Genomic determinants of the efficiency of internal ribosomal entry sites of viral and cellular origin
8
0
0
Texte intégral
Figure

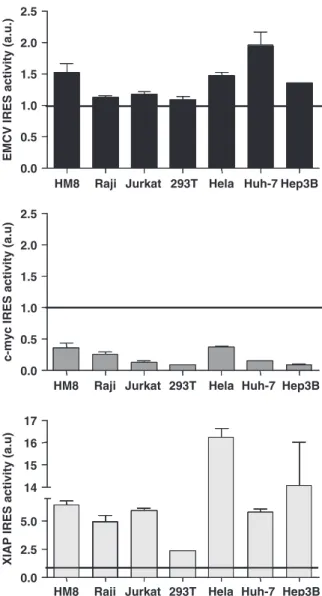

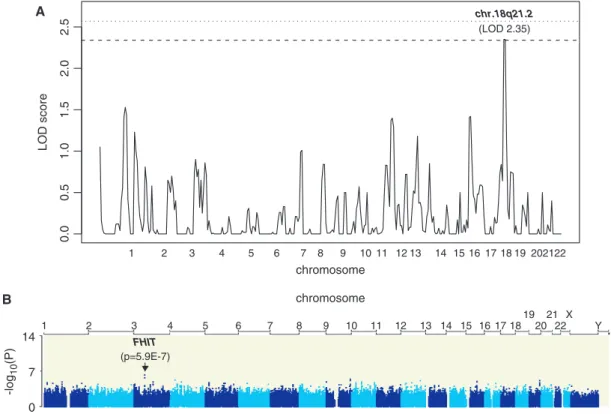
Documents relatifs
