U. F. R. STAPS
Centre de Recherches sur la Cognition et l’Apprentissage CNRS – UMR 6234
THÈSE
Pour l’obtention du grade de
DOCTEUR DE L’UNIVERSITÉ DE POITIERS
SPECIALITE : SCIENCES ET TECHNIQUES DES ACTIVITES PHYSIQUES ET SPORTIVES
Présentée par
Raghad FADEL
ÉTUDE CHRONOMÉTRIQUE ET ÉLECTROPHYSIOLOGIQUE DU
CONTRÔLE DES ERREURS DE PROGRAMMATION DU
MOUVEMENT CHEZ L’HOMME
Sous la direction des Professeurs
Camille-Aimé POSSAMAÏ & Michel AUDIFFREN
Membres du jury :
M. Michel AUDIFFREN, Professeur, Université de Poitiers (Co-directeur)
M. Thierry HASBROUCQ, Directeur de Recherche au CNRS, Marseille (Rapporteur)
M. François MAQUESTIAUX, Maître de Conférences, Université Paris-Sud 11 (Examinateur) M. Vincent NOUGIER, Professeur, Université de Joseph Fourier, Grenoble (Rapporteur) M. Camille-Aimé POSSAMAÏ, Professeur, Université de Poitiers (Directeur)
À ton âme… Ayman
Mes remerciements s’adressent, tout d’abord, à mes directeurs de thèse, le Professeur Camille-Aimé POSSAMA au Professeur Michel AUDIFFREN. Depuis mon arrivée en France et tout au long de ce travail doctoral, ils ont su m’apporter un soutien constant et des conseils précieux et avisés à la hauteur de leur expérience, de leurs compétences et de leurs réelles qualités humaines. Leur conception de la direction de recherche, alliant rigueur et bienveillance, m’a aidée à donner le meilleur de moi-même.
Mes remerciements s’adressent également aux Professeurs Thierry HASBROUCQ et Vincent NOUGIER, pour l’honneur qu’ils me font en acceptant d’être les rapporteurs pour ce travail de thèse.
Je tiens sincèrement à remercier Monsieur François. MAQUESTIAUX, pour avoir accepté de juger ce travail et pour sa participation au jury.
Je remercie particulièrement le Professeur Yannick BLANDIN, pour ses conseils, son soutien scientifique et ses qualités humaines qui m’ont aidé à mener à bien ce travail.
Merci à F. CHELLI pour avoir réalisé ma première expérience. Merci à tous les participants qui se sont prêtés au jeu des expérimentations
Mes remerciements chaleureux vont à l’ensemble du personnel de l’équipe ATCO, du CeRCA et de la MsHS pour l’aide et l’amitié qu’ils m’ont apportées ; un merci particulier à mes collègues de bureau pour tous ces moments passés dans la joie et la bonne humeur.
Un grand merci à mes amis au sens large. Je les remercie pour les bons moments passés avec eux, qui m’ont aidée à surmonter les moments de découragements. Merci à chacun pour sa présence.
Finalement j’adresse un grand merci à toute ma famille qui a toujours été présente lorsque j’en ai eu besoin, en particulier à mes parents, à ma sœur et à mon frère, sans qui je ne serais pas où j’en suis aujourd’hui.
INTRODUCTION GÉNÉRALE
...
7Chapitre.I. Partie théorique... 9
I.1. Système du traitement de l’information... 10
I.2. Paradigme de la chronométrie mentale ... 13
I.2.1 Procédure du temps de réaction ... 13
I.2.2 Différentes tâches de temps de réaction... 14
I.3. Programmation motrice ... 16
I.4. Préparation motrice ... 20
I.4.1 Définition ... 21
I.4.2 Paradigme de pré-signalisation (precuing) ... 22
I.4.3 Paradigme d’amorçage (priming)... 23
I.5. Inhibition... 28
I.6. Différentes méthodes d’analyse utilisées dans ce travail ... 33
I.6.1 Analyse des distributions de temps de réaction ... 33
I.6.2 Technique électromyographique ... 37
I.6.2.1 Fractionnement de temps de réaction... 38
I.6.2.2 Erreurs infraliminaires (ébauches d’erreur) ... 40
I.6.3 Fonction d’échange vitesse / précision... 41
I.6.4 Analyse des erreurs et des ébauches d’erreur... 42
Chapitre.II. Partie expérimentale ... 44
II.1. PREMIÈRE ÉTUDE... 45
II.1.1 Introduction ... 45
II.1.2 Objectif... 49
II.1.3 Méthode... 50
II.1.3.6 Statistiques ... 58
II.1.4 Résultats ... 58
II.1.4.1 Temps de réaction correct ... 58
II.1.4.2 Taux d’erreurs ... 59
II.1.4.3 Temps de réaction erroné dans le cas où la réponse exécutée est celle amorcée ... 60
II.1.4.4 Distributions de temps de réaction correct... 61
II.1.4.5 Fonctions de précision conditionnelle... 63
II.1.4.6 Temps de réaction erroné dans le cas où la réponse exécutée est la réponse préparée... 65
II.1.5 Discussion ... 68
II.2. DEUXIÈME ÉTUDE ... 72
II.2.1 Introduction ... 72
II.2.2 Méthode... 73
II.2.2.1 Sujets ... 73
II.2.2.2 Dispositif expérimental ... 73
II.2.2.3 Tâche et procédure ... 76
II.2.2.4 Protocole... 77
II.2.2.5 Traitement des données... 78
II.2.2.6 Statistiques ... 78
II.2.3 Résultats ... 78
II.2.3.1 Temps de réaction et taux d’erreurs pour la session d’entraînement ... 78
II.2.3.2 Temps de réaction correct et taux d’erreurs de la session expérimentale 79 II.2.3.3 Temps de réaction erroné dans le cas où la réponse exécutée correspondant à la réponse amorcée... 81
II.2.4 Discussion ... 83
II.3. TROISIÉME ÉTUDE... 85
II.3.1 Introduction ... 85
II.3.2 Méthode... 90
II.3.2.1 Sujets ... 90
II.3.2.2 Dispositif expérimental ... 90
II.3.2.3 Protocole... 90
II.3.2.4 Enregistrement électromyographique EMG... 91
II.3.3 Résultats ... 96
II.3.3.1 Temps de réaction et taux d’erreurs pour la session d’entraînement ... 96
II.3.3.2 Temps de réaction correct et taux d’erreurs de la session expérimentale .... ... 97
II.3.3.3 Temps de réaction dans le cas des essais purs-corrects ... 99
II.3.3.4 Temps pré-moteur dans le cas des essais purs-corrects ... 100
II.3.3.5 Temps Moteur dans le cas des essais purs-corrects ... 101
II.3.3.6 Proportion des essais incorrects-corrects ... 102
II.3.3.7 Taux d’ébauche d’erreur ... 103
II.3.3.8 Temps de correction ... 104
II.3.3.9 Corrélation entre l’enveloppe de la bouffée EMG correspondant à l’ébauche d’erreur et le temps de correction ... 105
II.3.3.10 Latence des activités incorrectes ... 107
II.3.4 Discussion ... 108
Chapitre.III. Discussion générale ... 114
PERSPECTIVES DE TRAVAIL... 123
BIBLIOGRAPHIE... 125
INDEX DES FIGURES ... 137
INDEX DES TABLEAUX... 142
INTRODUCTION GÉNÉRALE
Ces travaux de thèse s’inscrivent dans le cadre théorique de l’étude du traitement de l’information. L’être humain est assimilé à une machine à traiter de l’information. Les informations présentées au sujet représentent les entrées du système et la production d’une réponse motrice constitue la sortie du système. La structure interne de ce système est constituée d’une série d’étapes de traitement qui ne sont pas directement observables. Cependant, plusieurs méthodes expérimentales permettent d’appréhender la nature spécifique des opérations qui se déroulent entre la présentation d’un stimulus et la production d’une réponse motrice. La chronométrie mentale est le paradigme qui permet d’étudier les différentes étapes du traitement de l’information par l’intermédiaire de la mesure du temps de traitement des processus mentaux.
