J. Membrane Biol. 193, 23-34 (2003) DOI: 10.1007/s00232-002-2004-6
The Journal of
Membrane
Biology
9 Springer-Verlag New York Inc. 2003
Gating Properties of a Sodium Channel with Three Arginines Substituted by Histidines
in the Central Part of Voltage Sensor $4D4
F.J.P. Kiihn, N.G. Greeff
Physiologisches Institut, Universitfit Ztirich, Winterthurerstr. 190, CH-8057 Ztirich, Switzerland Received: 2 October 2002/Revised: 21 January 2003
Abstract.
In voltage-dependent sodium channels there is some functional specialization of the four different $4 voltage sensors with regard to the gating process. Whereas the voltage sensors of domains 1 to 3 control activation gating, the movement of the voltage sensor of domain 4 ($4D4) is known to be tightly coupled to sodium channel inactivation, and there is some experimental evidence that $4D4 also participates in activation gating. To further explore its putative multifunctional role in the gating process, we changed the central part of $4D4 in rat brain IIA (rBIIA) sodium channels by the simultaneous re- placement of the third (R1632), fourth (R1635) and fifth (R1638) arginine by histidine (mutation R3/4/ 5H). As a result, the time course of current decay observed in R3/4/5H was about three times slower, if compared to wild type (WT). On the other hand, the recovery, as well as the voltage dependence of fast inactivation, remained largely unaffected by the mu- tation. This suggests that at physiological pH (7.5) the effective charge of the voltage sensor was not significantly changed by the amino-acid substitutions. The well-known impact of site-3 toxin (ATX-II) on the inactivation was drastically reduced in R3/4/5H, without changing the toxin affinity of the channel. The activation kinetics of WT and R3/4/5H studiedat low temperature (8~ were indistinguishable,
while the inactivation time course of R3/4/5H was then clearly more slowed than in WT. These data suggest that the replacement of arginines by histidines in the central part of $4D4 clearly affects the move- ment of $4D4 without changing the activation kinetics.
Correspondence to: N.G. Greeff; email: [email protected]
Abbreviations: rBIIA, rat brain IIA; TEVC, two-electrode voltage clamp; Rs, series resistance; TTX, tetrodotoxin; IN~,, sodium cur- rent; MTSET, 2-trimethylammonium-ethyl-methanethiosulfonate; ATX-II, sea anemone toxin.
Key words:
Voltage sensor $4D4 - - Mutagenesis - -ATX-II - - Fast inactivation Activation
Introduction
Voltage-dependent sodium channels are character- ized by four distinct positively charged transmem- brane segments that serve as sensors for changes of the membrane potential (Catterall, 1986; Noda et al., 1986; Stfihmer et al., 1989; Yang and Horn, 1995). Besides, there are two kinetically defined types of gates for the control of channel activation and fast inactivation, respectively (Sigworth, 1993; Yellen, 1998). However, the choreography of interactions between voltage sensors and gates necessary for the opening and closure of the channel pore until now is poorly understood. The systematic analysis of sodi- um channels containing point mutations in either of the four $4 segments reveals that in particular the voltage sensor of domain 4 ($4D4) is involved in fast inactivation gating, whereas the S4-segments of do- mains 1-3 are mainly coupled to the activation pro- cess (Chahine et al., 1994; Chen et al., 1996; Kontis & Goldin, 1997; Kontis, Rounaghi & Goldin, 1997; Cha et al., 1999; Ktihn & Greeff, 1999). In a previous study we proposed a model where $4D4 moves in two separate steps with an initial step or delay perhaps involved in activation and a second step that controls fast inactivation (Ktihn & Greeff, 1999). In line with this hypothesis are the recent findings of Horn, Shinghua and Gruber (2000), which indicate that the movement of $4D4 is a two-step process, each step coupled to a different gate. On the other hand, the data of Sheets and Hanck (1995), Sheets et al. (1999) and Sheets, Kyle and Hanck (2000) suggest that the movement of $4D4 is selectively inhibited in the presence of site-3 toxins. As a result, the available gating charge was reduced by about one third and
24 F.J.P. Kfihn and N.G. Greeff: $4D4 Movement and Sodium Channel Gating
sodium channel fast inactivation kinetics was drasti- cally changed. However, corresponding effects on activation gating were not noticed by these authors.
To further elucidate the putative role of $4D4 in sodium channel gating, we continued our earlier strategy to disturb the mobility of this voltage sensor by the replacement of arginines by histidines. In ad- dition to the substitutions used so far, R1635H and R1638H and the double mutation R1635H/RI638H
(see
Kfihn & Greeff, 1999), we added R1632H and could successfully express the triple mutation. Thereby, we extended the histidine modification fur- ther towards the narrow pore that is regarded to represent that part of the channel protein where the actual voltage drop and voltage sensing occur (Yang, George & Horn, 1996). In this way, we wanted to inhibit further or freeze the movement of $4D4. Of special interest was also to inquire whether in addi- tion to the slowing of inactivation we would now observe a slowing or stop of activation, i.e., to search for evidence of an involvement of $4D4 in activation. Experiments were carried out also at low temperature for better temporal resolution and furthermore, we analyzed the impact of the known modifier of $4D4 mobility, the site-3 toxin ATX-II, on channel gating. The results strongly support earlier findings that the mobility of $4D4 determines the fast inactivation kinetics while we still found no convincing evidence for an involvement of this voltage sensor in activation gating.Materials and Methods
MUTAGENES1S AND EXPRESSION OF CHANNELS The cDNAs of both the wild-type rat brain IIA (rBIIA) sodium channel et subunit and the [~l subunit used in this study were de- rived from plasmid pVA2580 and pBluescript SK, respectively, and transferred into high-expression vector pBSTA. The resulting plasmids pBSTA(~) and pBSTA(]3) contain a T7 RNA polymerase promotor and Xenopus-[J-globin 5' and 3' untranslated sequences
that greatly increase the expression of exogenous proteins in oocytes (Shih et al., 1998). Site-directed mutagenesis was performed as previously described (Kfihn & Greeff, 1999). The presence of inadvertent mutations in other regions of the channel could be excluded because several mutant clones tested ~Jie]de~ ~he same results. In addition, the mutations were verified by DNA sequenc- ing. Capped, full-length transcripts were generated fro n 1 SacH
linearized plasmid DNA using T7 RNA polymerase (mMessage mMachine In Vitro Transcription Kit from Ambion, Austin, TX). Large oocytes (stage V-VI; 1.2 mm diameter on average) from
Xenopus laevis (NASCO, Ft. Atkinson, WI) were used. For surgical
removal of the oocytes, female frogs were anaesthetized with 0.15% tricaine (3-aminobenzoic acid ethyl ester; Sigma, St. Louis, MO) and placed on ice. Frogs were allowed to recover at least 6 mouths following surgery. All animal handling was carried out in accord- ance with methods approved by Swiss Government authorities.
One day before injection of complementary RNA (cRNA), the oocytes were defolliculated in a Ca2+-free solution containing 2 mg/ml collagenase (Boehringer, Mannheim, Germany), for ~2 h at
room temperature. Oocytes were microinjected with 50 ng cRNA of WT or mutant ~-subunit a n d - - i f coexpression was needed--100 ng of ]3-subunit in a 1:1 volume of 50 to 100 nl, and maintained at 18 + I~ in Modified Barth's Solution (MBS, in mM) 88 NaCI, 2.4 NaHCO3, 1 KC1, 0.82 MgS04, 0.41 CaCI2 0.33 Ca(NO3)2, 10 HEPES-CsOH, pH 7.5, supplemented with 25 U penicillin, 25 lag/ ml streptomycin-sulfate and 50 lag/ml gentamycin-sulfate. For the partial reduction of the large WT ionic current, TTX (RBI-Re- search Biochemicals International, Natick, MA) was added to the recording chamber. Sea anemone toxin (ATX-II) from Anemonia sulcata (Sigma) was used in concentrations between 0.13 JaM and 2 p.M.
