origine alimentaire non dégradées dans le rumen et des PDIM qui sont les protéines synthétisées dans le rumen et dont la production est limitée par l’énergie fermentescible (PDIME) ou par l’azote dégradable (PDIMN). Prévoir la valeur PDI d’un aliment nécessite d’évaluer deux paramètres spécifiques qui sont la dégradabilité théorique des matières azotées dans le rumen (DT), et la digestibilité intestinale des protéines alimentaires non dégradées (dr).
3. Influence du niveau alimentaire sur les dépôts de la carcasse
Les variations du niveau des apports alimentaires permettent de modifier la vitesse de croissance pondérale, la composition du croît des bovins et, en conséquence, la composition corporelle à l’abattage. La figure 4 illustre l’évolution de la composition du gain de poids vif et des quantités déposées de tissus adipeux et musculaires par jour selon la vitesse de croissance. Micol et al. (1993) ont démontré (Figure 5) que lorsque le gain de poids vif s’accroît (0,6 à 1,2 kg/j) du fait du niveau alimentaire, le dépôt de muscles représente 39% du gain de masse corporelle pour 0,6 kg/j et n’en représente plus que 27% pour 1,2 kg/j. Les dépôts adipeux correspondent respectivement à 29% et 50% de ces gains (Figure 6). Schoonmaker et al. (2003) ont aussi observé chez des bœufs de 218 j ayant eu un gain depoids moyen quotidien (GMQ) de 1,2 kg/j pendant 100 j résultait en une aire de musclelongissimus plus élevée qu’un GMQ de 0,8 kg/j, ce qui signifie une augmentation des dépôts
Figure 7 : Effet de l’énergie et de l’azote ingéré sur l’azote retenu par les ruminants (Balch, 1967)
Des études concernant l’effet de la restriction antérieure d’alimentation sur le développement des tissus ont été faites. Henrickson et al. (1965), ont remarqué qu’une diminution du niveau alimentaire entre 6 et 10-12 mois d’âge entraîne une croissance compensatrice ultérieure et conduit à des états d’engraissement moins importants en comparaison des animaux bien alimentés en continu. Des dépôts protéiques plus importants (Hoch et al., 2005) et une réduction de la croissance des tissus adipeux plus marquée que de celle des autres tissus (INRA, 1986) ont été obtenus après une restriction alimentaire.
Balch (1967) a développé le concept de « phases dépendantes à l’énergie ou aux protéines » pour la croissance des tissus maigres. Lorsque les quantités d’azote ingéré sont inférieures aux besoins, l’azote fixé est corrélé linéairement à l’azote ingéré et n’est pas affecté par l’EMI (énergie métabolisable ingérée (phase protéine-dépendante). A l’inverse, lorsque les quantités d’azote ingéré sont en excès, l’azote fixé est corrélé linéairement avec l’EMI et n’est pas affecté par l’azote ingéré (phase énergie-dépendante) (Figure 7). On peut ainsi déduire que l’azote et l’énergie sont deux facteurs indispensables qui font varier les dépôts protéiques et adipeux dans la carcasse, et que l’un ou l’autre peuvent être limitant selon qu’ils soient apportés en adéquation ou non par rapport aux besoins des animaux.
4. Influence de la nature des rations sur les dépôts de la carcasse
Il y a beaucoup d’études dans lesquelles les animaux sont alimentés ad libitum afin de comparer l’influence de la nature de la ration. Micol et al. (2007) ont observé que les animaux engraissés ad libitum avec une ration à base d’ensilage de maïs ont présenté plus de dépôts adipeux dans la carcasse que ceux engraissés ad libitum avec une ration à base de concentré ou une à base de foin pour la finition de jeunes bovins de race Blonds d’Aquitaine. Keady et
al. (2013) ont fait une comparaison entre des ensilages. Ils ont alimenté les taurillons avec 4kg
de concentré, mais les fourrages (ensilage de maïs vs ensilage d’herbe) sont offerts ad libitum. Il est observé que le lot nourri de l’ensilage de maïs a eu plus de poids de carcasse et déposé plus de tissus adipeux dans la partie non- carcasse que le lot ensilage d’herbe, mais la différence pour les dépôts de tissus adipeux dans la carcasse n’est pas significative. En plus, selon Schoonmaker et al. (2003) et Colman et al. (1995), l’épaisseur du gras et l’aire de muscle longissimus dans la carcasse, qui font partie respectivement les dépôts adipeux et protéiques, ont été augmentés pour les bœufs alimentés une ration ad libitum avec haut niveau de fourrage par rapport aux ceux nourris ad libitum avec une ration riche en concentré dans la phase de croissance, par contre le pourcentage de protéine longissimus s’est diminué.
Schoonmaker et al. (2003) et Schoonmaker et al. (2004) ayant utilisé des bœufs âgés de 119j (sevrés précocement) ont remarqué une amélioration du gras intramusculaire (IMF) après une période de croissance (100 j ou 153 j) lorsque les bœufs sont alimentés une ration de concentré ad libitum par rapport à une ration riche en fibre ad libitum. Des résultats similaires ont été remarqués par Wood et al. (2008) et Rhodades et al. (2007). Le dernier a observé que les rations riches en amidon distribuées à des bœufs de 8 mois, normalement sevrés, peuvent augmenter la disponibilité et l'absorption de glucose ainsi que la synthèse des acides gras à l’IMF, tandis que les animaux qui ont reçu des rations riches en fourrage ont diminué la disponibilité de glucose sans changements dans l'incorporation d'acétate dans les acides gras. Cela augmente la proportion d’IMF selon ce que Smith et Crouse (1984) avaient remarqué : les adipocytes intramusculaires utilisent préférentiellement le glucose comme substrat pour la synthèse des acides gras, tandis que la graisse sous-cutanée utilise l'acétate.
Des études sur des animaux alimentés avec des rations iso-énergétiques ont également été réalisées. Colman et al. (1995) ont comparé l’effet des céréales et de l’ensilage au même niveau d’EMI, et ont montré que les bœufs nourris avec des céréales ont eu des poids de carcasse et de carcasse plus élevés, déposé plus de tissus adipeux dans la carcasse et non-carcasse que ceux alimentés ad libitum avec une ration à base d’ensilage de sorgho durant la phase de croissance. Geay et al. (1997) ayant construit trois rations iso-énergie nette (EN) et iso-azote (PDI) avec différents fourrages (foin, ensilage d’herbe, ensilage de maïs), ont remarqué que les taurillons engraissés avec la ration à base de foin ont déposé moins de lipides dans la carcasse que ceux engraissés avec l’ensilage de maïs ou l’ensilage d’herbe. Des rations approximativement iso-énergétiques et iso-azotées ont aussi été établies par Costa et
al. (2013) à haut niveau d’ensilage de maïs (70% d’ensilage de maïs et 30% de concentré) et
bas niveau d’ensilage de maïs (30% d’ensilage de maïs et 70% de concentré). Ils ont noté que le lot alimenté à bas niveau d’ensilage de maïs a déposé plus de tissus adipeux dans la carcasse et dans les viscères en comparaison de celui de haut niveau d’ensilage de maïs.