Dans notre vie quotidienne, et a fortiori dans la plupart de nos activités physiques et sportives, on est amené à prendre des décisions en un temps limité afin de s’adapter aux différentes éventualités possibles. Le système sensori-moteur, avec les informations qu’il a à sa disposition, doit alors préparer les mouvements (programmation anticipée) dans le but d’optimiser la performance. Mais il arrive, afin que le comportement soit adapté à l’environnement, qu’il doive rapidement déprogrammer ce qu’il a traité et reprogrammer de nouveaux mouvements car de nouvelles informations, ne correspondant plus à ce qu’il avait prévu, lui parviennent. Par exemple un joueur de basket-ball se prépare à envoyer la balle à son partenaire qui se trouve sur sa gauche. Cependant, le déplacement d’un adversaire à proximité de celui-ci change la situation sur le terrain. Par conséquent, pour protéger la balle le joueur devra inhiber (déprogrammer) son mouvement planifié et en programmer un autre plus adapté à cette nouvelle situation : il peut soit renvoyer la balle vers un autre partenaire se trouvant du côté droit, soit avancer en dribblant pour marquer un point.
En fait, pendant la programmation d’une réponse motrice, l’objectif abstrait de l’action est traduit en programme moteur structuré par un ensemble des commandes musculaires (Keele, 1968, 1981). L’activation d’un programme moteur nécessite la spécification de différentes dimensions qui constituent le mouvement sollicité (Rosenbaum 1983) ; même un simple mouvement de pointage exige la spécification des différents paramètres comme : l’effecteur, la direction, l’amplitude, la durée et la vitesse. L’objet de ce travail est de préciser la nature des opérations cognitives impliquées dans la préparation des différents paramètres du mouvement et ainsi de tenter d’expliquer les processus de
déprogrammation-savoir si notre système, face à une situation différente de celle qu’il avait prévue, déprogramme la totalité de ce qu’il avait traité et reprogramme de nouveaux mouvements ou s’il réalise une déprogrammation partielle, conservant les paramètres du mouvement corrects et ne reprogrammant par conséquent que les dimensions du mouvement qui ne sont plus adéquates.
La première partie de ce manuscrit est consacrée à la présentation des fondements théoriques de ce travail de thèse. La seconde partie, qui s’appuie sur le cadre méthodologique défini par Rosenbaum (1980), Rosenbaum et Kornblum (1982), présente en détail chacune des trois expérimentations réalisées dans le cadre de ce travail, ainsi que les résultats obtenus. Enfin, la dernière partie consiste en une discussion générale.
I.1.
Système du traitement de l’information
La théorie du traitement de l’information constitue la base originelle de la psychologie cognitive. Elle suppose que les organismes vivants reçoivent des informations à partir de leur environnement et les saisissent de façon sélective par leurs organes récepteurs ; ces informations sont alors transformées et conduisent à certains comportements.
Chez les organismes supérieurs, et particulièrement chez l’Homme, ces transformations deviennent très complexes. Elles donnent lieu à des formes diverses de stockage de l’information ; la plus élaborée est celle qui constitue des représentations à long terme de l’environnement et de ses parties. Le traitement de l’information se présente alors sous la forme d’une suite de transformations de représentations transitoires en des représentations plus complexes.
Shannon et Weaver en (1949) ont proposé la théorie de la communication qui fournit pour la première fois une définition quantifiée de la notion d’information. Cette théorie présente un schéma minimal de la communication : la source émet des messages qui sont codés par le transmetteur en signaux susceptibles d’être véhiculés par un canal, eux-mêmes décodés par le récepteur sous une forme assimilable par le destinataire. La théorie de la communication quantifie l’information comme une fonction croissante de la réduction d’incertitude qu’elle apporte. Dans cette perspective elle a fourni une mesure de quantité d'information dont l'unité est le bit (Binary Digit). Le bit, c'est la quantité d'information contenue dans le choix élémentaire entre deux possibilités équiprobables, comme dans le cas du lancer d'une pièce de monnaie pour obtenir pile ou face, (une réduction de moitié de l’incertitude apporte un bit d’information).