Allowance for distribution of ATX-II in the bath and towards the oocyte membrane of 5 minutes was found sufficient, since no further increase of the effect was observed after 10 and 15 minutes. ELECTROPHYSIOLOGY AND DATA ACQUISITION Two-electrode voltage-clamp recordings were performed 2 to 6 days after cRNA-injection with a TEC-05 (npi-electronic, Tamm, Germany) that had been modified for improved compensation of the series resistance (R~) and for fast charging of the membrane capacitance (Greeff & Polder, 1998; Greeff & Kfihn, 2000). In- tracellular agarose cushion electrodes (Schreibmayer, Lester & Dascal, 1994) were filled with 3 M KCI and had a resistance be- tween 50 and 150 k~. Macroscopic INa and Ig signals were recorded using a PDP-11/73 computer (Digital Equipment, Maynard, MA) controlling a 16-bit A/D and 12-bit D/A interface (CED, Cam- bridge, UK). The oocytes were clamped at a holding potential of 100 mV for at least 5 minutes to ensure recovery from slow in- activation before recording started. The experiments were done at temperatures of + 15 or + 8 • I~ controlled by a Peltier element. Rs compensation was adjusted critically to accelerate the senling time of the capacitance transients within 200 p.s (without low pass filtering, see below). No analog subtraction was used, since the 16- bit ADC had a sufficiently fine resolution for digital subtraction of the linear transient and leak currents by scaled averages from pulses between -120 mV and -150 mV. Reduction of the re- maining asymmetry was achieved by finding a compromise between clamping speed and asymmetry, i.e., by low-pass-filtering the command signal at 5 kHz (8-pole Bessel). Signals were low-pass filtered at 5 kHz ( - 3 dB) and sampled at 10 or 20 kHz. The actual clamp speed at the oocyte membrane was determined from the integrated capacitance transient to have a time constant between 150 to 200 Its. Rs errors were <5 mV unless the currents exceeded ~20 30 laA. Data analysis was performed on the PDP-11 and ad- ditionally with the Windows-compatible programs UN-SCAN- IT T M (Silk Scientific, Orem, UT) and PRISM T M (GraphPad Software, San Diego, CA).
]~esults
EXPRESSION AND VOLTAGE-DEPENDENT GATING
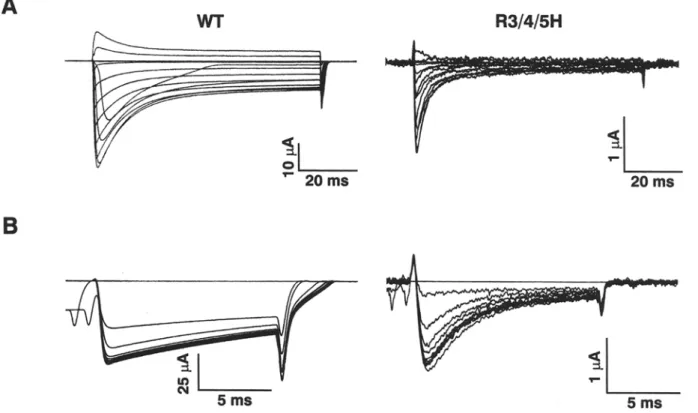
Figure 1A shows the macroscopic sodium currents of WT and triple mutant R1632H/R1635H/R1638H (subsequently denoted as R3/4/5H, since third to fifth arginines of voltage sensor $4D4 were changed to histidines). Despite of using a high-expression system,
(see
Methods) that usually produces sodium currents of 50-100 ~tA and gating currents of 5-10 ~A for WT and other $4D4 mutants in our lab (Kfihn & Greeff, 1999), the mutant R3/4/5H consistently showed a decreased expression level (maximum INa of 3-4 ~tA)F.J.P. Kfihn and N.G. Greeff: $4D4 M o v e m e n t and Sodium Channel Gating 25
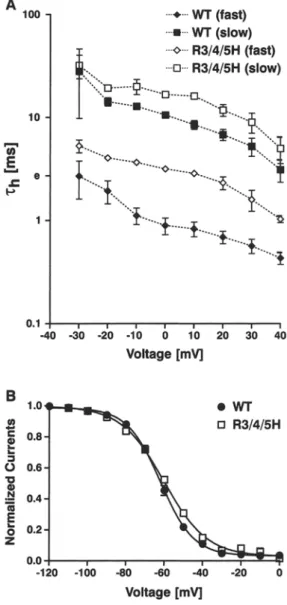
Fig. 1. Expression level and macroscopic Na + currents. (A) Na + currents from WT- and R3/4/5H- injected Xenopus oocytes were elicited by step-depolarizations of - 8 0 mV to + 80 mV, from a holding potential o f - 1 0 0 inV. In order to minimize possible Rs- distortions, the large W T sodium currents were reduced to 5-10 gA by application of TTX. The m u t a n t R3/4/5H produces only sodi- u m currents smaller than 4 gA and hence currents were not sig- nificantly disturbed by R~. (B) Corresponding current-voltage relationship o f peak Na + currents, normalized to the largest in- ward current. The data (mean • SEN of n = 3 cells) were fitted to the equation Y - G . ( X - VNa)/(1 + exp((VM X)/kv)) 3 where VNa is the reversal potential o f N a +, VM is the half-activation potential and kv, a slope factor. For WT, VNa - 50.6 • 1.2 mV, VM = 36.9 + 0.7 mV and kv - 7.04 • 0.7 mV and for R3/4/ 5H, Vy~ = 55.6 + 0.8mV, VM = 29.6 • 0 . 6 m V a n d k v - 9.6 • 0.6 mV.
that was at least one order of magnitude below the corresponding WT level. Likewise, the gating cur- rents of R3/4/5H were too small (<0.5 gA) to be unequivocally separated from asymmetry artifacts and baseline distortions, respectively. Nevertheless,
Fig. 2. Voltage independence of fast inactivation. (A) Semiloga- rithmic plot of the voltage dependence o f the inactivation time constants, rh~r~st~ and rh(slow~. The data (mean i SEM o f n = 4 cells) were obtained from current traces elicited between - 3 0 and + 4 0 mV from a holding potential o f - 1 0 0 m V (as shown in Fig. IA) and fitted to a double exponential of the form Y - (1 - exp (--t/Zm)) 3 ' [A1 9 exp(--t/rhl) + A 2 . e x p ( - t / r h 2 ) + A3]. (B) Steady- state inactivation at 0 mV induced by a 100 ms prepulse at voltages between - 1 2 0 mV and 0 mV in increments of 10 mV at a holding potential of - 1 0 0 mV. The curves are fitted to a standard Boltz- m a n n distribution of the form I/Ima• = 1/(1 + exp[( V - V05)/kv]). The half-maximal inactivation (V05) was - 6 2 • 0.4 m V for W T and - 6 0 . 2 • 0.6 mV for R3/4/5H. The slope factor (k~) of the curve was - 8 . 7 0 • 0.4 mV for W T and - 1 1 . 3 i 0.5 mV for R3/4/ 5H. Values are m e a n • SEN o f n = 5 cells.
the corresponding sodium current was large enough for the characterization of the gating kinetics of R3/ 4/5H.
In order to minimize possible Rs-distortions, the large WT currents were reduced by application of about 100 nM TTX to below 10 gA, as shown in Fig. 1A. The scaled current-voltage plots of peak currents
26 F.J.P. Kiihn and N.G. Greeff: $4D4 M o v e m e n t and Sodium Channel Gating
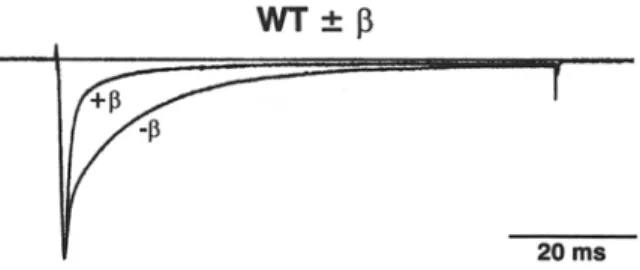
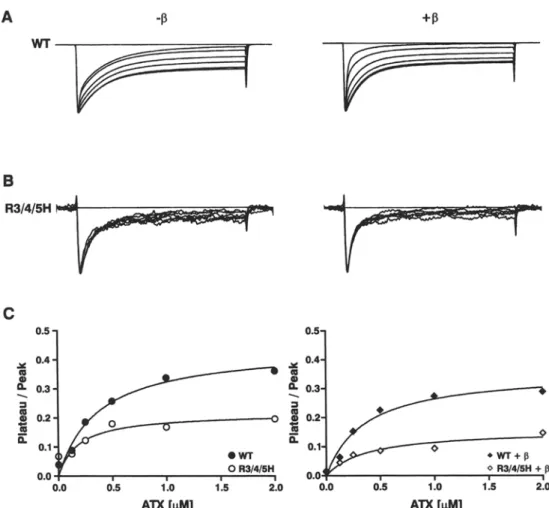
Fig. 3. Bimodal gating and effects o f the accessory [31 subunit on the inactivation kinetics. Typical current traces of W T and R3/4/ 5H elicited at 0 m V from a holding potential of - 1 0 0 mV with or without [31 coexpression; test pulse duration 80 ms. Traces were normalized to peak. Note the pronounced bimodal gating behavior o f W T with the drastic shift to the fast gating mode induced by [31
that is not observed in R3/4/5H. The corresponding inactivation time constants are given for 3 similar experiments each for W T and m u t a n t in the text. Functional differences due to limiting a m o u n t s of [31 subunits in the cells could be excluded because an excess o f [31 c R N A was coinjected.
