Identification of a CYP3A form (CYP3A126) in fathead minnow ( Pimephales promelas ) and characterisation of putative CYP3A enzyme activity
Texte intégral
Figure
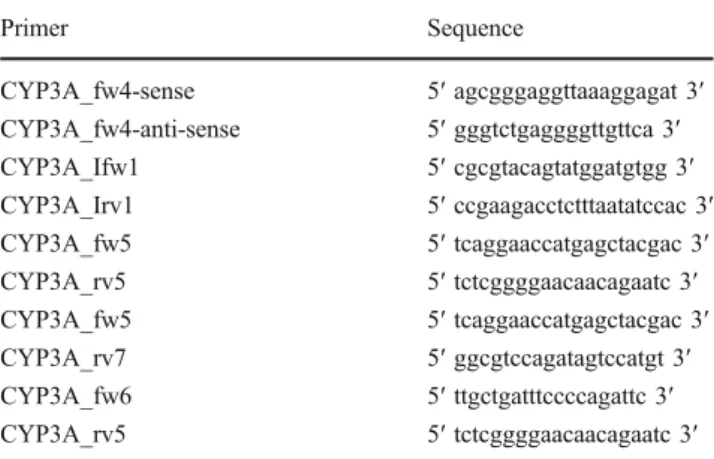


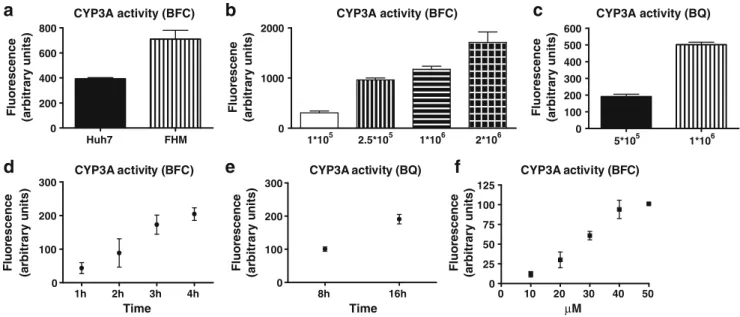
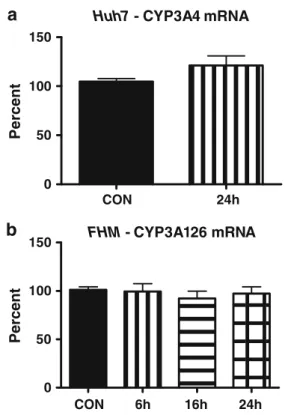
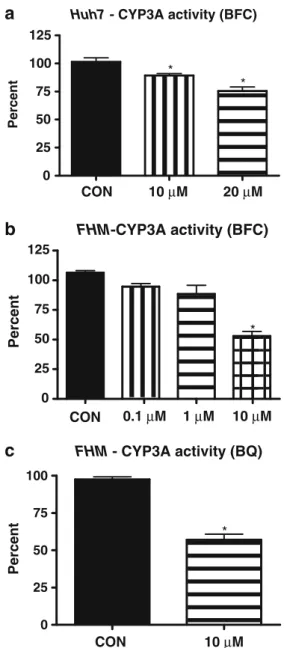
Documents relatifs
Therefore, the present study was designed to characterize the expression of plasma membrane nucleoside transporters, i.e., ENT1, ENT2, CNT1, CNT2 and CNT3, in HepaRG cells, as well
We found that in addition to the critical conserved and non-conserved amino acids within the VP1u PLA2 motif, amino acid residues outside the VP1u PLA2 motif are also important for
( ّنإ ﳚ جﺮــﺨﳌا ﺚﻳﺪــﺣ ّﺮ ا ﺎــﳍ دروأ ﺔــﻣﺎﻫ ﺔــﻴﺋﺎﻨﺛ ﰲ ضﻮــﳋا ﱃإ ﺎــﻧ ىﺮــﺧﻷا ﻲــﻫ ﲏــﺟ ﻦــﺑ تﻼﻴــﻠﻌﺗ ، تﻮﺼﻟا ﺔﻴﺋﺎﻨﺛ ﻲﻫو ﻻأ و فﺮﳊا. فﺮﺤﻟاو تﻮﺼﻟا :
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des
Besides the reduced fecundity and the clear VTG induction, in male fish the significant decrease of the number of tubercles was another clear indication for the estrogenic activity
In the present study, possible breed-, gender- and species-differences in P4503A (the P450 subfamily more expressed in the human liver) expression were studied in vitro in
Dans ce travail, le cadre de référence et les concepts en lien avec le thème sont définis tout en s’intéressant aux impacts qu’ont les soins infirmiers pédiatriques à