Dans la plupart des théorisations actuelles, descendantes plus ou moins lointaines de la théorie de la communication de Shannon et Weaver, il est admis que l’information apportée par un signal physique, généralement un stimulus externe, subit une série de transformations pour aboutir à la production d’un autre événement physique, généralement une réponse motrice. Selon les modèles, les différentes opérations sont regroupées en étapes de traitement ayant chacune une fonction spécifique. On distingue à l’entrée, les processus responsables de l’analyse du signal (Input), à la sortie, les processus responsables de l’élaboration de la
subdivision des processus d’entrée et des processus de sortie en une étape périphérique (neurorécepteurs, neurones responsables à transmettre le message nerveux aux effecteurs) et une étape centrale (le cerveau et la moelle épinière). Si on se met d’accord sur le fait que ces différentes étapes débutent de façon sérielle (les processus d’entrée commencent avant les processus associatifs dont le début précède le début des processus de sortie), il reste à savoir la façon dont les étapes s’organisent dans le temps. Les premiers grands modèles de la psychologie cognitive, par exemple, le modèle de Donders (1868) ont supposé que les processus cognitifs étaient réalisés de façon séquentielle par une suite de transformations successives. Selon cette conception, chaque étape attend d’avoir terminé son travail avant d’en transmettre le produit à l’étape suivante ; une seule étape est active à un instant donné. Il n’y aurait alors aucun recouvrement temporel entre les opérations réalisées au cours des différents processus de traitement. Les ordinateurs classiques, qui ne comportent qu’un seul processeur, procèdent de cette façon. L’opposé du traitement séquentiel est le traitement en parallèle (e.g., le modèle continu de MacClelland, 1979) où toutes les étapes du traitement peuvent être actives simultanément, les informations partielles sont transmises au fur et à mesure de leur élaboration, donc il y aurait un recouvrement temporel entre les opérations réalisées au cours des différents processus de traitement. Les ordinateurs élaborés depuis quelques années comportent une multiplicité de processeurs qui fonctionnent de façon simultanée et se partagent les traitements.
L’un des objectifs de la psychologie cognitive est de comprendre la nature spécifique des processus du traitement de l’information, c’est pourquoi elle s’efforce de modéliser ces différentes étapes. Une succession d’opérations cognitives est nécessaire à la mise en place d’un comportement. Classiquement il est nécessaire de percevoir une situation, prendre des informations dans son environnement, identifier et interpréter ces informations avant de prendre une décision et agir de manière adaptée aux contraintes de la situation et à ses évolutions préalables. De nombreux modèles du traitement de l’information comprenant un nombre plus ou moins important de stades ont été proposés pour déterminer les opérations mentales élémentaires (ou « processus ») nécessaires à l’élaboration d’un comportement. Une des caractéristiques majeures de ces modèles est de considérer les processus mentaux comme une succession d’étapes, chacune consacrée à une fonction particulière. Les trois principales fonctions de ce modèle sont : la perception du stimulus, la sélection de la réponse et la programmation du mouvement (Schmidt, 1988 ; Theios, 1975). Les opérations perceptives prélèvent les informations dans l’environnement et les encodent en vue d’un traitement. Les opérations décisionnelles constituent la traduction stimulus-réponse. Elles déterminent la
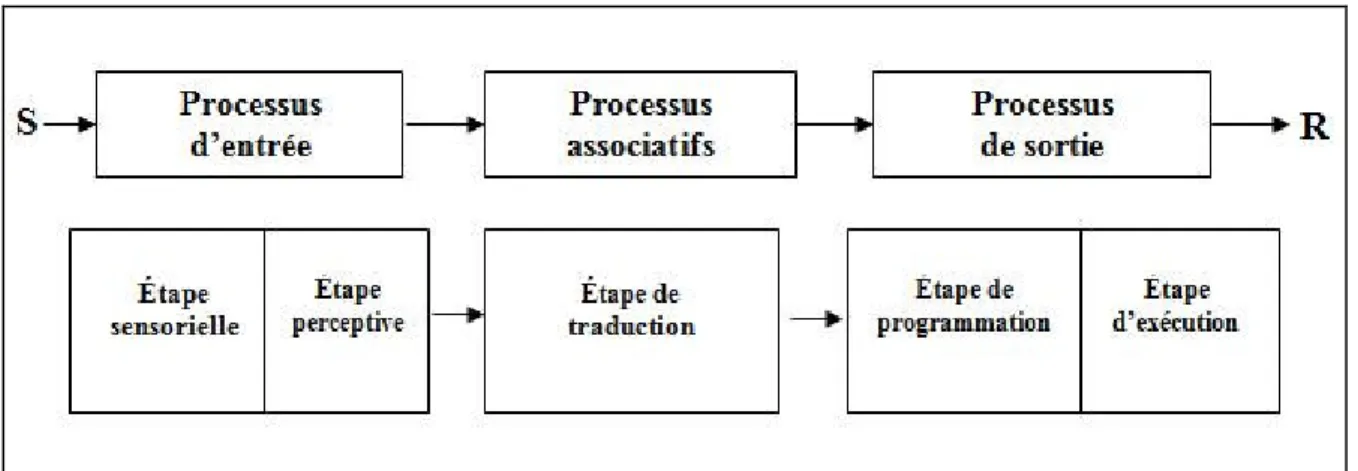
réponse appropriée au stimulus présenté. Les opérations motrices structurent et organisent les commandes nécessaires à l’exécution de la réponse sélectionnée (Klapp, 1978). Le modèle auquel il est le plus souvent fait référence dans la littérature, est celui proposé par Theios (1975). Ce modèle décrit le traitement de l’information en cinq étapes (figure1) :
1. une étape sensorielle (processus d’entrée périphérique) qui permet de détecter un événement dans l’environnement ;
2. une étape perceptive (processus d’entrée central) qui permet d’identifier cet événement ;
3. une étape centrale de traduction stimulus – réponse qui permet de choisir la réponse à exécuter, étant donnée la nature de l’environnement ;
4. une étape de programmation de la réponse (processus central de sortie) : Les opérations de programmation de la réponse ont pour objet l’organisation du système moteur pour contrôler le mouvement désiré.
5. une étape d’exécution de la réponse (processus de sortie périphérique) qui permet de contracter les muscles appropriés pour exécuter le mouvement désiré.