Table 1. Time constants o f INa-decay of W T and R3/4/5H at different bath temperatures
Voltage W T (T = 15~ rh (fastmow) W T (T = 8~ rh (f~.~t/slow~ R3/4/5H (T = 15~ rh (fastmowl R3/4/5H ( T - 8~ rh (f~st/slow/ 20 mV 1.94 + 0.8/14.2 + 2.3 3.09 • 0.9/19.5 • 0.3 4.05 • 0.4/19.3 • 2.0 14.3 • 0.9/212 • 77
0 mV 0.89 • 0.3/10.6 =k 1.2 1.92 • 0.01/15.3 + 1.8 3.17 • 0.2/16.7 • 0.7 10.4 • 1.1/59 • 7.6 20 mV 0.69 • 0.2/6.83 • 1.4 1.39 + 0.1/11 • 0.6 2.32 • 0.7/11.8 • 3.2 8.6 + 0.4/54.1 + 12.4 The data (mean • SD of n -- 3 cells) were obtained as described in the legend o f Fig. 2.
obtained from WT or R3/4/5H and fitted as de- scribed in the figure legend suggest a slight depolar- izing shift of ~7 mV for the mutant (Fig. 1B). This shift might be due to some still uncompensated Rs- distortion, which would shift the
I/V-curve
of WT to more hyperpolarizing potentials. As discussed in context of Fig. 8, a distortion of peak currents and hence thel/V-curve,
is also likely due to changes in inactivation kinetics.T I M E C O U R S E OF F A S T INACTIVATION
The kinetics of current decay was analyzed by per- forming double-exponential fits from normalized current traces elicited between - 3 0 mV and + 40 mV (Fig. 2A). In the absence of the [31-subunit, rat brain IIA (rBIIA) sodium channels show a pronounced bimodal gating if expressed in
Xenopus
oocytes, as already recognized for neuronal and skeletal muscle sodium channels (e.g., Isom et al., 1992; Patton et al., 1994). As described in the context of Fig. 3, we noted that coexpression of the 131-subunit exerts different effects on the inactivation gating of WT and R3/4/5H and, therefore, we compared the inactivation time courses without [31 coexpression. The fast gating channels of WT inactivated approximately ten times faster than the corresponding slow-mode channels, which result is in agreement with data obtained from other groups (Moorman et al., 1990; Zhou et al., 1991; Ji et al., 1994; Moran et al., 1998). If compared to WT, the current decay of R3/4/5H also showed a slow and a fast inactivating phase(see
Fig. 1A), as verified by the double-exponential fit (Fig. 2A). Thefast-mode kinetics determined at 15 ~ bath tem- perature was about three times slower in R3/4/5H than observed in WT, whereas the corresponding time constants of the slow mode were only about 1.5- fold slower in the mutant (Fig. 2A and Table 1). For the most significant voltage range between - 2 0 mV and + 40 mV, the voltage dependence of fast inacti- vation of both kinetic populations was determined as an e-fold decrease within 74 mV for WT and 79 mV for R3/4/5H (Fig. 2A). Likewise, the voltage depen- dence of steady-state inactivation was slightly de- creased in the mutant, but the midpoint of the curve was almost identical in WT and R3/4/5H (Fig. 2B). Therefore, similar to the previously described muta- tions R4H, R5H and R4/5H (Kfihn & Greeff, 1999), the triple mutation R3/4/5H slowed the fast in- activation kinetics and, in addition, moderately decreased the voltage dependence of channel in- activation.
COEXPRESSION OF THE ~ I - S U B U N I T
Hitherto, the effect of the accessory 131 subunit on the gating kinetics of ~x subunits expressed in
Xenopus
oocytes were primarily studied in WT sodium chan- nels (for review
see
Isom, De Jongh & Catterall, 1994) and in sodium channels containing mutations that are located outside of the $4 voltage sensors (e.g., Balser et al., 1996; Moran et al., 1998; Kfihn & Greeff, 2002). As shown in Fig. 3, the slow gating mode was very prominent in recordings obtained from the WT rBIIA ~ subunit where f31 coexpression largely shifts the time course of inactivation to the fast gatingF.J.P. Kfihn and N.G. Greeff: $4D4 Movement and Sodium Channel Gating 27
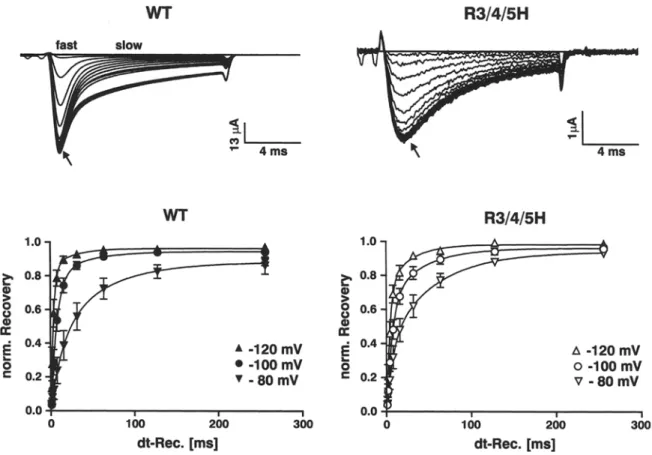
Fig. 4. Recovery from fast inactivation. (A) Original recordings of INa during recovery from fast inactivation of WT and R3/4/5H. The inactivating conditioning pulse to 0 mV from a holding po- tential of -100 mV had a duration of 100 ms. Interpulse intervals were increased logarithmically from 1 ms to 256 ms at a recovery potential of -100 mV and were followed by a 13-ms test pulse to 0 mV. Responses are superimposed for all recovery periods. Control traces obtained without inactivating prepulse are also included
(arrows). Note the predominance of the fast or the slow gating mode in WT at different points in the course of the current decay (as indicated). (B) Time course of recovery from fast inactivation of WT and R3/4/5H obtained from peak 1Na at three different re-
covery potentials, pulse protocol as described in A. For the cal- culation of the recovery kinetics, the current traces with prepulse were routinely normalized to the current traces without prepulse in order to compensate for the possible decrease due to rundown of the current amplitude in the course of the experiment. For the same reason, the first and the last recovery time interval of the pulse protocol were of identical duration (256 ms). The data (mean 4- SEM) were obtained from three (WT) and four (R3/4/5H) single-cell experiments and fitted to a double exponential curve of the form:
Y - AI 9 (1 - exp(-t/~m)) + A2. (1 - exp(-t/~R2)) + P with cor- responding recovery time constants given in the text.