Si la nature de la ration a un impact sur la composition des carcasses, c’est sans doute parce qu’une fois ingérée, elle influence la population microbienne dans le rumen, qui ensuite influence à son tour le type et les quantités de nutriments absorbables libérés après digestion (acides gras volatils, glucose, lactate et acides aminés) et par conséquent peut influencer le développement des tissus et la composition corporelle à l’abattage. Les rations riches en amidon favorisent un niveau de propionate plus élevé à partir de la fermentation ruminale
et/ou une plus grande quantité de glucose absorbé dans l'intestin grêle venant de l'amidon non dégradable par rapport aux rations riches en fourrages (Schoonmaker et al., 2010). Cela provoque une augmentation d’insulinémie (Graugnard et al., 2009), une amélioration d’utilisation du glucose par les tissus périphériques en faveur de la lipogenèse (Schoonmaker
et al., 2003).
5. Modélisation de la croissance chez les bovins et modèle de
l’INRA MECSIC
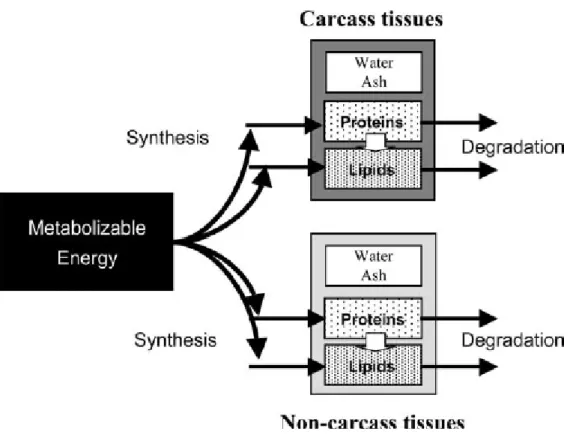
Un modèle nommé MECSIC (modèle MECaniste pour la SImulation de la Croissance), a été conçu et construit par Hoch et Agabriel (2004a), peut être utilisé dans le système INRA de recommandations alimentaires pour simuler la croissance des bovins et la composition corporelle de différents types d'animaux dans diverses conditions nutritionnelles. La présentation du modèle et les équations, le schéma du modèle sous Vensim, ainsi que ses paramètres sont respectivement en Annexe 2, 3 et 4. Ce modèle prend en compte les processus majeurs impliqués dans l'évolution des protéines et des lipides dans la carcasse et non-carcasse. Cette évolution est contrôlée par des procédés de synthèse et de dégradation de macromolécules qui dépendent de l’âge physiologique des animaux et de l’apport d’énergie métabolisable (Figure 8). Pour un apport d’EM donné, il simule l’accrétion (différence synthèse et dégradation) des tissus lipidiques et protéiques dans la carcasse et dans le cinquième quartier. Cependant, il présente des limites : il considère l’énergie métabolisable comme l’intrant, mais ne tient pas en compte sa composition, c’est-à-dire que le type de ration ingérée par des animaux n’est pas détaill à l’entrée du modèle. Cette prise en compte pourrait permettre de mieux expliquer et modéliser l’orientation des dépôts adipeux et protéiques dans la carcasse, la prédiction des réponses des animaux à différents types de régimes devrait ainsi être plus précise.
Matériels et Méthodes
Cette étude s’appuie sur deux expérimentations : « Engraissement » et
« Digestibilité », les deux essais ayant utilisé exactement les mêmes rations.
Expérimentation d’ « Engraissement »
Celle-ci reposait sur 36 broutards Charolais, sevrés à 8 mois et présentant un poids vif initial de 360 ± 33 kg. Ils ont été répartis en 4 lots homogènes selon leurs performances avant sevrage. Ils ont reçu deux types de rations à base d’ensilage de maïs (MPEE) et d’enrubannage d’herbe (ER), distribuées à 2 niveaux alimentaires (haut : 0,97 et 0,94 UFV/kg MS ; ou bas : 0,92 et 0,90 UFV/kg MS, ces valeurs viennent de Sepchat et al., (2013)). Les objectifs de croissance théorique des animaux sont de 1600g de gain moyen quotidien (GMQ) pour les lots hauts et 1400g pour les lots bas. Les animaux ont été abattus lorsque les 4 lots avaient le même poids vif moyen, soit 700 kg pour atteindre un objectif de poids de carcasse de 420 kg. L’âge des animaux à l’abattage fut donc variable, de 15 à 18 mois.
Durant toute la période de l’expérimentation, les animaux ont été pesés tous les quinze jours et les quantités quotidiennes de fourrages et de concentrés distribuées ainsi que les refus ont été mesurés individuellement. Les organes et viscères furent pesés à l’abattage et les rendements de carcasse calculés.
Expérimentation de « Digestibilité »
1. Animaux et logements
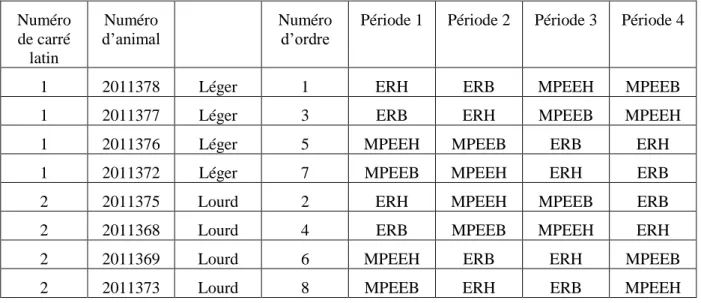
L’expérimentation a été réalisée sur 8 taurillons Charolais âgés de 10 mois (sevrés à l’âge de 8 mois) et de poids initial moyen de 405 ± 26,5kg. Un schéma factoriel 2x2 (2 rations à 2 niveaux d’énergie différents) et un Carré Latin répliqué (Tableau 2) permettant de supprimer les effets « animaux » et « périodes » ont été construits. Deux groupes de 4 animaux ont été constitués (1 groupe par Carré Latin) d’après leur poids initiaux afin d’enlever l’effet pondéral. La mise en groupe s’est faite en fonction de l’âge, du poids et du GMQ (naissance-sevrage). Les animaux ont reçu chacun successivement les 4 traitements alimentaires (administrés selon 4 périodes) :
Tableau 2 : Schéma de carré latin de « Digestibilité »
Numéro de carré latin Numéro d’animal Numéro d’ordrePériode 1 Période 2 Période 3 Période 4
1 2011378 Léger 1 ERH ERB MPEEH MPEEB
1 2011377 Léger 3 ERB ERH MPEEB MPEEH
1 2011376 Léger 5 MPEEH MPEEB ERB ERH
1 2011372 Léger 7 MPEEB MPEEH ERH ERB
2 2011375 Lourd 2 ERH MPEEH MPEEB ERB
2 2011368 Lourd 4 ERB MPEEB MPEEH ERH
2 2011369 Lourd 6 MPEEH ERB ERH MPEEB
- Ensilage de maïs, avec un haut niveau énergétique (MPEEH)
- Ensilage de maïs avec un bas niveau énergétique (MPEEB)
- Enrubannage d’herbe avec un haut niveau énergétique (ERH)
- Enrubannage d’herbe avec un bas niveau énergétique(ERH)
Chacune des 4 périodes a duré 3 semaines, dont 2 semaines d’adaptation (S1 et S2) et 6 jours de mesures (S3). Les animaux étaient installés en case individuelle avec des équipements de collecte de fèces et d’urine.