Figure 1 : Modèle montrant les principales étapes de la chaîne de traitement de l’information
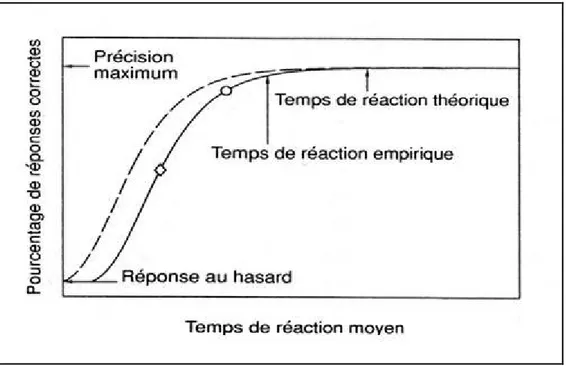
Pour comprendre la nature spécifique des opérations se déroulant dans le système de traitement de l’information et pour en inférer leur organisation temporelle, différentes méthodes ont été utilisées comme les méthodes psychophysiques et les indices physiologiques (l’électromyographie par exemple), mais l’outil méthodologique le plus largement utilisé pour étudier ces processus est le paradigme de la chronométrie mentale. Dans ce paradigme, la contrainte supplémentaire est que la réponse correcte doit être effectuée le plus rapidement
I.2. Paradigme de la chronométrie mentale
La chronométrie mentale peut être définie comme l’étude de la séquence temporelle du traitement de l’information dans le cerveau humain, ou autrement dit, comme une mesure précise des processus psychologiques. La chronométrie mentale, qui a trouvé une application dans diverses disciplines, notamment la psychophysiologie cognitive, les neurosciences cognitives et les neurosciences comportementales, est l’un des principaux paradigmes de la psychologie cognitive et expérimentale. Les psychologues ont mis au point et affiné la chronométrie mentale dans ces 100 dernières années en utilisant le temps de réaction comme variable dépendante. En fait la variable comportementale clé de la chronométrie est le temps de réaction (TR). D’un point de vue théorique, le TR reflète la durée des processus de traitement de l’information. L’intérêt principal de l’étude du TR consiste à préciser la nature et le nombre des stades internes, inobservables en tant que tels, qui se déroulent entre l’occurrence d’un stimulus et la production d’une réponse permettant ainsi de décrire la structure du système de traitement de l’information implémenté dans le cerveau.
I.2.1
Procédure du temps de réaction
Le temps de réaction (TR) commence avec la présentation du stimulus (visuel, auditif, tactile,…) et se termine avec l’exécution du mouvement. Dans la procédure de tâche du TR (figure 2), le signal de réponse (SR), auquel le sujet doit répondre le plus rapidement et le plus précisément possible, est précédé d’un signal avertisseur. L’intervalle de temps séparant le signal avertisseur du SR est communément appelé période préparatoire pour des raisons qui seront développés ultérieurement. Ce dernier informe que la survenue du SR est imminente, permettant ainsi au sujet d’augmenter son niveau de préparation. Lors d’une tâche de TR, il est important que les sujets respectent la consigne de réagir le plus rapidement possible tout en évitant les erreurs. Lorsque ces consignes sont respectées, on considère que le temps qui s’écoule entre le SR et l’exécution de la réponse correspond au TR et représente le temps minimal requis pour effectuer l’ensemble des opérations conduisant à la réponse adéquate.
Figure 2 : Procédure chronométrique d’analyse du TR
I.2.2
Différentes tâches de temps de réaction
Parmi les différentes tâches de TR on distingue les tâches de TR simple (TRS), et les tâches de TR de choix (TRC). Dans une tâche de TRS, un stimulus unique est associé à une réponse unique, donc il n’y a pas d’incertitude événementielle ni sur le SR ni sur la réponse. Le sujet est confronté à une situation où il sait quelle réponse il doit réaliser, toutes ses caractéristiques étant connues avant le SR. D’un point de vue théorique, on peut avoir une incertitude temporelle dans la tâche de TRS, dans ce cas l’incertitude porte sur le moment d’occurrence du stimulus, ce qui minimise le risque que le sujet anticipe la réponse en synchronisant sa réponse avec le SR pour diminuer le TR. Une tâche de temps de réaction du choix (TRC) se compose de plusieurs stimuli et de plusieurs réponses. Généralement, une réponse spécifique est associée à chacun des stimuli possibles. Le sujet ne sait pas à l’avance quel sera le mouvement à produire, cette information étant contenue dans le stimulus. Pratiquement, dans la tâche de TRC où l’incertitude est événementielle, on peut jouer aussi sur l’incertitude temporelle.
Les premières études utilisant le temps de réaction (TR) ont été menées par Donders (1868-1969). Son travail tentait de décrire les processus qui se déroulent dans le cerveau. Particulièrement, il s’est intéressé à mesurer le temps nécessaire pour effectuer les différentes opérations de traitement mental. Donders a utilisé la technique de soustraction pour mesurer le temps des différents processus mentaux face à différentes tâches de TR. Il a utilisé trois
Tâche (a) qui correspond à une tâche de TRS (le stimulus est la syllabe « ki » et le sujet doit répéter cette syllabe) où seules les étapes « détection » du stimulus et « exécution » de la réponse sont mises en jeu.
Tâche (b) qui correspond à une tâche de TRC (le stimulus est l’une des cinq syllabes suivantes [ka, ke, ki, ko, ku] et le sujet doit répondre en répétant cette syllabe) : dans cette tâche toutes les étapes de traitement de l’information sont présentes « détection » et « discrimination » du stimulus, « sélection » et « exécution » de la réponse.
Tâche (c) qui correspond à une tâche Go-No go (le stimulus est à nouveau une des 5 syllabes, mais le sujet ne doit répéter que la syllabe « ki ») où seulement les étapes « détection » et « discrimination » sont sollicitées.
La différence entre les temps de réaction observés dans la tâche (c) et ceux observés dans la tâche (a) fournit une estimation du temps de discrimination et la soustraction entre les temps de réaction observés dans la tâche (b) et ceux de la tâche (c) donne une estimation du temps de sélection de la réponse (Figure 3).
Figure 3 : Méthode soustractive de Donders.
Le TR est typiquement plus rapide en condition de TRS qu’en condition de TRC. Cette différence est notamment imputée au temps supplémentaire requis pour sélectionner et programmer la réponse appropriée en condition de TRC (Theios, 1975).
Dans ce contexte, le modèle du système de traitement de l’information constitue un cadre de référence pertinent et la chronométrie mentale un outil indispensable à l’étude des processus mentaux sous-jacents à la production d’une réponse motrice. La procédure chronométrique
soutient l’idée selon laquelle le mouvement est organisé avant son déclenchement. Le système moteur doit retrouver et organiser un programme moteur qui par la suite contrôlera le mouvement et il doit diriger les muscles pour que ceux–ci se contractent dans l’ordre approprié, avec le degré de force voulu et au moment adéquat, pour produire le mouvement de façon effective. Cette conception d’une organisation anticipée du mouvement est apparue comme une des plus importantes dans le domaine de la programmation motrice (Keele, 1968). Les auteurs qui s’inscrivent dans ce courant de recherche postulent que la notion de programmation motrice occupe une place centrale dans le contrôle de l’action.