m o d e ( p r e v i o u s l y d e s c r i b e d b y Bennett, M a k i t a & G e o r g e , 1993; P a t t o n et al., 1994). F u r t h e r m o r e , the kinetics o f b o t h g a t i n g m o d e s was slightly a c c e l e r a t e d w i t h c o r r e s p o n d i n g time c o n s t a n t s (rh(fast) / Zh(slow) in ms; m e a n i SEN, n = 3), as d e r i v e d f r o m similar re- c o r d i n g s as s h o w n in Fig. 3, for W T - 1 3 : ( 0 . 6 9 • 0.13 / 14.8 + 4.6) a n d for W T + 13:(0.69+0.11 / 9 . 1 + 1.7)
I n c o n t r a s t , the i n a c t i v a t i o n t i m e c o u r s e o f R 3 / 4 / 5H s h o w e d visually o n l y little c h a n g e d u e to the co- e x p r e s s i o n o f the 131 s u b u n i t , a n d the slow c o m p o n e n t was a n y w a y small a n d h a r d l y c h a n g e d in m a g n i t u d e d u e to the 131 s u b u n i t (Fig. 3). T h e c o r r e s p o n d i n g time
constants (rh(fast) / Zh(slow)
in ms; m e a n + SEN, n = 3, as d e r v i e d f r o m r e c o r d i n g s as s h o w n in Fig. 3 were for R 3 / 4 / 5 H - ~ : (3.8 + 0.6 / 18.8 + 1.6) a n d R 3 / 4 / 5 H + 13:(2.24 + 0.5 / 16.6 • 6.5). Thus, the i m p a c t o f the 131 s u b u n i t o n the time c o u r s e o f i n a c t i v a t i o n was significantly less p r o n o u n c e d in R 3 / 4 / 5 H c o m p a r e d to W T .RECOVERY FROM FAST INACTIVATION
I n the a b s e n c e o f
131
c o e x p r e s s i o n , the r e c o v e r y t i m e c o u r s e o f W T was clearly b i m o d a l , with a slow c h a n n e l p o p u l a t i o n s h o w i n g i n c o m p l e t e r e c o v e r y still after a n i n t e r p u l s e i n t e r v a l o f 256 m s at - 1 0 0 mV, w h e n the fast m o d e c h a n n e l s h a d c o m p l e t e l y recov- ered (Fig. 4A). I n c o n t r a s t , R 3 / 4 / 5 H d i d n o t s h o w such clearly s e p a r a b l e r e c o v e r y kinetics (Fig. 4A). T h i s result agrees with the m o r e m o n o p h a s i c inacti- v a t i o n kinetics o f R 3 / 4 / 5 H d u e to the r e l a t i v e l y s m a l l slowly i n a c t i v a t i n g p o p u l a t i o n , as d e s c r i b e d in the c o n t e x t o f Figs. 2 a n d 3. H o w e v e r , the d o u b l e - e x p o - n e n t i a l fits o f the r e c o v e r y time c o u r s e s r e c o r d e d at r e c o v e r y p o t e n t i a l s o f - 8 0 m V , - 1 0 0 m V a n d - 1 2 0 m V d e m o n s t r a t e t h a t the recoveries o f W T a n d R 3 / 4 / 5H were r a t h e r s i m i l a r (Fig. 4B), a n d trials to fit the r e c o v e r y o f the m u t a n t w i t h one e x p o n e n t i a l o n l y were m a r k e d l y worse. T h e c o r r e s p o n d i n g r e c o v e r y28 F.J.P. Kfihn and N.G. Greeff: $4D4 Movement and Sodium Channel Gating A 1.o ~,
o.s
0.6 0.4 O 0.2 0.0O
2;0
5;o
7;0
10~) 1250
dt-Rec. [ms] B 1.0- 0.8- o er04!
o e- 0.2-0,0
o
I;O
2;o
dt-Rec. [ms] zx -120 mV]
0-100 mV J R4Hv
-80 mV 9 -120 mV ] 9 -100 mV I WT 9 -80 mVeWT
]
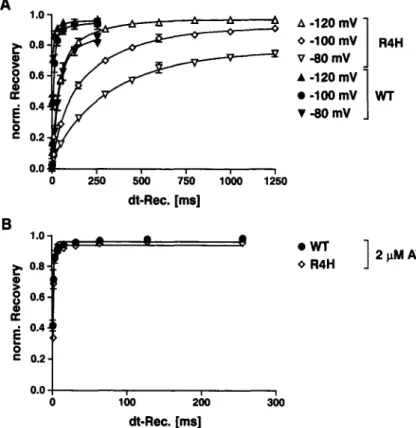
2 I~M ATX O R4HFig. 5. Recovery kinetics o f WT and mutant R1635H in the presence o f 2 I~M ATX-II. (A) Comparison of the recovery time course from fast inactivation of WT and R1635H (R4H) in the absence of ATX-I1 (see also Kfihn & Greeff, 1999). Same pulse protocol as described in the legend o f Fig. 4, but with regard to the different recovery kinetics of WT and R4H, separate interpulse-in- tervals were used. The data (mean • SEM of n = 2 cells) were obtained from single-cell experiments at the indicated recovery potentials and fitted as de- scribed in the legend of Fig. 4B with dominant recovery time constants in ms. WT-ct: 38 + 3.9 ( - 8 0 mV), 10.7 4- 4 . 6 ( - 1 0 0 m V ) , 4 . 6 4- 1.1 ( - 1 2 0 mV); RI635H<c 320 4- 39 ( - 8 0 mV), 199 • 26 ( 100 mV), 71 4- 11 ( - 1 2 0 mV). (B) Recovery of WT and R4H in the presence of 2 l,tM ATX-II obtained at a recovery potential of - 1 0 0 inV. Pulse protocol and fitting o f the data as described in the legend o f Fig. 4 with dominant recovery time constants in ms (mean + SEM o f n = 3 cells for WT and n = 5 cells for R4H) WT: 1.27 4- 0.64; R4H: 0.96 4- 0.24.
time
constants ("OR(fast) /
"OR(slow ) in ms) were for WT: 15.5 + 22.1 / 60.6 • 91.8 ( - 8 0 mV); 7.3 i 2.2 / 42.4 • 68.7 (-100 mV); 3.1 • 0.9 / 26.5 • 48.3 (-120 mV) and for R3/4/5H: 7.6 • 4.8 / 55.3 • 25.2 ( - 8 0 mV); 6.7 4- 2.2 / 41.0 4- 26.8 (-100 mV); 3.8 + 1.1 / 27.0 4- 21.4 (-120 mV). Thus, only at - 8 0 mV the recovery kinetics of WT was moderately delayed and less complete compared to R3/4/5H, which was most probably caused by the predominating slow-mode channels of WT. In comparison to our previously studied mutation R1635H (R4H), which drastically slows the recovery from fast inactivation (examples shown in Fig. 5A), the triple mutation R3/4/5H re- covers similarly to WT, as also observed for the double mutation R4/5H (Kfihn & Greeff, 1999).MODIFICATION BY SITE-3 TOXINS (ATX-II)
Site-3 toxins have been shown to selectively impair the movement of voltage sensor $4D4 (Sheets & Hanck, 1995). Moreover, Sheets et al. (1999) de- monstrated that the replacement of each of the three outermost arginines of voltage sensor $4D4 does not change the strong impact of these toxins on the in- activation time course as observed in WT, suggesting that the toxin effect surpasses the impact of the in- dividual mutations. This result supports the conclu- sions of Sheets et al. (2000) that the movement of voltage sensor $4D4 is actually inhibited by site-3 toxins, at least during the early phase of inactivation. In line with these data is our finding that in the pre- sence of 2 gM ATX-II, the very slow recovery kinetics
of R4H (Fig. 5A, see also Kfihn & Greeff, 1999) be- came as fast as the corresponding WT recovery (Fig. 5B). Thus, the impact of site-3 toxins seems to be independent from the previously changed kinetics of the mutated voltage sensor $4D4.
However, the comparison of the macroscopic current decay of/Na recorded from WT and R3/4/5H in the presence of 2 laM ATX-II (Fig. 6A) reveals that for R3/4/5H the toxin exerted only minor effects on the fast inactivation kinetics, whereas WT displays the characteristically slowed inactivation time course and a substantial plateau current as already described by others (e.g., Benzinger, Tonkovich & Hanck, 1999). The corresponding time constants given in Table 2 demonstrate that both gating modes of WT were profoundly slowed by ATX-II, whereas in R3/4/ 5H, both time constants were even slightly decreased in the presence of the toxin. Also included in Table 2 are the corresponding activation time constants (rm) that were determined as described in the legends of Figs. 2A and 8C. The data show that in particular for WT, the activation time constants were markedly reduced in the presence of ATX-II. In contrast, the ~m - values of R3/4/5H were only minimally affected by the toxin.
