Subtelomeric Transcription and its Regulation
Texte intégral
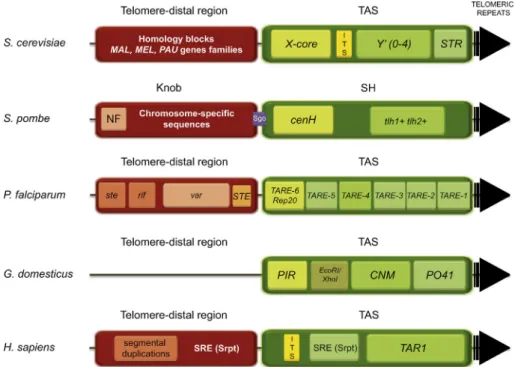
Figure
Documents relatifs
Before considering the use of absolute qPCR data in comparative studies, it remains crucial to validate absolute qPCR-based TL estimates by comparing among study, indi- viduals
Selon l’exercice d’une fonction d’encadrement 50 49 Managers Non managers 50 42 58 51 54 52 Moins de 20 salariés 20 à 249 salariés 250 à 499 salariés 500 à 1999 salariés 2000
Banking Marketing Mix.. :تايوححلما سرهف تدفصلا يىخدلما - ءادهالإ - سىشلا تملو - صخللما - ثايىخدلما ضسهف - ٌايشالأ ضسهف - ٌوادجلا ضسهف د ـــــــــ أ
152 Les souches d’ETEC peuvent produire des entérotoxines thermostables (sta) et / ou thermolabiles (lt) (Sjoling et al. Dans cette étude ces gènes codant pour les ETEC ont
The values of the ergonomic indicators are compared for the three cases (without assistance, with weight compensation, and with the co-worker robot).. C.1.1
Le professeur Marc Nihoul, directeur du Centre de protection juridique du citoyen de l’Université de Namur, lors de son intervention au colloque « L’article 159 de la Constitution
On peut alors facilement tracer l'axe (d).. Calcule la mesure de