I.3. Programmation motrice
L’apparition de la notion de programme moteur date des années soixante (e.g., Henry & Rogers, 1960). L’idée à l’origine de cette apparition concerne la traduction des intentions du sujet en actes moteurs. Le programme moteur est considéré comme l’instance assurant cette traduction et constitue l’interférence entre “l’idée motrice” et l’action (Rosenbaum, 1985). Cette instance est qualifiée de mémoire globale du mouvement (Henry, 1961 ; Henry & Rogers, 1960 ; Keele, 1968). En 1961, Henry définit un programme moteur comme une mémoire motrice ou encore un programme neuromoteur stocké.
Le concept de programme moteur a été proposé consécutivement à des expériences de désafférentation. La désafférentation se traduit par une interruption de la transmission des informations sensorielles de la périphérie vers le système nerveux central par les voies afférentes. Le flot des informations peut-être interrompu soit de manière temporaire (e.g., anesthésie locale), soit de manière irréversible (e.g., section des nerfs). Depuis les études menées chez des singes désafférentés chirurgicalement (e.g., Polit & Bizzi, 1978), on sait qu’il est possible d’exécuter des mouvements avec un membre désafférenté. Le contrôle du mouvement peut ainsi s’effectuer sans qu’il n’y ait de retour sensoriel (feedback) en provenance du membre impliqué dans le mouvement. Ces données suggèrent l’existence de représentations internes qui permettent d’organiser le mouvement volontaire en l’absence d’afférences. Le travail de Keele (1968) est considéré comme un des plus influents en matière de conceptualisation du programme moteur. Keele propose deux arguments en faveur de la programmation motrice dans le contrôle du mouvement rapide. Le premier est que le
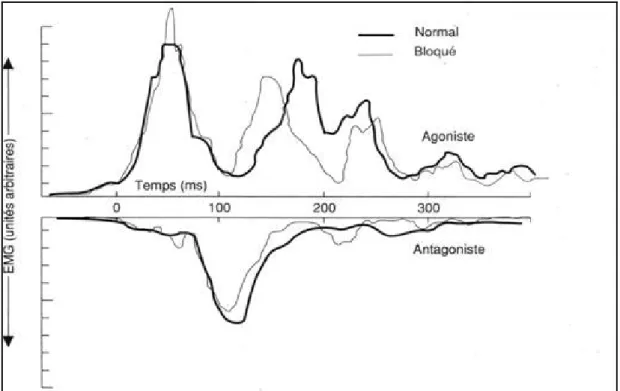
(1979). La tâche consiste à exécuter un mouvement rapide de segment (actionner un levier vers une cible). Ils ont recueilli le pattern d’activités des muscles agoniste et antagoniste impliqués dans le mouvement à travers des enregistrements électromyographiques (EMG). Lors d’exécution du mouvement on observe d’abord une bouffée d’activité du muscle agoniste (triceps) qui met en mouvement le membre pour l’amener vers la position finale, ensuite cette bouffée prend fin et pendant ce temps apparaît une bouffée du muscle antagoniste (biceps), vraisemblablement pour ralentir le membre, et finalement l’agoniste est réactivé vers la fin du mouvement pour stabiliser le segment au niveau de la cible. Ce pattern triphasique est caractéristique des mouvements rapides de ce type (en traits gras sur la figure 4). Dans certains essais, le levier était bloqué dans sa position initiale de manière inattendue, de telle sorte qu’aucun mouvement ne pouvait survenir. Les données EMG enregistrées dans cette condition bloquée (trait fin sur la figure 4) révèlent que pendant les 100-150 premières ms, les tracés EMG en condition bloquée sont similaires à ceux en condition normale ; le pattern d’activité musculaire agoniste et antagoniste intervient au même moment pour les deux conditions. Ces résultats supportent l’hypothèse qu’au moins une partie du mouvement est contrôlée par un programme central et que les détails de l’action sont spécifiés à l’avance et stockés en mémoire. Ces caractéristiques sont celles du programme moteur.
Figure 4 : Activité EMG des muscles agoniste (triceps) et de l’antagoniste (biceps). Les tracés fins représentent l’enregistrement d’un mouvement qui a été bloqué mécaniquement au départ et les tracés gras l’enregistrement d’un mouvement rapide d’extension du coude (Reproduit d’après Wadman, Denier van der Gon, Geuze & Mol, 1979).
Les premières recherches chronométriques consacrées à l’étude des processus de programmation motrice ont utilisé une procédure de TRS (Henry & Roger 1960). Les résultats de cette étude montrent que le TRS augmente proportionnellement à la complexité du mouvement. Cette augmentation représente le temps nécessaire aux processus mentaux pour élaborer les commandes motrices du mouvement à réaliser. Ces auteurs considèrent que le temps de programmation d’un mouvement est déterminé par le nombre de groupes musculaires sollicités et le nombre de centres de coordination neuromoteurs requis. Dans ce cadre d’analyse, on considère que c’est le programme moteur qui détermine le nombre et la nature des muscles à contracter ainsi que l’ordre et la durée de leur contraction.
La procédure de TRS présente l’inconvénient d’inciter le sujet à anticiper l’apparition du stimulus et synchroniser sa réponse avec ce dernier. Le mouvement à réaliser est en effet connu à l’avance et le moment d’apparition du stimulus est prédictible, donc le SR est un signal de synchronisation pour déclencher la réponse motrice, et le TRS ne mesure pas précisément le temps de détermination des commandes motrices.
MacKenzie, 1981). Cette procédure confronte le sujet à une situation où l’incertitude sur le mouvement à réaliser n’est levée qu’à l’apparition du SR. Les résultats de ces travaux confirment ceux obtenus avec une procédure de TRS ; le TRC augmente avec la complexité motrice (Klapp 1977, Klapp et al., 1976, 1974, 1979). Rosenbaum (1980) envisage deux types d’interprétation à ce résultat. La première interprétation fait référence à une conception de sélection du programme moteur et la deuxième à une conception de paramétrisation du programme moteur. Soit le sujet sélectionne en mémoire autant de programmes moteurs qu’il y a de réponses possibles et charge celui qui correspond au SR. Soit il ne sélectionne qu’un programme moteur, puis détermine les commandes motrices constitutives de ce programme pour l’adapter aux exigences environnementales.