2. Aliments et Rations
2.1 Rationnement pré-expérimental
Dès leur arrivée dans les installations et durant 4 semaines, les 8 animaux ont été alimentés avec un régime comprenant du foin et du concentré pour mesurer leur capacité d’ingestion, afin de calculer les apports alimentaires offerts aux animaux et d’éviter qu’il y ait du refus durant la phase expérimentale.
2.2 Aliments et Régimes expérimentaux
Les deux régimes à base d’ensilage de maïs sont constitués de concentré blé-maïs, de tourteau de colza, d’urée et de lithotamne tandis que les deux régimes à base d’enrubannage d’herbe ont été complémentés avec de la drêche de blé, de la pulpe de citrus et du lithotamne (Tableau 3). Les fourrages et les concentrés font respectivement varier la nature de la ration (amidon vs fibre) et le niveau alimentaire (haut vs bas).
2.3 Rationnement
Les apports alimentaires ont été établis en fonction de la capacité d’ingestion et des besoins alimentaires des animaux (énergie, azote, minéraux) prédits par le système d’alimentation de l’INRA. Les rations avaient les mêmes teneurs estimées initialement en UF et PDI au sein de chacun des niveaux Haut et Bas.La Composition en ingrédient et les valeurs nutritionnelles estimées des rations sont marquées dans le Tableau 3. Les animaux ont reçu 2 repas par jour : à 9h et 16h.
Tableau 3 : Composition en ingrédient et valeurs nutritionnelles estimées des
rations de jeunes bovins charolais de « Digestibilité »
MPEEH MPEEB ERH ERB
Proportion des aliments (g/100g MS) Ensilage de maïs 52,3 65,4 Blé-maïs 26,8 17,0 Tourteau colza 19,1 15,7 Enrubannage d’herbe 43,6 49,3 Drêche blé 24,4 21,9 Pulpe citrus 30,8 27,5 Urée 0,5 0,5 Lithotamne 1,3 1,3 1,3 1,3 Valeurs nutritionnelles UFV (/kg MS) 0,93 0,87 0,93 0,87 PDIN (g/kg MS) 100,53 88,95 100,66 92,50 PDIE (g/kg MS) 94,47 86,84 94,47 86,84 PDI/UFV 101,13 100 101,13 100
Pour tenir compte de la croissance des animaux au cours de l’expérimentation, les quantités de MS offertes ont été ajustées à chaque période expérimentale. Pendant S1, les animaux ont reçu leur nouvelle ration ad libitum. Les rations ont été ajustées et fixées au bout de 12 jours en fonction de la croissance et de la capacité d’ingestion. Les changements de régime se sont déroulés pendant les deux derniers jours de la période (jour 20 et jour 21), 50% du régime précédant et 50% du régime suivant pour le jour 20 et 75% du régime suivant et 25% du régime précédant pour le jour 21. Ainsi les taurillons ont reçu le nouveau régime à 100% le premier jour de la période suivante.
3. Prélèvements et mesures
Prélèvement des échantillons : en semaine de digestibilité (S3), les différents ingrédients de la ration sont prélevés individuellement et quotidiennement puis poolés sur la semaine par aliment et par période. Les refus sont pesés chaque jour et prélevés s’ils excèdent 500g frais/jour, puis poolés par animal et par période. Les féces et les urines ont été pesées chaque jour et collectées en pool par animal/période, leurs prélèvements ont été réalisés en utilisant des équipements de récolte. Les matières sèches ont été mesurées juste après les prélèvements pour tous les échantillons ci-dessus (exceptées les urines).
Mesures et analyses chimiques : Pour les échantillons des aliments offerts, des refus et des féces, nous avons mesuré la teneur en matière sèche (MS, pour les ensilages: 60 °C pendant 48h ; pour les concentrés, les refus et les féces : 103°C pendant 24h), la teneur en matière organique (MO) (NF ISO 6496, 2011), le pourcentage de NDF (Neutral Detergent Fiber) et d’ADF (Acid Detergent Fiber) (NF V18-122, 1997), l’azote total (N-Kjeldahl total) (ISO 5983-1:2005) et l’énergie brute (EB) (IKA Calorimeter system C200). Nous n’avons mesuré que la teneur en N-Kjeldahl total pour les échantillons d’urine.
4. Calculs et analyses statistiques des résultats
Les quantités d’aliments ingérés ont été calculées à partir des quantités d’aliments distribuées et des quantités refusées. La digestibilité apparente (MS, MO, N, énergie, NDF et ADF) et le bilan d’azote ont été calculés de la façon suivante, si on prend l’exemple de N :
Digestibilité apparente de N (%) = (quantités ingérées de N – quantité fécale de N) * 100/ quantités ingérées de N
Les calculs de la valeur énergétique mesurée des rations sont indiqués ici :
ED = EB - EBféces en kJ/j
EM = ED – EBCH4 – EU en kJ/j
EB-CH4 = [Quantité (g/j)/16,0425 (g/mol)] * 891 (kJ/mol) en kJ/j
EU/EB = 2,9 + 0,017 MAT – 0,47 NI – 1,64 PCO en %
Avec MAT : matière azotée totale en g/kg MS dans la ration ; NI : niveau d’ingestion de MS (kg de MSI/100kg de poids vif, MSI%PV) ; PCO : Proportion de concentré dans la ration (0 < PCO < 1).
EU = EB * (EU/EB)/100 en kJ/j
ENEV = EM * kmf en kJ/j kmf est calculé en Annexe 1
UFV = ENEV (kJ/j) * 0,239 (kcal/kJ)/1820 (kcal)
La valeur azotée mesurée des rations est calculée par aliment de la même façon que celle en Annexe 1.