On the other hand, the comparison of Figs. 4A and 6B demonstrates that the recovery kinetics of R3/ 4/5H was accelerated by ATX-II but to a significantly smaller extent than observed in WT or R1635H (Fig. 5).
The reduced impact of ATX-II on the time course of fast inactivation and recovery of R3/4/5H
F.J.P. Kfihn and N.G. Greeff: $4D4 Movement and Sodium Channel Gating 29
Fig. 6. Effect of ATX-II on the inactivation kinetics of WT and R3/4/5H. (A) Typical sodium current families recorded in the presence of 2 gM ATX-II (for comparison see Fig. 1A), Currents were elicited by depolarizing pulses from -80 mV to + 80 mV from a holding potential of -100 mV; test pulse duration 80 ms. For the comparison of the effects on the time constants of activation and
inactivation induced by the toxin, see Table 2. (B) Recovery from fast inactivation of WT and R3/4/5H performed at a recovery potential of -100 mV, as described in the legend of Fig. 4 (for comparison, see also Fig. 4A). Note the different impact of the toxin on the fast inactivation kinetics as well as on the recovery kinetics of WT and R3/4/5H.
Table 2. Time constants of INa decay of WT and R3/4/5H obtained at 0 mV in the absence or in the presence of ATX WT (MBS) WT (ATX) R3/4/5H (MBS) R3/4/5H (ATX) T m 0.89 • 0.3 0.29 • 0.04 0.71 :t_ 0.2 0.51 • 0.1 Th(fast ) 0.86 • 0.4 8.55 • 1.6 3.70 -k 0.7 2.62 • 0.4 ~h(slow) 11.8 • 4.8 55.1 • 24.7 19.1 • 1.1 16.0 • 2.0
The data (mean in ms + SD of gt = 3 6 cells) were obtained as described in the legends of Figs. 2 and 8. ATX was used at a concentration of
2 p,M. Recordings were performed at 15~
c o u l d also reflect a d e c r e a s e d t o x i n affinity o f the m u t a n t a l t h o u g h the p u t a t i v e b i n d i n g site o f site-3 t o x i n s was l o c a l i z e d in the $ 3 - $ 4 e x t r a c e l l u l a r l o o p o f d o m a i n 4 ( R o g e r s et al., 1996). T h e r e f o r e , we e x a m - i n e d the t o x i n b i n d i n g in c o n c e n t r a t i o n e x p e r i m e n t s o f A T X - 1 I ; the i n a c t i v a t i o n kinetics was s t u d i e d in W T a n d in R 3 / 4 / 5 H at five different t o x i n c o n c e n - t r a t i o n s , a n d f u r t h e r in the a b s e n c e (Fig. 7A) o r in the p r e s e n c e (Fig. 7B) o f the 131 s u b u n i t . T h e results o b - t a i n e d a n d fitted to a s i n g l e - l i g a n d - t o - r e c e p t o r curve s h o w t h a t the affinity o f r a t b r a i n I I A s o d i u m c h a n - nels to A T X - I I was l a r g e l y unaffected b y the m u t a - t i o n R 3 / 4 / 5 H (Fig. 7C) w i t h a n E C s o o f b e t w e e n 0.2 to 0.4 g ~ A T X - I I . Likewise, the c o e x p r e s s i o n o f the [31 s u b u n i t d i d n o t significantly c h a n g e the effect o f A T X - I I o n s o d i u m c h a n n e l gating. T h u s , o n l y the
intrinsic effect o f the t o x i n (the size o f the p l a t e a u c u r r e n t relative to the p e a k ) was m a r k e d l y r e d u c e d in R 3 / 4 / 5 H , m o s t p r o b a b l y d u e to a c h a n g e d m o b i l i t y o f $4D4, which is in a g r e e m e n t with o u r o t h e r d a t a o f this m u t a n t ( s e e below).
ANALYSIS OF THE GATING KINETICS AT LOW TEMPERATU RE T h e s l o w e d i n a c t i v a t i o n t i m e c o u r s e as well as the r e d u c e d i m p a c t o f A T X - I I o n R 3 / 4 / 5 H suggest t h a t the m o b i l i t y o f v o l t a g e s e n s o r $ 4 D 4 is m a r k e d l y c h a n g e d in the m u t a n t . C o r r e s p o n d i n g l y , in case o f a n i n v o l v e m e n t o f $ 4 D 4 in c h a n n e l a c t i v a t i o n , R 3 / 4 / 5H s h o u l d also d i s p l a y different a c t i v a t i o n kinetics, if c o m p a r e d to W T . I n o r d e r to get s o m e i n f o r m a t i o n
30 FJ.P. Ktihn and N.G. Greeff: S4D4 Movement and Sodium Channel Gating
Fig. 7. Concentration-dependent effect of ATX-II on ratio of plateau to peak sodium currents. ( A - C ) Effect of ATX-II at bath concentrations of 0, 0.13, 0.25, 0.5, 1.0, and 2.0 gM on macroscopic sodium currents of WT and R3/4/5H obtained from single-cell experiments without (A) or with (B) [31 coexpression. Depolarizing pulses were to 0 mV from a holding potential of -100 mV, test pulse duration 80 ms. Traces were normalized to peak. (C) Ratios of plateau currents to peak currents obtained from A and B were plotted against toxin concentration. Note that the maximal effect of
ATX-II was reached at concentrations of about 2 ~.IM both in WT and R3/4/5H. The single-ligand binding was checked graphically in a Lineweaver-Burke-plot where the data points are close to a straight line (compare Denac et al., 2002). Accordingly, the data were fitted to a single-ligand-to-receptor binding curve: y = y ... . x / ( E C s o + x). ECsoATX in gM was for WT 0.4 (-13i) and 0.4 (+ 131), and for R3/4/5H 0.2 (-130 and 0.4 (+ 131); Ymax in % was for WT 45 (-131) and 36 (+131), and for R3/4/5H 22 (131) and
16(+ 13~).
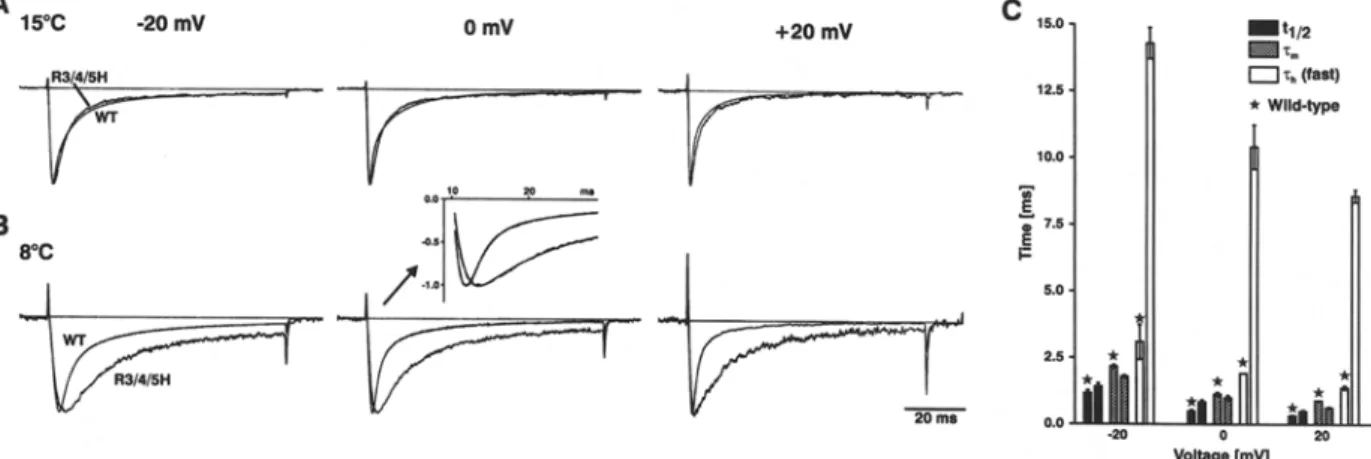
a b o u t the very fast activation kinetics o f W T and m u t a n t r B I I A s o d i u m channels, we recorded at a b a t h t e m p e r a t u r e o f 8~ where the channel kinetics is relatively slow c o m p a r e d to the capacitance transient, which reflects the speed o f voltage control. Figure 8 shows n o r m a l i z e d current traces o f W T and R 3 / 4 / 5 H s u p e r i m p o s e d a n d recorded at 15~ (A) a n d 8~ (B), respectively. T h e currents were elicited by step d e p o l a r i z a t i o n o f 80 ms d u r a t i o n to the indicated voltages r a n g i n g between - 2 0 m V and + 20 m V f r o m a h o l d i n g potential o f - 1 0 0 inV. First o f all, the c o m p a r i s o n d e m o n s t r a t e s that the inactivation ki- netics o f R 3 / 4 / 5 H was twice m o r e sensitive to low t e m p e r a t u r e t h a n observed in W T ( s e e c o r r e s p o n d i n g time c o n s t a n t s included in Table 1). These d a t a sug- gest that, especially in R 3 / 4 / 5 H , the mobility o f $ 4 D 4 was affected by low temperature.