La conception d’une sélection du programme moteur assimile le système nerveux à un énorme réservoir contenant une représentation de chaque mouvement que le sujet est capable d’exécuter. Selon ce modèle, la programmation du mouvement dépend d’un processus de sélection du programme moteur lui correspondant. Le réservoir, permettant cette sélection, est activé pour répondre au besoin immédiat du système. Cette théorie met en relation deux notions contradictoires : la capacité du sujet à produire un nombre infini de mouvements et la capacité limitée du stockage en mémoire d’un grand nombre de programmes moteurs. Il semble, en effet, invraisemblable que le système nerveux soit en mesure de conserver la représentation complète de chaque mouvement qu’il est possible de réaliser.
La conception paramétrique de la notion de programmation motrice s’inscrit dans le cadre d’une analyse proposée par les modèles sériels de la théorie de traitement de l’information (e.g., Theios, 1975). Ces modèles suggèrent que les processus impliqués dans la programmation d’une réponse motrice correspondent à des opérations appartenant au versant moteur du traitement qui sélectionnent et organisent la réponse requise (Glencross, 1979 ; Klapp, 1978). Cette modélisation suppose qu’une fois le mouvement choisi, une traduction de cette intention motrice en commandes motrices est réalisée. Ces commandes motrices prennent alors la forme d’un programme moteur ajusté aux conditions environnementales. Dans ce cadre conceptuel, le modèle paramétrique proposé par Rosenbaum (1980, 1983) distingue deux niveaux hiérarchisés de processus centraux responsables du contrôle anticipé du mouvement. Dans un premier niveau, un programme moteur serait sélectionné parmi un nombre limité de programme stockés en mémoire à long terme. Ce programme moteur est appelé prototype du mouvement ou protoprogramme. Dans un second niveau, ce protoprogramme serait complété en fonction des exigences environnementales par une spécification des différentes valeurs le composant. Dans cette perspective, un mouvement
peut être, à partir de son protoprogramme, conservé en mémoire et réalisé dans une infinité de conditions différentes. Plus récemment, une autre proposition du programme moteur a été suggérée par Schmidt (1988) (le programme moteur généralisé ; PMG) pour répondre aux deux problèmes trouvés dans le concept du programme moteur. Le premier concerne le stockage des programmes moteurs qui nécessite des capacités de stockage improbables. Le deuxième concerne la nouveauté ; on est capable de produire de nouvelles actions qui ne sont pas représentées dans un programme moteur déjà présent en mémoire. Le PMG est une représentation abstraite stockée en mémoire, permettant de réaliser un nombre important de modifications d’une même action. Aussi il permet de produire des mouvements appartenant à une même classe d’action (e.g., réaliser une signature sur un chèque ou sur une tableau ; dans les deux cas on a le même pattern de résultats alors que les groupes musculaires qui sont sollicités sont différents).
Malgré quelques divergences liées au mode d’élaboration du programme moteur en situation de TRC (sélection et paramétrisation), l’ensemble des auteurs s’accorde sur la nature des commandes motrices le composant. L’idée partagée est que la sélection d’une réponse entraîne la spécification des caractéristiques du mouvement (Glencross, 1979 ; Klapp, 1978 ; Roenbaum, 1980, 1983). Le programme moteur serait ainsi formé par un ensemble d’instructions déterminant les caractéristiques spatio-temporelles et cinématiques d’un mouvement (Lépine, Glencross, & Requin, 1989). Les instructions du programme moteur concernent les dimensions temporelles en termes de durée, de vitesse et les dimensions cinématiques en termes de force requise. Ces dimensions sont qualifiées de paramètres du mouvement. Les paramètres sont sélectionnés dans des registres moteurs en fonction de l’état initial du système musculaire et de l’objectif de l’action, assemblés pour construire le programme, puis transmis aux effecteurs périphériques (Lépine et al., 1989). Dans cette perspective, les auteurs jouent sélectivement sur le nombre, la nature et/ou la connaissance des paramètres du mouvement.
I.4. Préparation motrice
En neurosciences cognitives, on considère que le rôle des processus préparatoires est de pré-activer certaines régions cérébrales afin d’améliorer les processus de traitement de
I.4.1
Définition
Les processus préparatoires permettent une amélioration de la performance généralement appréciée par une diminution du temps de réaction. Cette diminution du TR peut résulter de la mise en jeu de deux types de processus préparatoires : des processus qui consistent à effectuer certaines opérations pendant la période préparatoire (PP), réduisant ainsi le nombre d’opérations à effectuer après la présentation du SR, et des processus qui modulent, pendant la PP, l’état fonctionnel des systèmes de traitement responsables des opérations qui suivent l’apparition du SR, conduisant ainsi à une diminution de leur durée effective d’exécution. Requin, (1985), Requin, Brener et Ring (1991), ont défendu l’idée que les processus préparatoires regroupent deux types de processus distincts : la préparation temporelle non-spécifique et la préparation événementielle spécifique.
La préparation temporelle vise à résoudre le problème du « quand » : à quel moment faut-il exécuter une action. Ce problème est important, car on sait qu’un haut niveau de préparation non-spécifique ne peut ni être atteint immédiatement ni être maintenu longtemps. Un exemple prototypique est la phase de départ dans la compétition de sprint ; le TR au signal de départ sera diminué selon la qualité de l’anticipation de l'athlète en ce qui concerne le moment de son apparition. Au contraire, la préparation spécifique qui vise à résoudre le problème du « quoi » (quelle réponse produire ?), peut être maintenue pendant très longtemps. Comme exemple de ce type de préparation, on peut évoquer un joueur de tennis qui attend de la part de son adversaire un coup droit frappé, prépare ses déplacements de telle sorte qu’il puisse se trouver en bonne position quand il recevra la balle. Ce type de préparation est lié à l’incertitude événementielle sur la nature du signal et donc de la réponse associée ; elle rend compte du fait que le TR est d’autant plus court que l’identité et la localisation du signal d’exécution sont prévisibles (Holender, 1980). Elle correspondrait à une activation préalable des modules de traitement impliqués dans l’analyse de l’événement attendu et dans la sélection et la programmation du mouvement qui lui sont associées. En d’autres termes, la préparation spécifique rend possible la disponibilité en mémoire à court terme, au moment du SR, des attributs qui caractérisent l’événement à identifier et des paramètres du mouvement à réaliser.