Une analyse variance a été réalisée sur l’ingestion et la digestibilité des différents éléments dans la ration et la partition d’azote à l’aide de logiciel R, avec 5 facteurs fixes : la nature de la ration, le niveau alimentaire, la période, le carré latin, ainsi que l’interaction entre la nature de la ration et le niveau alimentaire. Le facteur animal a été considéré comme aléatoire. Nous avons obtenu, grâce à cette analyse, une moyenne pour chaque variable et des différences significatives entre les lots (test à 5% d’erreur).
5. Ajustement du modèle MECSIC
La dynamique du modèle de croissance MECSIC est donnée par l’ingestion (MSI) et la concentration en énergie métabolisable des différentes rations recalculées par semaine (CEM en MJ/kg MS). Les 3 paramètres sensibles du modèle (cMEu, LipC1 et LipNC1) ont été ajustés pour que les sorties du modèle correspondent au mieux pour chaque traitements aux résultats mesurés de PV, PVV, PCarcasse chaude et DAT des animaux à l’abattage.
cMEu est le coefficient de l’utilisation de l’énergie pour calculer kME (0,8 < cMEu < 1,2), il correspond à l’efficacité des synthèses, plus cMEu est inférieur à 1, plus l’animal est efficace. LipC1 est le coefficient d’ajustement de la synthèse des lipides dans la carcasse,
LipC1initial est égal 1, plus il est grand, plus de dépôts des lipides dans la carcasse. LipNC1 est le coefficient d’ajustement de la synthèse des lipides dans non-carcasse, LipNC1initial est égal 1, plus il est grand, plus de dépôts des lipides dans non-carcasse.
Tableau 4 : Ingestion moyenne des animaux pendant l’essai
« Digestibilité »
MPEEH ERH MPEEB ERB ESM P-value
Nature Niveau Niveau*Nature MSI (kg/j) 8,03 7,72 7,79 7,30 0,20 0,06 0,11 0,66 MOI (kg/j) 7,56 6,97 7,31 6,56 0,18 <0,01 0,09 0,67 NI (g/j) 188 187 169 175 3,78 0,50 <0,001 0,37 EBI (MJ/j) 146,55 139,94 141,66 131,96 3,61 0,04 0,09 0,67 NDFI (kg/j) 2,30 2,99 2,47 2,97 0,11 <0,001 0,49 0,42 ADFI (kg/j) 1,25 1,61 1,29 1,60 0,10 <0,001 0,78 0,75
Tableau 5 : Pertes fécales, urinaires et de méthane par les animaux
pendant l’essai « Digestibilité »
MPEEH ERH MPEEB ERB ESM P-value
Nature Niveau Niveau*Nature MS fécale (kg/j) 2,13 2,07 2,17 2,06 0,09 0,36 0,88 0,79 MO fécale (kg/j) 1,87 1,69 1,89 1,68 0,09 0,03 0,91 0,86 EB fécale (MJ/j) 39,91 37,43 40,43 37,31 1,82 0,14 0,92 0,86 N fécal (g/j) 63 76 60 74 2,89 <0,001 0,43 0,98 NDF fécal (g/j) 1159 940 1152 956 54,37 0,0011 0,93 0,84 ADF fécal (g/j) 642 528 636 541 28,93 0,0017 0,90 0,76 EB urinaire (MJ/j) 5,65 5,30 5,51 5,14 0,10 <0,01 0,15 0,92 N urinaire (g/j) 51 53 50 55 2,56 0,25 0,80 0,55 EB-CH4 (MJ/j) 9,38 9,74 9,19 9,57 0,61 0,55 0,77 0,99
Tableau 6 : Digestibilité et bilan d’azote
MPEEH ERH MPEEB ERB ESM P-value
Nature Niveau Niveau*Nature
N (%) 66,94 59,55 64,45 58,10 1,04 <0,001 0,07 0,62 MS (%) 73,58 72,84 72,16 71,66 0,69 0,38 0,07 0,86 MO (%) 75,44 75,75 74,15 74,41 0,66 0,67 0,06 0,97 Energie (%) 72,92 73,17 71,53 71,63 0,71 0,81 0,05 0,92 NDF (%) 49,92 68,21 53,32 67,55 1,58 <0,001 0,40 0,21 ADF (%) 46,22 66,70 50,54 65,85 1,68 <0,001 0,31 0,14 Bilan de N (g) 75 59 58 47 3,4326 <0,001 <0,001 0,57 Bilan de N /N digestible ingéré (%) 59,37 52,53 53,95 44,98 2,48 <0,01 0,02 0,67 Bilan de N /N ingéré (%) 39,45 31,32 34,66 26,12 1,52 <0,001 <0,01 0,90
Résultats
1. Expérimantiation de « Digestibilité »
Le tableau 4 montre l’ingestion des animaux de l’expérimentation « Digestibilité ». On peut observer que les animaux alimentés d’ensilage de maïs ont tendance à ingérer plus de MS (P = 0,06) et ont ingéré plus de MO, EB que ceux alimentés d’enrubannage d’herbe (P < 0,05). L’ingestion de NDF et d’ADF pour les traitements d’enrubannage d’herbe est supérieure en comparaison de ceux d’ensilage de maïs. La différence d’ingestion de N entre les deux niveaux alimentaires est significative (P < 0,001).
Selon les résultats des pertes (Tableau 5), les traitements d’enrubannage d’herbe ont plus d’azote, moins de MO, de NDF et d’ADF dans les féces, et moins d’énergie brute dans les urines que leur homologues (P < 0,05), cependant il n’y a pas de différence significative entre les deux niveaux alimentaires pour ces pertes (P > 0,05).