Next, we determined b o t h the h a l f times to acti- vation (q/2) a n d the activation time constants (rm) o f W T and R 3 / 4 / 5 H f r o m single current traces recorded at 8~ u n d e r optimized clamping conditions in a 20 ms time w i n d o w a n d with a resolution o f 20 ~ts per d a t a point ( s e e sample traces in inset o f Figure 8B). The activation time c o n s t a n t (rm) and two inactiva- tion time c o n s t a n t s ('Ch(fast)/'Ch(slow), s e e legend) were derived. It is clearly seen that the activation kinetics o f R 3 / 4 / 5 H was n o t significantly different f r o m W T if one takes further into a c c o u n t that the two inde- p e n d e n t characteristic p a r a m e t e r s o f activation, rm and tl/2, yielded slightly o p p o s i n g but close to W T d a t a and, in addition, that the a b o u t 5-fold slowed inactivation time course o f the m u t a n t certainly af- fects the m e a s u r e m e n t o f the c o r r e s p o n d i n g activa- tion kinetics (Fig. 8C; s e e also Discussion).
F.J.P. Kfihn and N.G. Greeff: $4D4 Movement and Sodium Channel Gating 31
Fig. 8. Activation and inactivation kinetics of WT and R3/4/5H at
low temperature. (A-B) Comparison of typical single-current traces of WT and R3/4/5H recorded at 15~ (A) or 8~ (B). Currents were elicited by depolarizing pulses of 80 ms duration to the indi- cated voltages from a holding potential of -100 mV. Traces were normalized to peak. The data were fitted to a triple-exponential curve Y - (1 - exp(-t/~m)) 3. [ A 1 9 exp(--t/~hl)+A2, exp(--t/zh2)
+A3] with activation time constant (rm) and inactivation time constants (Zh(ram/ rh(slow) as also shown in Table 1). An example of data and fit is demonstrated in the inset for 0 mV at 8~ with -c m in ms for WT/R3/4/5H equal to 1.01/1.07 and %,(r~so in ms equal to
1.99/9.55. (C) Effects of mutation R3/4/5H on time constants of half activation (tl/2) , activation (Tin) and fast inactivation (rh(ram) obtained at 8~ (compare inset in B), with minimal Rs-distortion and at constant clamping speed. The corresponding WT values are indicated by asterisks. The values for q/2 in ms (mean • SD ob- tainedat 20mV/0mV/ +20mVforn = 3 cells) were WT:1.16 :t- 0.2 / 0.49 + 0.1 / 0.31 • 0.1 and R3/4/5H: 1.41 • 0.2/0.83 • 0.1 / 0.48 • 0.1) and for zm in ms (mean + SD obtained at 20 mV/0 mV/+20 mV for n = 2 cells) were WT: 2.16 • 0.1/1.13 • 0.1/0.89 • 0.01 and R3/4/5H: 1.78 • 0.1/0.99 + 0.1/0.63 • 0.05).
Discussion
In continuation of our former a p p r o a c h to inhibit the m o v e m e n t of $4D4 by the systematic replacement of central arginines to histidines in this voltage sensor (Kfihn & Greeff, 1999), we introduced the triple mutation R1632H + R1635H + R1638H ( = R3/4/SH) and analyzed the effects on the gating kinetics of rat brain I I A sodium channel. First of all, the m u t a n t showed a considerably reduced expression c o m p a r e d to WT. This was reflected both on the level of the ionic current as well as of the gating current, the latter being too small to be separated from baseline noise and a s y m m e t r y artifacts, respectively. The presence of inadvertent mutations in other regions of the channel could be excluded because several clones were tested yielding the same results. Likewise, a specific reduction of the available gating charge of R3/4/5H due to the inhibition of the mobility of $4D4, as observed by Sheets et al. (1995; 1999; 2000) in the presence of site-3 toxins, is rather unlikely be- cause the well-resolved sodium current of R3/4/5H displayed quite normal activation and inactivation properties. Instead, it is conceivable that the triple m u t a t i o n in some way interferes with the proper folding of the protein because the substituted argi- nines possibly participate in a network of charged residues that stabilizes the channel structure. Similar results were described for some mutants of $4 seg- ments of voltage-dependent sodium channels (Stfih- mer et al., 1989) or potassium channels (Papazian et al., 1995), but these mutations caused the loss of
functional expression altogether. In other studies performed in our own lab we noticed that some sodium channel m u t a n t s also showed decreased ex- pression levels, if c o m p a r e d to W T (e.g., Paramyo-
tonia congenita equivalent m u t a t i o n R1626C or
IFMI488-1490QQQ; unpublished data). There, we
have some experimental data that these channels, which show strongly reduced or completely abolished fast inactivation properties, were leaky for sodium during expression in Xenopus oocytes and, therefore, indirectly reduce the expression level (Greeff & Kfihn, 2OOO).
In a previous study we have demonstrated that the replacements of arginines by histidines in the central part of voltage sensor $4D4 (mutation R4H, R4/SH and R5H) predominantly induce steric effects rather than changing the voltage dependence of the channel (Kfihn & Greeff, 1999). Accordingly, the slight decrease of the voltage dependence for entry and exit from the inactivated state observed in R3/4/ 5H suggests that the substitution of the third arginine is responsible for this additional effect. This conclu- sion is supported by the data of other groups (Chen et al., 1996; Yang et al., 1996; Sheets et al., 1999). As to the charge, Histidine has, depending on the local protein environment, a p K of 5.6-7.0 and therefore, the charge of this amino acid could change under varying p H conditions (Sancho et al., 1992). Despite the fact, that there was no control of the intracellular solution using the two-electrode voltage-clamp tech- nique, we varied the extracellular p H between 5 and 9 without seeing any effects (data not shown).
32 F.J.P. Kiihn and N.G. Greeff: $4D4 Movement and Sodium Channel Gating As established by the authors (Ktihn & Greeff,
1999), the mutation R1635H (R4H) in the central part of $4D4 dramatically increased (~20-fold) the recovery time constants of both fast inactivation and voltage-sensor immobilization (visible in the gating current) and therefore, exerted one of the strongest effects on the kinetics of $4D4 compared to previ- ously described mutations of this voltage sensor (Chahine et al., 1994; Chen et al., 1996; Kontis & Goldin, 1997). In contrast to recovery, R4H showed only a small deceleration of the kinetics of current decay compared to WT. On the other hand, the double mutation R1635H + R1638H (R4/5H) ana- lyzed in the same study, moderately (~2-fold) slowed the recovery kinetics but increased the time constants of fast inactivation to a larger extent than observed in R4H. Similarly, the mutation R3/4/5H significantly slowed the time course of fast inactivation, whereas the recovery kinetics was almost identical to WT. These results suggest that a single histidine-for-argi- nine substitution in the central part of $4D4 is less tolerable for the recovery from fast inactivation, and vice versa less critical for the development of fast inactivation, than the corresponding double or triple substitution. In line with this observation are the data of Groome et al. (2002) who found that different al- terations of the structure of the arginine moiety at either R4 or R5 impedes or facilitates the movement of voltage sensor $4D4 during deactivation.
The bimodal gating behavior characteristic for
WT rBIIA sodium channels, if expressed in
Xenopus
oocytes without the accessory [31 subunit, is less pronounced in R3/4/5H, which is reflected by the monophasic recovery time course of the mutant. Accordingly, the pronounced shift to the fast gating mode induced by [31 coexpression is not observed in R3/4/5H. This result suggests a putative role of $4D4 for the determination of the gating mode during ex- pression of rat brain sodium channels in
Xenopus
oocytes. Accordingly, recent findings of Mantegazza et al. (2001) had demonstrated the principal role of the C-terminal domain of brain and cardiac sodium channels for the kinetics and voltage dependence of inactivation.
