Influence de 6 semaines d'entraînement par intervalles à haute intensité sur l'autorégulation cérébrale dynamique d'hommes entraînés en endurance
100
0
0
Texte intégral
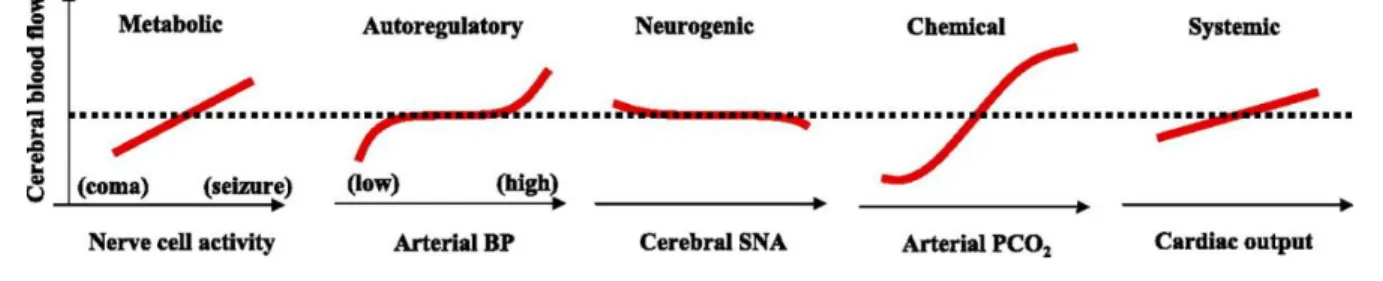
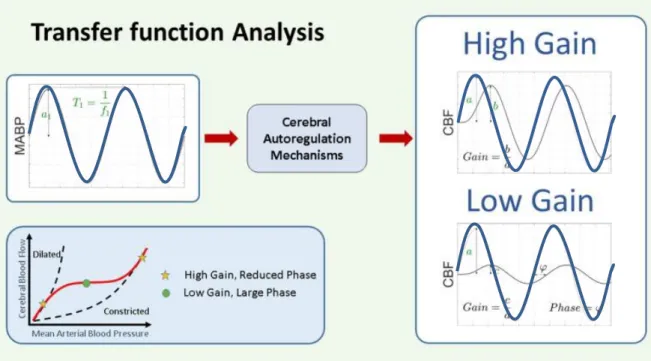
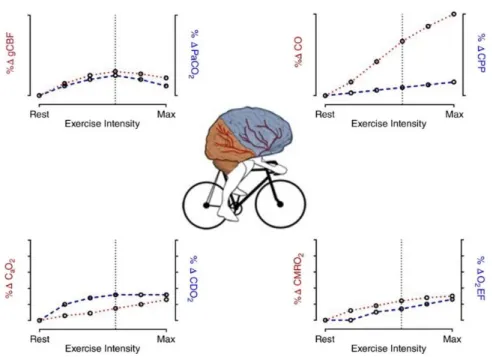
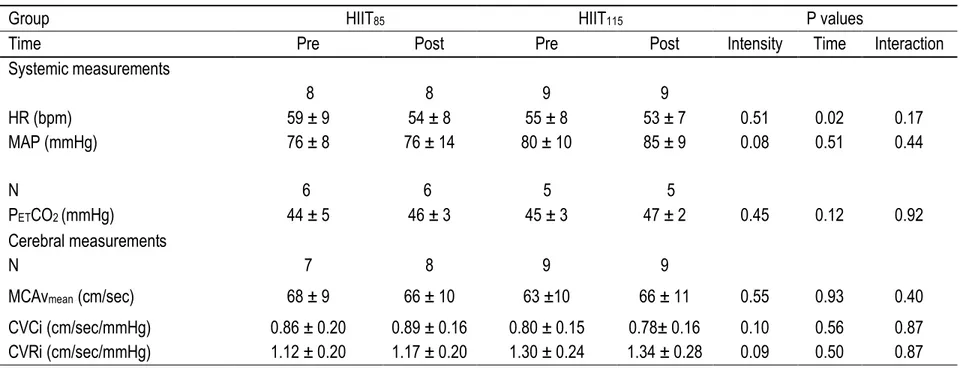
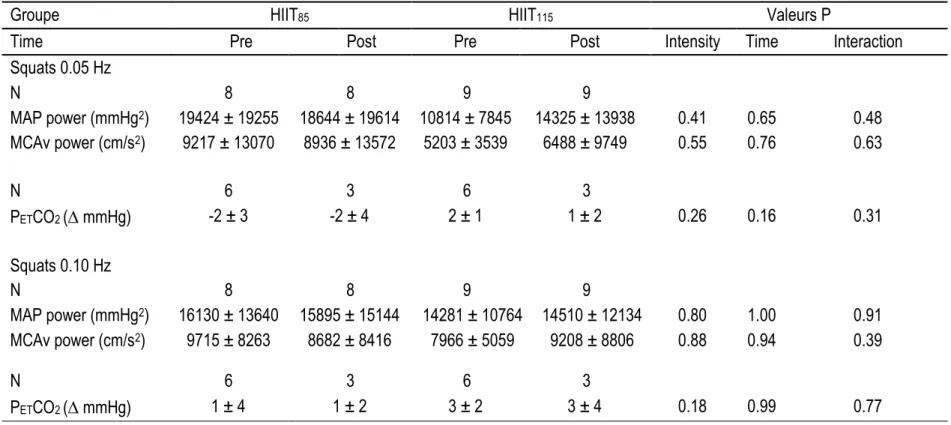
Figure



+6

Documents relatifs