En ce qui concerne la digestibilité des composants chimiques et le bilan d’azote des rations (Tableau 6), les deux traitements d’ensilage de maïs ont la digestibilité d’azote plus importante (7% d’écart pour MPEEH vs ERH ; 6% d’écart pour MPEEB vs ERB), et une digestibilité du NDF et de l’ADF inférieure à l’enrubannage d’herbe (NDF : 18% d’écart pour MPEEH vs ERH, 14% d’écart pour MPEEB vs ERB ; ADF : 20% d’écart pour MPEEH vs ERH, 15% d’écart pour MPEEB vs ERB ). Pour le bilan de N et les deux rapports : Bilan de N/N digestible ingéré et Bilan de N/N ingéré, on peut remarquer une différence significative (P < 0,05) avec MPEE > ER (Bilan de N/N digestible ingéré : 7% d’écart entre les deux traitements Haut, 9% d’écart entre les deux traitements Bas ; Bilan de N/N ingéré : 8% d’écart entre les deux traitements Haut, 9% d’écart entre les deux traitements Bas), Haut > Bas (Bilan de N/N digestible ingéré : 5% d’écart entre les deux traitements MPEE, 8% d’écart entre les deux traitements ER; Bilan de N/N ingéré : 5% d’écart entre les deux traitements MPEE, 5% d’écart entre les deux traitements ER). En plus, les traitements à haut niveau alimentaire ont tendance à avoir une digestibilité de N, MS, MO et d’énergie plus importante que ceux à bas niveau alimentaire (0,05 < P < 0,1) (N : 2% d’écart entre les deux traitements MPEE, 1% d’écart entre les deux traitements ER ; MS : 1% d’écart entre les deux traitements MPEE, 1% d’écart entre les deux traitements ER ; MO : 1% d’écart entre les deux traitements MPEE, 1% d’écart entre les deux traitements ER ; Energie : 1% d’écart entre les deux traitements MPEE,
Tableau 7 : Valeurs nutritionnelles calculées des rations d’après les
résultats mesurés dans l’essai de « Digestibilité »
MPEEH ERH MPEEB ERB
ED (MJ/kg MS) 13,28 13,28 13,00 12,97 EM (MJ/ kg MS) 11,41 11,33 11,11 10,95 UFV (/kg MS) 0,95 0,94 0,91 0,89 PDIN (g/ kg MS) 91,08 104,32 84,29 103,02 PDIE (g/ kg MS) 80,91 108,20 76,91 105,77 PDI/UFV 85,48 111,08 84,50 115,52
Tableau 8 : Ingestion calculée pour l’essai d’« Engraissement » à partir
des valeurs nutritionnelles de rations évaluées en digestibilité
MPEEH ERH MPEEB ERB
MSI totale (kg) 1839 2160 1766 2238
UFV totales ingérées 1736,43 2022,50 1601,88 1988,30 PDI totales ingérées (kg) 147,27 228,12 135,21 229,04 NDF total ingéré (kg) 519,44 837,37 557,03 935,21 MSI ingérée (kg/j) 9,10 9,19 8,29 8,91 UFV (/j) 8,60 8,61 7,52 7,92 PDI (g/j) 729 971 635 913 NDF (kg/j) 2,57 3,56 2,62 3,73 Durée d’engraissement (j) 202 235 213 251
Tableau 9 : Poids et GMQ moyen (en kg et kg/j) aux différentes phases de
l’engraissement et durée d’engraissement (en j)
Sur une même ligne, deux moyennes indicées par des lettres différentes (a, b et c) sont significativement différentes au seuil α = 5%.
2% d’écart entre les deux traitements ER).
Si on compare les valeurs nutritionnelles (Tableau 7) des rations de « Digestibilité » mesurées et calculées à partir des analyses chimiques avec celles estimées (Tableau 3) d’après les tables du livre rouge (INRA, 2007), la valeur UFV (/kg MS) mesurée et calculée augmente légèrement (+2% pour MPEEH, +1% pour ERH, +5% pour MPEEB, +2% pour ERB) pour les 4 rations par rapport à celle estimée. Les valeurs PDIN (g/kg de MS), PDIE (g/kg de MS) et le rapport PDI/UFV mesurées et calculées diminuent pour les traitements d’ensilage de maïs, et augmentent pour ceux d’enrubannage d’herbe en comparaison avec les valeurs estimées [PDIN (g/kg de MS) : - 9 % pour MPEEH, + 4% pour ERH, - 5% pour MPEEB, + 11% pour ERB ; PDIE (g/kg de MS) : - 14% pour MPEEH, + 15% pour ERH, - 11% pour MPEEB, + 22%pour ERB ; le rapport PDI/UFV : - 15% pour MPEEH, + 11% pour ERH, - 16 % pour MPEEB, + 16% pour ERB].
2. Expérimontation d’« Engraissement »
Selon l’ingestion des animaux d’ « Engraissement » (Tableau 8), l’ingestion d’UFV a été au même niveau pour les deux traitements de haut niveau (MPEEH et ERH), et le traitement ERB a eu plus d’ingestion d’UFV que MPEEB, en plus, l’ingestion de PDI pour les traitements ER a été supérieure par rapport à MPEE.
Les résultats de croissance, d’efficacité alimentaire et de composition de la carcasse à l’abattage (Tableaux 11, 12, 13 et 14) viennent de Sepchat et al. (2013). Pour atteindre l’objectif de poids vif moyen de 700 kg, les animaux du traitement ERH ont mis 33 jours de plus que ceux du traitement MPEEH, et ceux de ERB 38 jours de plus que MPEEB. Après une période de faible croissance au début (environ 60 j), les animaux des lots ER ont eu une croissance plus soutenue en fin d’engraissement (Tableau 9). A l’inverse, la croissance des lots MPEE était plus rapide au début, puis moins soutenue en fin d’engraissement. Durant toute la période d’engraissement, la vitesse de croissance pour les animaux d’ERB a été inférieure aux ceux des trois autres traitements. Les objectifs de croissance, 1600 g de GMQ pour les lots hauts et 1400 g pour les lots bas, ont été atteints pour les animaux des traitements d’enrubannage et dépassés pour ceux des traitements d’ensilage de maïs.
Tableau 10 : Différents critères d’efficacité alimentaire
Sur une même ligne, deux moyennes indicées par des lettres différentes (a, b et c) sont significativement différentes au seuil α = 5%.
Tableau 11 : Résultats d’abattage et répartition des tissus adipeux dans la
carcasse
Sur une même ligne, deux moyennes indicées par des lettres différentes (a, b et c) sont significativement différentes au seuil α = 5%.
Tableau 12 : Fixation (abattage-mise en lot) des composants chimiques du
corps au cours de l’engraissement
Sur une même ligne, deux moyennes indicées par des lettres différentes (a, b et c) sont significativement différentes au seuil α = 5%.
Quant à l’efficacité alimentaire (Tableau 10), pour le rapport gain de poids/MSI, les traitements d’ensilage de maïs ont eu une efficacité alimentaire plus importante que ceux d’enrubannage d’herbe en gain de poids par kg de MSI (186 vs. 162 g gain de poids par kg de MSI pour le niveau haut, 191 vs 154 g gain de poids par kg de MSI pour le niveau bas).Le traitement MPEEB a été plus efficace pour le rapport gain/UFV ingérée en comparaison avec les trois autres traitements. Il a aussi eu le rapport protéine fixée/UFV ingérée plus élevé qu’ERB. Si on s’intéresse aux protéines fixées/PDI ingérées, les animaux du traitement MPEEB ont été plus efficaces que les deux lots à base d’enrubannage d’herbe.