Apart from single or multiple arginine substitu- tions also site-3 toxins selectively change the mobility of voltage sensor $4D4 (Sheets et al., 1995; 1999; 2000; Benzinger et al., 1999). Furthermore, Sheets et al. (1999) demonstrated that the replacement of the three outermost arginines of $4D4 did not change the effect of site-3 toxin on the fast inactivation kinetics although each of these mutations alone markedly slowed the inactivation time course of the channel. In line with this observation is our finding that the 20- fold slowed recovery time courses of mutant R4H became as fast as the corresponding WT recovery in the presence of ATX-II. Hence, the impact induced by the toxin was independent of the corresponding
effect of the mutations. These data support the hy- pothesis of Sheets et al. (2000) that site-3 toxins se- lectively inhibit the movement of voltage sensor $4D4. However, a remaining question is the extent of this inhibition. If site-3 toxins actually arrest the movement of $4D4, this should abolish fast inacti- vation because a series of experimental data have clearly demonstrated the principal role of this voltage sensor in sodium channel fast inactivation (Chahine et al., 1994; Yang & Horn, 1995; Chen et al., 1996; Yang et al., 1996; Kontis & Goldin, 1997; Ktihn & Greeff, 1999). Moreover, if the movement of $4D4 is in some way coupled to the activation process, which is suggested by the data of Horn et al. (2000), then channel activation should be prevented or at least severely impaired by the toxin, which is obviously not the case.
In fact, we found that the impact of ATX-II on the fast inactivation kinetics of R3/4/5H was con- siderably reduced compared to WT. To our knowl- edge, this is the first mutation in a region other than the putative binding site that reduces the efficacy of ATX-II without changing significantly the affinity of the channel for the toxin. According to our hypoth- esis, the effect of ATX-II can be explained by elec- trostatic interactions of the positively charged toxin with the outermost arginines of $4D4. As a result, the outward movement of $4D4 is delayed leading to slowed inactivation kinetics and vice versa the return of the voltage sensor is favored, implicating an ac- celerated recovery from fast inactivation. Therefore, the data of the current study demonstrate that the mobility of $4D4 was considerably changed by the mutation R3/4/5H, which was reflected by the slowed inactivation kinetics and the reduced impact of ATX- II.
While this study has clearly confirmed the well- established hypothesis that the voltage sensor $4D4 is involved in fast inactivation, it shall now be discussed what bearing our data have for a role of this segment for activation. Recalling the finding that R3 moves through the "gating pore", as shown by cysteine mutagenesis and MTSET accessibility experiments (Yang et al., 1996), it would appear that a replace- ment by the bulky histidine would hinder the outward movement of $4D4 through the "gating pore" at an earlier stage than R4H or R5H. According to our previous study, R4H and R5H have only an impact on fast inactivation but not on activation (Ktihn & Greeff, 1999). We then concluded that at least one voltage-dependent conformational step controls the inactivation gate. However, this did not exclude a first step of outward movement of S4D4 that might control an activation gate or might delay inactiva- tion. For the extended mutation
R3/4/5H
it appears plausible that it would block the outward movement at an earlier stage and would more likely indicate a putative involvement of $4D4 in activation.F.J.P. Kfihn and N.G. Greeff: $4D4 Movement and Sodium Channel Gating 33
To analyze a putative impact on activation, the experiments obtained at the lower temperature of 8~ were used where the fast sodium channel currents are slow in comparison to the speed of the clamp. Furthermore, we used an optimized two-electrode voltage clamp that allowed us to establish fast voltage steps at the whole oocyte (Greeff & Polder, 1998). This clamp design had already proved its worth for the recording of the very fast sodium channel gating currents (Kiihn & Greeff, 1999; Greeff & Ktihn, 2000). Special care was taken to adjust the Rs com- pensation such that the speed-limiting capacitance transient was fast and of equal speed in WT and mutant channel-expressing oocytes. Rs errors were further reduced by choosing oocytes with small so- dium currents, especially WT. For the analysis, cur- rents were recorded in 100 and 20 ms time windows to allow fitting of both, the inactivation and the ac- tivation phase (Fig. 8B and 8C). While rl,(t~tl is in- creased by a factor of 4 to 5, activation is hardly changed in R3/4/5H compared to WT. The data also show that the fast inactivation kinetics of R3/4/5H at 8 ~ was slowed to a clearly larger extent than ob- served in WT, which might suggest that the move- ment of the mutated $4D4 segment was reduced by sterical obstruction. As a regular feature it is ob- served that the time to half activation
tj/2
is about 20% larger in the mutant, while the fitted tm is about 20% smaller than in WT. The fitted equation is of the Hodgkin-Huxley type with parallel activation and inactivation, i.e., a product of a rising and a declining exponential. For such systems it is easily shown that the apparentt~/2
becomes larger for a constant tm but increasing th (f~t). It is further easily seen that the open-probability po at peak ionic current gets larger for slower inactivation and constant gn; slow inacti- vation allows more channels to be open before they inactivate. In our data, the WT po is between 0.3 and 0.5 while it is about 0.8 for the mutant, as obtained by the fitting. This has two consequences for the inter- pretation of our data: (i) TheI/V
curves (Fig. 1B), which usually show peak ionic currents and not the fraction of activated channels, should be corrected for inactivation. Then, the small difference, being anyway partly due to some Rs error for WT would become even less significant; however, a correction for inactivation can only properly be done if the coupling mode of activation to inactivation is known, which is presently not the case. (ii) The other conse- quence is that the time course of the sodium current traces should not be compared with peaks normal- ized, but the peak for the mutant should be larger according to the larger po and then the initial time course becomes as fast as in WT or even faster, de- pending on the model one uses to correct for inacti- vation.In summary, we conclude that activation is not slowed in R3/4/5H and, therefore, the data of this
study still give no support for an involvement of $4D4 as voltage sensor for an activation gate. How- ever, this question is still not fully settled and our data have to be seen as experimental evidence to be compared with other data that support a first acti- vation step (Horn et al., 2000) or do not support it (Sheets et al., 2000); also data about $4D4 obtained by a new experimental approach conform with both (Chanda & Bezanilla, 2002). Finally, it has to be mentioned that our study has dealt with the clear-cut effects on fast inactivation. Other studies suggest an additional role of the $4D4 mid-region arginines (Mitrovic, George and Horn, 2000) or the P-loop region of domain 4 (Hilber et al., 2002) for the so- called slow inactivation. Such slow inactivation was not addressed in the study reported here, and there- fore, we exclusively used pulse protocols where slow inactivation effects were negligible.
We thank Dr. Alan L. Goldin (Irvine, CA) for providing the cDNA of the wild-type rat brain I1A sodium channel (pVA2580) and the high expression vector (pBSTA). We are grateful to Dr. William A. Catterall (Seattle, WA) for providing the cDNA of the [31-subunit, to Rainer Polder (npi electronic GmbH, Tamm, Germany) for his support in developing the modified two-electrode voltage clamp and to Christian Gasser (ZOrich, Switzerland) for his graphical assistance. The work was supported by the Swiss National Science Foundation (grant No. 31-37987.931 and the Hartmann-Mtiller- Stiftung.
References
Balser, J.R., Nuss, H.B., Chiamvimonvat, N., P~rez-Garcia, M.T., Marbtin, E., Tomaselli, G.F. 1996. External pore residue mediates slow inactivation in lal rat skeletal muscle sodium channels. J. Physiol. 494:431M42
Bennett, P.B., Makita, N., George, A.L., Jr. 1993. A molecular basis for gating mode transitions in human skeletal muscle sodium channels. FEBS Lett. 326:21 24
Benzinger, G.R., Tonkovich, G.S., Hanck, D.A. 1999. Augmen- tation of recovery from inactivation by site-3 Na channel tox- ins. J. Gen. Physiol. 113:333 346
Catterall, W.A. 1986. Molecular properties of voltage-sensitive sodium channels. Ann. Rev. Biochem. 55:953 985
Cha, A., Ruben, P.C., George, A.L., Jr., Fujimoto, E., Bezanilla, F. 1999. Voltage sensors in domains lIl and IV, but not I and II are immobilized by Na ~ channel fast inactivation. Neuron.
