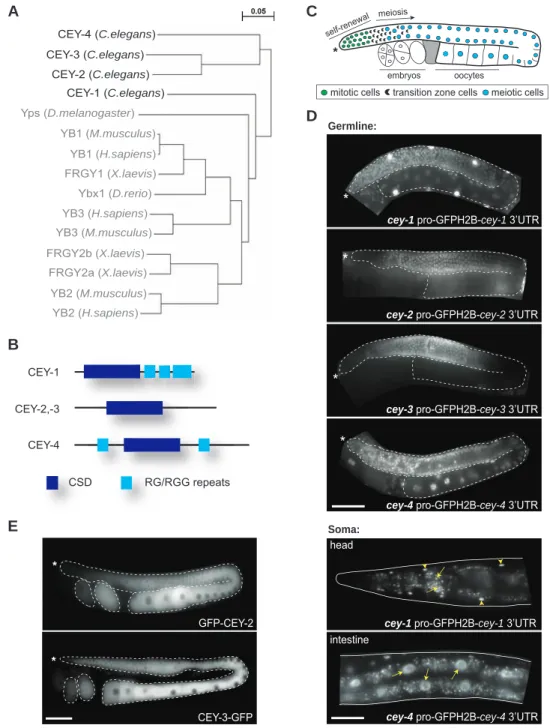
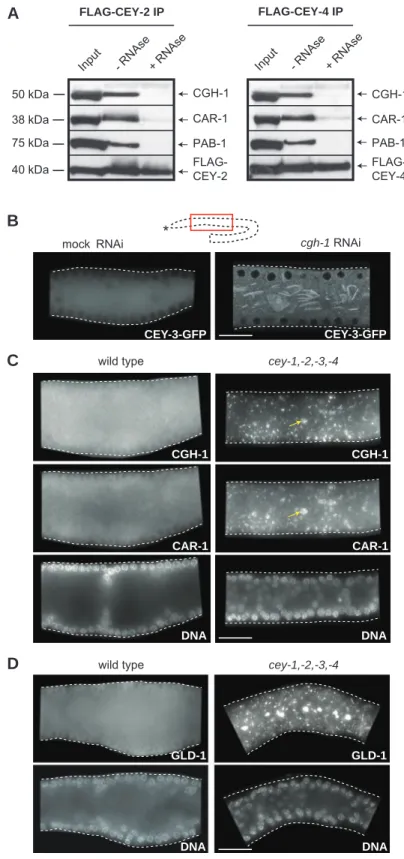
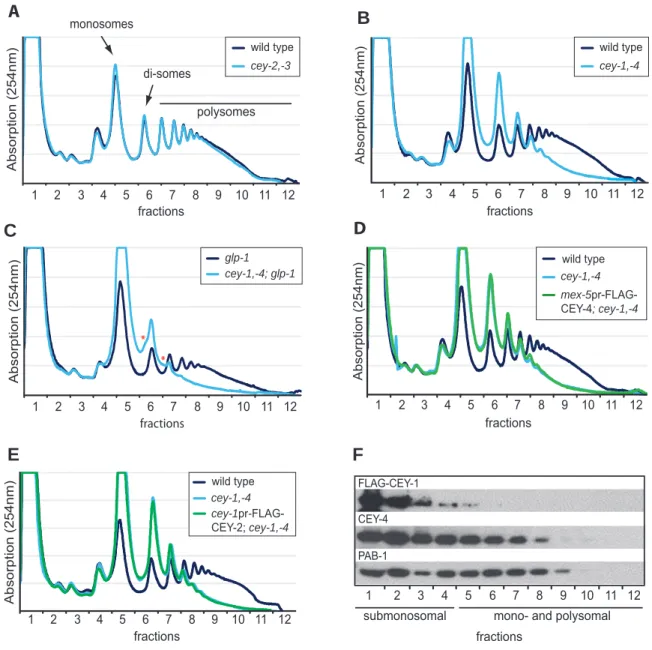
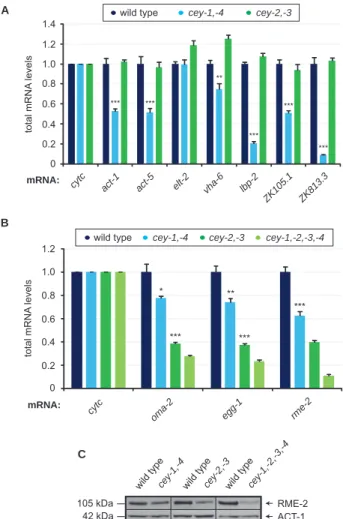
Functional characterization of C. elegans Y-box-binding proteins reveals tissue-specific functions and a critical role in the formation of polysomes
17
0
0
Texte intégral
Figure


+5


Documents relatifs