Il n’y a pas de différence significative pour le poids vif vide, le poids de carcasse, et le rendement commercial entre les 4 traitements (Tableau 11). Les animaux du traitement MPEEH a déposé significativement plus de tissus adipeux dans le cinquième quartier et plus de tissus adipeux totaux par rapport aux trois autres traitements, et aussi plus de tissus adipeux dans la carcasse que ceux de EHR. Quant à la fixation des composants chimiques du corps (Tableau 12), la même situation a été observée que les animaux qui ont consommé MPEEH ont eu significativement plus de lipides en comparaison avec trois autres traitements. Le traitement de MPEEB a eu de façon numérale plus de lipide dans le corps, plus de dépôts adipeux dans le cinquième quartier et la carcasse que ceux d’ER.
3. Optimisation des paramètres du modèle MECSIC
Comme marqué dans « Matériels et Méthodes », la concentration en énergie métabolisable (CEM) a été calculée comme l’intrant de la simulation. Les 3 paramètres sensibles du modèle (cMEu, LipC1 et LipNC1) (Tableau 13) ont été ajustés afin que les sorties du modèle correspondent au mieux pour chaque traitements aux résultats mesurés de PV, PVV, PCarcasse chaude et DAT des animaux à l’abattage. Les résultats des simulations réalisées avec ces paramètres en comparaison des mesures réelles sont rapportés au Tableau 14. Les PV, PC et DAT sont assez bien ajustées mais on peut noter les écarts relativement élevés de PVV et de PCC pour le traitement MPEEH, l’écart de PVV pour MPEEB, et l’écart de PV pour ERH. Il faudrait donc intervenir sur d’autres paramètres et notamment sur ceux qui modulent le contenu digestif pour avoir une simulation plus précise. Sur la base des 3 paramètres ajustés, on peut noter que les traitements d’ensilage de maïs sont les plus efficaces (cMEu plus faible).Le dépôt de lipides modulé par la valeur de LipC1 est augmenté par rapport au modèle initial pour les traitements d’ensilage de maïs. Ils déposent théoriquement davantage de lipides dans la carcasse, tandis que LipCN1 est diminué par rapport au modèle
Tableau 13 : Les paramètres optimisés
Valeurs des paramètres
Traitement cMEu LipC1 LipNC1
MPEEH 0,88 0,15 0,1
MPEEB 0,88 0,15 0,1
ERH 0,94 0,1 0,07
ERB 0,95 0,1 0,09
Tableau 14 : Comparaison des résultats sur la composition de la carcasse
mesurés à l’abattage et prédits par MECSIC
Durée d’engraissement PV PVV PCC DAT MPEEH Observé 202 708 648 425 91 MECSIC 203 710 634 412 86 Ecart -2 14 13 5 MPEEB Observé 213 694 635 412 80 MECSIC 217 694 619 404 82 Ecart 0 16 8 -2 ERH Observé 235 708 649 419 75 MECSIC 238 720 643 418 80 Ecart -12 6 1 -5 ERB Observé 251 711 640 424 78 MECSIC 250 713 637 418 78 Ecart -2 3 6 0
initial pour les traitements à base d’enrubannage d’herbe. Ces rations induiraient donc moins de lipides dans les viscères.
Discussion
1.
Expérimentation de « Digestibilité »
La différence d’ingestion entre les deux natures de ration de « Digestibilité » est compréhensible, puisque les traitements d’enrubannage d’herbe ont ingéré plus de fibre, qui ont plus d’unité d’encombrement.
Selon Jarrige (1986), l’augmentation du niveau alimentaire a généralement pour effet de diminuer la digestibilité des rations (Robertson et Van Soest, 1975), du fait d’une diminution du temps de séjour des aliments dans le rumen, et/ou d’une baisse du pH qui influence l’activité cellulolytique du jus de rumen, et/ou une excrétion accrue des aliments dans les féces à cause d’une activité amylolytique dans l’intestin grêle insuffisante due à un pH inférieur au pH optimum (Wheeler et Noller, 1977). Mais dans notre étude, les digestibilités de MS, MO, N, énergie, ont tendance à augmenter avec l’augmentation du niveau alimentaire, ce qui n’est pas en accord avec les tendances connues.
La forte digestibilité de l’azote pour les traitements MPEE correspond à une quantité faible d’azote dans les féces. Ceci, plus le fait que le bilan d’azote, ainsi que les deux rapports Bilan de N/N digestible ingéré et Bilan de N/N ingéré ont la même tendance de différence entre les 4 traitements: MPEE > ER, Haut > Bas, signifie que la nature énergétique de la ration « Amidon » augmente l’azote retenu, ainsi que l’efficacité d’utilisation d’azote, par rapport à la ration « Fibre ».
Comme indiqué en haut, la valeur nutritionnelle mesurée des rations est différente de celle estimée à partir des tables (INRA, 2007). La valeur UFV (/kg MS) mesurée et calculée des 4 rations augmente légèrement en comparaison de celle estimée. Malgré ces modifications, la valeur UFV (/kg MS) est identique pour les deux traitements Hauts et pour les deux traitements Bas. Mais l’ingestion d’UFV n’est pas identique comme prévu à cause de la différence de MSI. Les animaux de MPEE ont ingéré plus d’UFV que ceux d’ER pour les deux niveaux alimentaires. Les valeurs PDIN (g/kg de MS), PDIE (g/kg de MS) et le rapport PDI/UFV mesurées et calculées diminuent pour les rations d’ensilage de maïs, et augmentent
pour celles de l’enrubannage d’herbe par rapport aux valeurs estimées. L’ingestion de PDI n’est plus identique entre les traitements : MPEEB < MPEEH < ERB < ERH.
2.
Expérimentation d’« Engraissement »
L’ingestion d’UFV par jour est au même niveau entre MPEEH et ERH, tandis que ERB ingère plus d’UFV par jour que MPEEB. L’ingestion de PDI par jour a la même tendance que celle de l’essai de « Digestibilité » : MPEEB < MPEEH < ERB < ERH.
Les animaux des traitements ER ont mis plus de temps pour atteindre l’objectif de poids vif moyen de 700 kg que ceux des MPEE. Cela correspond à ce que Schoonmaker et al. (2003) ont observé, pour atteindre le même objectif d'épaisseur du gras (1,27 cm), les bœufs ayant reçu une ration ad libitum à haut niveau de fourrage durant la période de croissance ont mis plus de temps que ceux nourris avec une ration ad libitum à haut niveau de concentré même s’ils ont tous reçu la même ration riche en concentré pendant la période d’engraissement.