Il est évident que la préparation à l’action comprend des processus de planification, qui ajustent notre système sensori-moteur aux exigences environnementales. Les processus mentaux sous-jacents à la préparation à l’action ne sont pas directement observables. C'est
pourquoi, depuis longtemps les psychologues visent à découvrir ces processus cachés. Les psychologues et les spécialistes en neurosciences ont utilisé la technique de pré-signalisation (precuing) qui a été présentée par Leonard (1953), puis développée par Rosenbaum (1980) pour inférer les mécanismes sous-jacents à la préparation à l’action.
Dans l’expérience rapportée par Leonard en 1953, l’information préalable était totale : elle définissait complètement le mouvement qui devait être fait. Il s’agissait d’une tâche de TR continu dans laquelle le sujet devait manipuler un levier pour atteindre le plus rapidement possible l’angle d’un triangle où une lampe était allumée. Sitôt cette cible atteinte, la lampe s’éteignait et elle était remplacée par une autre lampe vers laquelle le sujet devait à nouveau pointer le levier (condition sans information préalable). Dans une autre condition, lorsque le sujet était en train de pointer le levier vers la cible, une lampe s’allumait dans un autre angle du triangle indiquant au sujet quelle allait être la cible suivante (condition avec information préalable). Le résultat important est que le TR était plus rapide dans la condition avec information que dans la condition sans information. Une interprétation possible de ce résultat est que, lorsque le sujet reçoit une information préalable, il est capable de l’utiliser pour programmer en avance le prochain mouvement et ainsi gagner un temps précieux.
I.4.2
Paradigme de pré-signalisation (precuing)
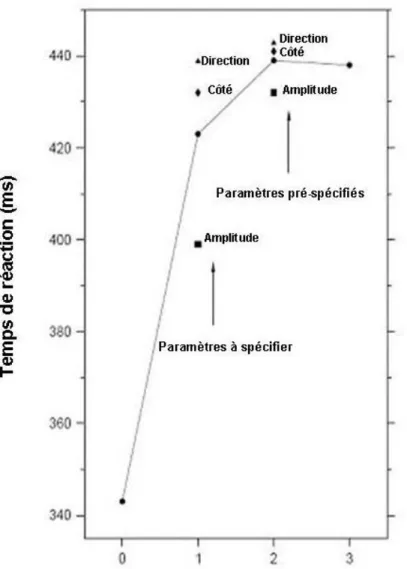
Quelques années plus tard, dans le cadre de son travail de thèse, Rosenbaum (1980) a étudié l’effet d’une information préalable dans le cas où celle-ci apporte non pas une information totale, mais une information partielle. Son idée a consisté à utiliser des réponses pouvant être spécifiées par la combinaison d’un certain nombre de paramètres indépendants. Dans son expérience, il s’agissait du côté (main droite ou gauche), de la direction (mouvement d’adduction ou abduction), et enfin de l’amplitude (petite ou grande) ; ce qui donne donc huit réponses possibles (e.g., un mouvement d’adduction de grande amplitude réalisé avec la main gauche) associées chacune à un SR différent. L’information donnée au sujet préalablement à la présentation du SR pouvait porter sur zéro, un, deux ou trois paramètres : par exemple la réponse pouvait être un mouvement d’abduction (information sur le seul paramètre de direction du mouvement), ou bien un mouvement d’abduction de petite amplitude (information à la fois sur la direction et l’amplitude du mouvement), ou encore un
Il ne lui reste alors à programmer pendant la phase de réponse que les paramètres restants du mouvement. Cette technique repose sur l’hypothèse que le processus de programmation consiste en un assemblage d’opérations séparables qui spécifient les dimensions du mouvement et qui ont chacune une durée mesurable. En effet, Rosenbaum propose le modèle paramétrique de la programmation du mouvement dans lequel l’ensemble des opérations de processus de programmation est transformé d’une façon sérielle parce que le temps de programmation est additif avec le nombre de paramètres pré-signalés. Supposons le cas où l’information porte sur zéro paramètre (information préalable nulle) ; à l’apparition du SR, le sujet doit programmer chacun des trois paramètres, temps de programmation qui contribue à la durée totale du TR. Supposons maintenant que le SP informe sur le côté et sur la direction ; pendant la PP, le sujet peut préprogrammer ces deux paramètres qui ne laissent que l’amplitude à programmer pendant le temps de réaction. Dans ce cadre, l’interprétation de Rosenbaum est donc que l’information préalable affecte les processus moteurs. S’il est possible d’apporter un certain nombre d’arguments en faveur de cette interprétation (e.g., Rosenbaum, 1980 ; Miller, 1982 ; Bonnet, Requin, & Stelmach, 1982 ; Larish & Frekany, 1985 ; Possamaï, Burle, Osman, &, Hasbroucq, 2002), d’autres auteurs, notamment Goodman et Kelso (1980) soutiennent que l’effet de réduction du TR, lorsque la quantité d’information préalable augmente, a pour origine la réduction d’incertitude apportée par cette information. Pour reprendre l’exemple précédent, dans la situation où le SP n’apporte aucune information (zéro bit), celle apportée par le SR est de trois alors que dans la situation où le SP spécifie le côté et la direction, soit une information de deux bits, le SR n’apporte plus qu’un bit d’information. Dans ce cadre théorique, l’effet de l’information préalable porte non pas sur les étapes motrices, mais sur les étapes décisionnelles centrales.
Par la suite, Rosenbaum et Kornblum (1982) ont proposé une autre approche, la technique d’amorçage (en anglais priming) qui permet de dépasser le problème méthodologique causé par la confusion de variable inhérente à la technique de pré-information, et qui permet en même temps de répondre à la problématique quant aux effets de l’information préalable.
I.4.3
Paradigme d’amorçage (priming)
Cette technique consiste à présenter un SP, appelé dans ce cas (amorce), qui fournit toujours une information totale concernant les dimensions du mouvement à exécuter. Cette amorce peut s’avérer valide, neutre ou invalide suite à la présentation du SR. Quand l’information est valide (majorité des essais), l’amorce annonce la même réponse que le SR.