22:73-87
Chahine, M., George, A.L., Jr., Zhou, M., Ji, S., Sun, W., Barchi, R.L., Horn, R. 1994. Sodium channel mutations in paramyo- tonia congenita uncouple inactivation from activation. Neuron.
12:281 294
Chanda, B., Bezanilla, F. 2002. Tracking voltage-dependent con- formational changes in skeletal muscle sodium channel during activation. J. Gen. Physiol. 120:629-645
Chen, L.-Q., Santarelli, V., Horn, R., Kallen, R.G. 1996. A unique role for the $4 segment of domain 4 in the inactivation of so- dium channels. J. Gen. Physiol, 108:549- 556
Denac, H., Mevissen, M., Ktihn, F.J.P., KO.hn, C., Guionaud, C.T., Scholtysik, G., Greeff, N.G. 2002. Molecular cloning and functional characterization of a unique mammalian cardiac
34 F.J.P. KiJhn and N.G. Greeff: $4D4 Movement and Sodium Channel Gating
Na(v) channel isoform with low sensitivity to the synthetic in- activation inhibitor (-)-(S)-6-amino-alpha-[(4-diphenylmethyl-1- piperazinyl)-methyl]-9H-purine-9-ethanol (SDZ 211-939).
J. Pharm. Exptl. Ther. 303:89 98
Greeff, N.G., Kfihn, F.J.P. 2000. Variable ratio of permeability to gating charge of rBIIA sodium channels and sodium influx in
Xenopus oocytes. Biophys. J. 79:2434-2453
Greeff, N.G., Polder, R. 1998. Optimization of a two-electrode voltage clamp for recording of sodium ionic and gating currents from Xenopus oocytes. Biophys. J. 74:A402
Groome, J.+ Fujimoto, E., Walter, L., Ruben, P.C. 2002. Outer and central charged residues in DIVS4 of skeletal muscle sodium channel have differing roles in deactivation. Biophys. J. 82:1293 1307
Hilber, K., Sandtner+ W., Kudlacek, O., Schreiner, B., Glaaser, I., Schuetz, W., Fozzard, H.A., Dudley, S.C., Todt, H. 2002. In- teraction between fast and ultra-slow inactivation in the volt- age-gated sodium channel: Does the inactivation gate stabilize the channel structure? J. Biol. Chem. 277:37105 37115 Horn, R., Shinghua, D., Gruber, H.J. 2000. Immobilizing the
moving parts of voltage-gated ion channels. J. Gen. Physiol. 116:461475
Isom, L.L., De Jongh, K.S., Catterall, W.A. 1994. Auxiliary su- bunits of voltage-gated ion channels. Neuron. 12:1183 1194 Isom+ L.L., De Jongh, K.S., Patton, D.E., Reber, B.F.X., Offord,
J., Charbonneau, H., Walsh, K., Goldin, A.L., Catterall, W.A. 1992. Functional coexpression of the L3-1 subunit of the rat brain sodium channel. Science. 256:839 842
Ji, S., George, A.L., Jr., Horn, R., Barchi, R.L. 1994. Voltage- dependent regulation of modal gating in the rat SkMI sodium channel expressed in Xenopus oocytes. J. Gen. Physiol. 104:625 643
Kontis, K.J., Goldin, A. 1997. Sodium channel inactivation is al- tered by substitution of voltage sensor positive charges. J. Gen.
Physiol. 110:403 ~, 13
Kontis, K.J., Rounaghi, A., Goldin, A. 1997. Sodium channel ac- tivation gating is affected by substitutions of voltage sensor positive charges in all four domains. J. Gen. Physiol. 110:391 401
Kfihn, F.J.P., Greeff, N.G. 1999. Movement of voltage sensor $4 in domain 4 is tightly coupled to sodium channel fast inactivation and gating charge immobilization. J. Gen. Physiol. 114:167 183 Kfihn, F.J.P., Greeff, N.G. 2002. Mutation D384N alters recovery of the immobilized gating charge in rat brain IIA sodium channels. J. Membrane Biol. 185:145 155
Mantegazza, M., Yu, F.H., Catterall, W.A., Scheuer, T. 200l. Role of the C-terminal domain in inactivation of brain and cardiac sodium channels. Proc. Natl. Acad. Sci. USA 98:15348-15353 Mitrovic, N., George, A.L., Jr., Horn, R. 2000. Role of domain 4
in sodium channel slow inactivation. J. Gen. Physiol. 115:707 718
Moorman, J.R., Kirsch, G.E., VanDongen, A.M.J., Joho, R.H., Brown, A.M. 1990. Fast and slow gating of sodium channels encoded by a single mRNA. 1990. Neuron. 4:243 252
Moran, O., Melani, R., Nizzari, M., Conti, F. 1998. Fast- and slow- gating modes of the sodium channel are altered by a para- myotonia congenita-linked mutation. J. Bioenerg. Biomembr. 30:579-588
Noda, M., Ikeda, T., Suzuki, H., Takeshima, H., Takahashi, H., Kuno, M., Numa, S. 1986. Expression of functional sodium channels from cloned cDNA. Nature. 322:826 828
Papazian, D.M., Shao, X.M., Seoh, S-A., Mock, A.F., Huang, Y., Wainstock, D.H. 1995. Electrostatic interactions of $4 voltage sensor in Shaker K + channel. Neuron. 14:1293-1301
Patton, D.E., Isom, L.L., Catterall, W.A., Goldin, A.L. 1994. The adult rat brain [3L subunit modifies activation and inactivation gating of multiple sodium channel ~ subunits. J. Biol. Chem. 269:17649-17655
Rogers, J.C., Qu, Y., Tanada, T.N., Scheuer, T., Catterall, W.A. 1996. Molecular determinants of high affinity binding of a- Scorpion Toxin and Sea Anemone Toxin in the $3-$4 extra- cellular loop in domain IV of the Na + channel c~ subunit.
J. Biol. Chem. 271:15950 15962
Sancho, J., Serrano, L., Fersht, A.R. 1992. Histidine residues at the N- and C-termini of alpha-helices: perturbed pKas and protein stability. Biochemistry. 31:2253-2258
Schreibmayer, W., Lester, H.A., Dascal, N. 1994. Voltage clamp- ing of Xenopus laevis oocytes utilizing agarose-cushion elec- trodes. Pfluegers Arch. 426:453~458
Sheets, M.F., Hanck, D.A. 1995. Voltage-dependent open-state inactivation of cardiac sodium channels: gating current studies with Anthopleurin-A toxin. J. Gen. Physiol. 106:617 640 Sheets, M.F., Kyle, J.W., Kallen, R.G., Hanck, D.A. 1999. The Na
channel voltage sensor associated with inactivation is localized to the external charged residues of domain IV, $4. Biophys. J. 77:747-757
Sheets, M.F., Kyle, J.W., Hanck, D.A. 2000. The role of the pu- tative inactivation lid in sodium channel gating current immo- bilization. J. Gen. Physiol. 115:609 619
Shih, T.M., Smith, R.D., Toro, L., Goldin, A.L. 1998. High-level expression and detection of ion channels in Xenopus oocytes.
Meth. Enzymol. 293:529-556
Sigworth, F.J. 1993. Voltage gating of ion channels. Q. Rev. Bio-
phys. 27:1J,0
Stiihmer, W., Conti, F., Suzuki, H., Wang, X., Noda, M., Yahagi, N., Kubo, H., Numa, S. 1989. Structural parts involved in ac- tivation and inactivation of the sodium channel. Nature. 339:597 603
Yang, N., Horn, R. 1995. Evidence for voltage dependent $4 movements in sodium channels. Neuron. 15:213-218
Yang, N., George, A.L., Horn, R. 1996. Molecular basis of charge movement in voltage-gated sodium channels. Neuron. 16:113 122
Yellen, G. 1998. The moving parts of voltage-gated ion channels.
Q. Rev. Biophys. 31:239-295
Zhou, J., Potts, J.F., Trimmer, J.S., Agnew, W.S., Sigworth, F.J. 1991. Multiple gating modes and the effect of modulating fac- tors on the ~tl sodium channel. Neuron. 7:775 785