La diminution de la vitesse de croissance pour les traitements d’ensilage de maïs peut résulter du fait que les animaux s’approchent de l’état de maturité selon la courbe de croissance Gompertz, ou de l’acidose subaiguë durant une longue période quand les animaux sont alimentés avec une ration riche en amidon. Pour les niveaux Hauts, le traitement de MPEEH a ingéré autant d’UFV, mais moins de PDI par jour que celui de l’ERH, mais il n’y a pas de différence de vitesse de croissance durant l’engraissement. Pour les niveaux Bas, même si les animaux de MPEEB ont eu moins d’ingestion d’UFV et de PDI par jour en comparaison avec ceux d’ERB, ils ont la vitesse de croissance supérieure.
En ce qui concerne le rapport gain de poids/MSI, Schoonmaker et al. (2003) ont remarqué que les bœufs qui ont ingéré ad libitum une ration riche en amidon ont eu pas de différence significative de GMQ pendant la période de croissance, mais ont été 27% plus efficaces et ont moins de MSI durant la période d’engraissement par rapport aux ceux nourris
ad libitum une ration riche en fibre. Mais Retallick et al. (2010) n’a noté aucune différence du
rapport gain de poids/MSI avec la diminution du niveau d’amidon dans les rations durant la période de croissance, par contre, le rapport s’est diminué linéairement avec la diminution du niveau d’amidon pendant l’engraissement. La meilleure efficacité alimentaire observée avec l’ensilage de maïs dans notre étude par comparaison à l’enrubannage d’herbe est compatible
avec Keady et al. (2013), Schoonmaker et al. (2004, 2003), Nuernberg et al. (2005) et Steen et Kilpatrick (2000).
La différence de poids vif vide et de poids de carcasse entre les 4 traitements n’a pas été significative, ce qui semble logique puisque les animaux sont abattus lorsque les animaux des 4 traitements avaient le même poids vif moyen.
Si on s’intéresse à l’effet de la nature de la ration sur la composition de la carcasse, il y a beaucoup résultats qui sont comparables(Costa et al., 2013 ; Keady et al., 2013 ; Micol et
al., 2007 ; Schoonmaker et al., 2004, 2003 ; Geay et al.,1997 ; Colman et al., 1995). Costa et al. (2013) ont construit des rations approximativement iso-énergétiques et iso-azotées en
changeant les proportions d’ensilage de maïs et de concentré dans la ration. Ils ont observé que le lot alimenté à haut niveau de concentré a déposé plus de tissus adipeux dans la carcasse et dans les viscères en comparaison de celui à haut niveau d’ensilage de maïs. Keady et al., (2013) ont également marqué que l’ensilage de maïs a une efficacité de stockage d’énergie à la carcasse plus forte qu’ensilage d’herbe. Ces résultats sont en accord avec le notre pour les niveaux Hauts. Les animaux MPEEH ont quand même déposé autant de protéines (même si l’ingestion quotidienne et totale de PDI des lots MPEE était inférieure à celle des lots ER, Tableau 6) et plus de tissus adipeux dans la carcasse et le cinquième quartier par rapport au celui de ERH en consommant la même quantité d’UFV ingérée par jour, et moins d’UFV totale durant la période d’engraissement (durée d’engraissement plus courte). Pour les niveaux Bas, les animaux du traitement MPEEB a eu la même quantité de dépôts protéiques et adipeux dans la carcasse et le cinquième quartier que ceux des traitements ER malgré une ingestion quotidienne et totale de PDI et surtout d’UFV plus faible. Cela signifie que l’ensilage de maïs permet une meilleure utilisation de l’énergie et de l’azote pour déposer des lipides et des protéines que l’enrubannage d’herbe. Par rapport à l’effet du niveau alimentaire, notre étude a mis en évidence que les différents niveaux alimentaires influaient davantage dans les traitements d’ensilage de maïs qu’enrubannage d’herbe. Les animaux du traitement MPEEH a eu plus de dépôts adipeux dans le cinquième quartier et de dépôts adipeux totaux que MPEEB, tandis que les traitements d’ER ont déposé la même quantité de tissus adipeux. Par contre, Schoonmaker et al.(2003) ont signalé que les bœufs programmés d’avoir 1,2kg/j de GMQ ont dépôsé plus de protéines que ceux programmés d’avoir 0,8kg/j de GMQ. Ce n’est pas le cas dans notre étude. Le désaccord peut prévenir de l’écart de niveau alimentaire (1,6 et 1,4 kg/j pour notre étude vs 1,2 et 0,8kg/j pour celle de Schoonmaker et al.(2003)).
3.
Dynamique des dépôts et ajustement du modèle MECSIC
L’ajustement du modèle a nécessité de moduler le paramètre aMW. Celui-ci intervient dans le calcul de l’efficacité de dépôt de l’énergie métabolisable disponible. Ceci est toujours le cas lorsque l’on applique ce modèle à des données ainsi que l’avait montré (Ferard et al 2012). Les valeurs différentes selon les lots de ce paramètre sensible reflètent les écarts de valorisation métabolique des rations. Les meilleures s’observent avec les rations « Maïs » riches en amidon. Les deux autres paramètres LipC1 et LipNC1 sont liés aux synthèses maximales des lipides dans la carcasse et non carcasse. Leurs ajustements avaient aussi été nécessaires lorsque le modèle s’était confronté à des données indépendantes sur bœufs Angus disponibles à l’Université de Davis (Garcia et al., 2008).
Selon les résultats de cette étude, les paramètres du modèle MECSIC reflétant l’efficacité et le métabolisme des lipides devraient donc pouvoir s’ajuster lorsqu’on applique ce modèle dans des conditions différentes de nature de la ration. L’évolution du modèle devrait donc prendre en compte ces différences et les intégrer complètement à l’avenir dans le schéma (nouvelles variables, nouvelles relations.).
Conclusion et perspective
La présente étude vise à évaluer l’effet de la nature de la ration et du niveau alimentaire sur la composition de carcasse et à optimiser les paramètres du modèle MECSIC. Les analyses chimiques des aliments et l’essai de « Digestibilité » dans notre étude ont permis de mesurer les valeurs nutritionnelles des rations, qui sont différentes de celles estimées à partir des tables des aliments (INRA, 2007), surtout pour PDIN et PDIE. Nous avons obtenu les résultats intéressants sur les valeurs nutritionnelles des rations, qui permettent d’interpréter les résultats de la croissance, de la composition corporelle et des dépôts de l’essai « Engraissement ». Quant à l’optimisation du modèle MECSIC, faute de temps, nous avons réalisé les simulations du modèle à base des données pour chaque lot, il vaudra mieux de le faire pour chaque animal, cela permettrait d’améliorer la précision des simulations.
