Mosquito Gonotrophic Cycle and Multiple Feeding Potential: Contrasts Between Anopheles and Aedes (Diptera: Culicidae)
5
0
0
Texte intégral
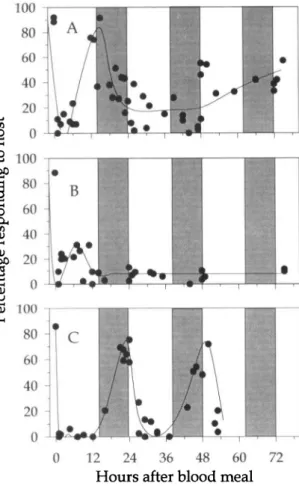
Figure

Documents relatifs