L’ensemble de cette étude nous permet de conclure que la nature de la ration a une influence notable sur les dépôts adipeux. Durant l’engraissement chez les jeunes bovins, les animaux alimentés avec de lensilage de maïs déposent plus de tissus adipeux dans la carcasse et non-carcasse par rapport aux ceux nourris d’enrubannage d’herbe en ingérant le même niveau d’UFV. L’effet de différent niveau alimentaire sur les dépôts adipeux pour l’ensilage de maïs est plus remarquable que l’enrubannage d’herbe.
Dans l’avenir, un indicateur quantitatif permettant de définir et distinguer la nature de la ration et induire des paramètres appropriés pourra être pris pour faire évoluer le modèle MECSIC. La validation du modèle de croissance sur des données mesurées devrait permettre de construire un outil utilisable dans la pratique d’élevage. Cet outil permettra aux éleveurs de prédire à l’avance le poids de carcasse, la répartition des dépôts adipeux etc… de leurs animaux en fonction de la ration ingérée, et d’ajuster au mieux la ration pour une production plus rentable selon la demande du marché et du prix des aliments.
Références bibliographiques
BALCH C.C.,1967. Problems in predicting the value of non-protein nitrogen as a substitute for protein in rations for farm animals. World Rev. Anim. Prod., 3, 84-91.
COLMAN S.W., GALLAVAN R.H., WILLIAMS C.B., PHILLIPS W.A., VOLESKY J.D., RODRIGUEZ S., BENNETT G.L., 1995. Silage or limit-fed grain growing diets for steers : I. Growth and carcass quality. 73:2609-2620.
COLMAN S.W., GALLAVAN R.H., PHILLIPS W.A., VOLESKY J.D., RODRIGUEZ S., 1995. Silage or limit-fed grain growing diets for steers : II. Empty and carcass composition. 73:2621-2630.
COSTA A.S.H., COSTA P. BESSA R.J.B., LEMOS J.P.C., SIMOES J.A., SANTOS-SILVA J., FONTES C.M.G.A., PRATES J.A.M., 2013. Carcass fat partitioning and meat quality of Alentejana and Barrosa young bulls fed high or low maize silage diets. 93: 405-412.
FERARD A., GARCIA-LAUNAY F.,CABON G., BASTIEN D., AGABRIEL J., 2012, Variabilité de l’efficacité d’utilisation de l’énergie ingérée chez les jeunes bovins charolais : mise en évidence d’une évolution interannuelle. Renc. Rech. Ruminants, 19, 298.
FRANCE J., THORNLEY J.H.M., 1984. Mathematical models in agriculture. Butterworths et Co, London, UK, 335p.
GARCIA F., SAINZ R.D., AGABRIEL J., BARIONI L.G., OLTJEN J.W. 2008. Comparative analysis of two dynamic mechanistic models of beef cattle growth. Animal Feed Science and Technology. 143: 220-241.
GEAY Y., PICARD B., JAILLER R.D., JAILLER R.T., LISTRAT A., JURIE C., BAYLE M.C., TOURAILLE C., 1997. Effets de la nature de la ration sur les performances, les caractéristiques musculaires et la qualité de la viande de taurillon salers. Renc. Rech. Rum., 4, 307-310.
GRAUGNARD D.E., PIANTONI P., BIONAZ M., BERGER L.L., FAULKNER D.B., LOOR J., 2009. Adipogenic and energy metabolism gene networks in longissimus lumborum during rapid post-weaning growth in Angus and Angus*Simmental cattle fed high-starch or low-starch diets. BMC Genomics, 10:142.
HENRICKSON R.L., POPE L.S., HENDRICKSON R.F., 1965. Effect of rate of gain of fattening beef calves on carcass composition. Anim. Scien., 24, p: 207-513.
HOCH T., JURIE C., PRADEL P., CASSAR-MALEK I., JAILLER R., PICARD B., AGABRIEL J., 2005. Effects of hay quality on intake, growth path, body composition and muscle characteristics of Salers herfers. Anim. Res. 54, 241-257.
HOCH T., AGABRIEL J., 2004a. A mechanistic dynamic model to estimate beef cattle growth and body composition: 1. Model description. Agrie. Syst., 81, 1-15.
HOCH T., AGABRIEL J., 2004b. A mechanistic dynamic model to estimate beef cattle growth and body composition: 1. Model evaluation. Agrie. Syst., 81, 17-35.
HOCH T., PRADEL P., AGABRIEL J., 2004. Modélisation de la croissance de bovines : évolution des modèles et applications. INRA Prod. Anim., 17(4), 303-314.
HORN G.W., CRAVEY M.D., MCCOLLUM F.T., STRASIA C.A., KRENZER E.G.,CLAYPOOL J., CLAYPOOL P.L., 1995. Influence of high-starch vs high-fiber energy supplements on performance of stocker cattle grazing wheat pasture and subsequent feedlot performance. J ANIM SCI, 73:45-54.
INRA, 2007. Alimentation des bovins, ovins et caprins. Besoins des animaux. Valeurs des aliments Tables INRA.
INRA, 1986. Production de viande bovine, p35, 45-46, 54.
ISO 5983-1:2005. Animal feeding stuffs -- Determination of nitrogen content and calculation of crude protein content -- Part 1: Kjeldahl method.
INSTITUT DE L’ELEVAGE, 2011. Dossier Economie de l’Elevage numéro 245 : Dynamique de la production de jeunes bovins. 13- 32.
KEADY T.W.J., GORDON A.W., MOSS B.W., 2013. Effects of replacing grass silage with maize silages differing in inclusion level and maturity on the performance, meat quality and concentrate-sparing effect of beef cattle. Aniaml, 7:5, pp 768-777.
Manuel d’utilisation IKA Calorimeter system C200; EN 61010; EN 50082; EN 55014; EN 60555.
MICOL D., DUBROEUCQ H., MARTIN C., GARCIA F., MIALON M.M., AGABRIEL J., 2007. Utilisation de rations de valeurs contrastées pour la finition de jeunes bovins de race Blond d’Aquitaine. Renc. Rech. Rum.,14, 233-236.
MICOL D., ROBELIN J., GEAY Y., 1993: Composition corporelle et caractéristiques biologiques des muscles chez les bovins en croissance et à l’engrais. INRA Prod. Anim., 6(1), 61-69.
NF ISO 6496, 2011. Aliments des animaux - Détermination de la teneur en eau et en d'autres matières volatiles.
NF V18-122 Août 1997. Aliments des animaux - Détermination séquentielle des constituants pariétaux -Méthode par traitement aux détergents neutre et acide et à l'acide sulfurique.
NUERNBERG K., DANNENBERGER D., NUERNBERG G., ENDER K., VOIGT J., SCOLLAN N.D., WOOD J.D., NUTE G.R., RICHARDSON R.I., 2005. Effect of a grass-