Quand elle est neutre, aucune information sur la réponse à exécuter n’est donnée au sujet ; en d’autres termes l’amorce n’annonce pas une réponse. Enfin, dans le cas d’une information invalide, le SR annonce une autre réponse que celle signalée par l’amorce (figure 5)
Figure 5 : Procédure de la technique d’amorçage consiste à présenter une amorce qui est suivie après une période préparatoire par le signal de réponse qui peut être le signal amorcé (amorce valide) ou un autre signal (amorce non valide).
Lorsque l’amorce est valide, la réponse à exécuter est la réponse préparée. Par contre, lorsque l’amorce est invalide la réponse à exécuter diffère de la réponse préparée par un, plusieurs ou tous les paramètres. Cette procédure permet au sujet de préparer les dimensions spécifiées par l’amorce, parce qu’il y a une forte probabilité que le SR soit la réponse amorcée. Dans ce contexte, et lorsque l’amorce est invalide, le TR correspond au temps nécessaire pour modifier totalement ou partiellement les valeurs des paramètres du programme moteur amorcé. Le changement du TR est interprété par le temps nécessaire à déprogrammer et à reprogrammer les dimensions non valides du mouvement. Autrement dit, cette technique permet de mesurer le temps nécessaire pour modifier partiellement ou complètement un programme moteur incorrectement paramétré.
Dans ces dernières années également, les physiologistes et les neuroscientifiques se sont intéressés à mieux comprendre les mécanismes sous jacents à la préparation à l’action.
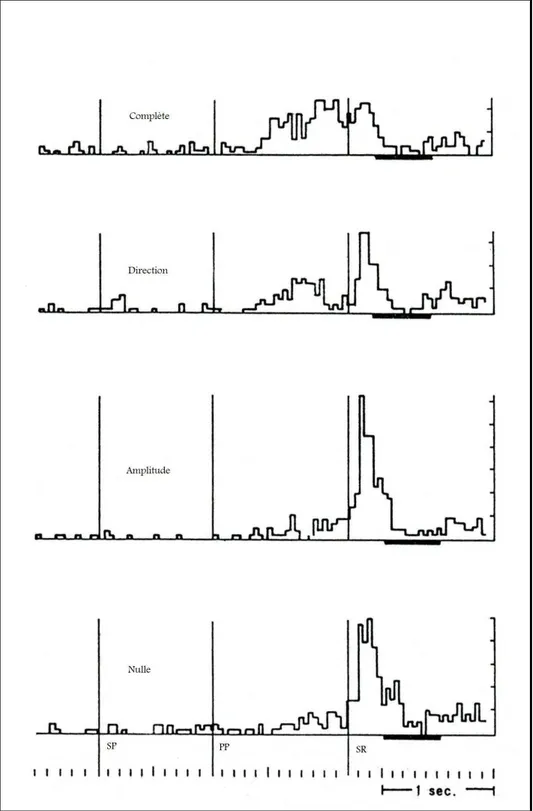
unitaire dans le cortex moteur primaire et pré-moteur de manière spécifique en fonction de la nature de cette information (amplitude, direction). La pré-signalisation de la direction provoquait, une diminution du TR et un changement dans l’activité neuronale pendant la PP par rapport à la condition où le SP apportait une information sur l’amplitude et celle où le SR n’apportait aucune information. Ainsi, le TR était plus court et l’activité neuronale était plus importante dans la condition où les deux paramètres étaient pré-signalés (figure 6). Deux mécanismes neurophysiologiques ont été mis en évidence dans cet effet de préparation motrice (Rhiele & Requin, 1989) :
1. Le premier serait responsable d’une prédisposition des aires motrices impliquées dans l’élaboration de la commande motrice. Celle-ci consisterait en une augmentation progressive, avant l’apparition du SR, de l’excitabilité des neurones spécifiquement impliqués dans la programmation de la direction du mouvement. Cette opération d’une façon non spécifique permet la facilitation de la programmation et donc la réduction du TR.
2. Le deuxième révèle un traitement anticipé plutôt qu’une facilitation globale. En d’autres termes, il consiste à programmer partiellement ou totalement le mouvement en avance, ce qui permet également de réduire le TR (Rosenbaum, 1980, 1983).
Figure 6 : Chez les singes l’information pré-signalée apportée par le signal préparatoire (SP) module l’activité neuronale unitaire pendant la période préparatoire (PP) en fonction de la nature et du nombre d’information (amplitude, direction,). L’activité neuronale est la plus importante lorsque le SP apporte des informations complètes. Cette activité est moins importante lorsque le PS apporte des informations sur la direction. L’activité neuronale est la même lorsque le SP apporte une information sur l’amplitude où lorsqu’il n’apporte aucune information. Figure adaptée de l’étude de Rhiele et Requin (1989).
les activités électroencéphalographiques liées à l’événement (en anglais : Event-related brain potential, ERP) pour analyser ses différentes composantes potentielles comme par exemple :
-
Le potentiel de préparation latéralisé (LRP ; Lateralized Readiness Potential). Dans ce cas, le tracé EEG présente, pendant la PP, une grande négativité au-dessus du cortex moteur controlatéral à la main impliquée dans la réponse (pour plus de détails, voir la discussion de la troisième étude) (e.g., Kutas & Donchin, 1980 ; Eimer, 1995 ; Eimer & Coles, 2003). Leuthold et Jentzsch (2001, 2002) ont réalisé une tâche de pré-signalisation pour laquelle le SP portait une information partielle, totale ou nulle sur la main impliquée dans la réponse (index droit ou gauche) et sur la direction du mouvement (extension ou flexion). Leurs résultats ont montré que l’amplitude du LRP était plus importante lorsque le SP fournissait une information totale que lorsqu’il fournissait une information partielle sur le doigt. Ils ont également montré une absence du LRP lorsque le SP portait sur une information partielle de direction.-
La variation contingente négative (CNV ; Contingent Negative Variation). Cette onde négative correspond à des activités qui se développent pendant la PP dans une tâche de TR (Walter, Cooper, Aldridge, McCallum, & Winter, 1964). La CNV comprend deux composantes : une onde précoce qui reflète les activités d’orientation vers le SP et une onde tardive qui reflète la préparation motrice. Dans une tâche de pré-signalisation, Mackay et Bonnet (1990) ont montré que l’amplitude de l’onde CNV était affectée par le nombre d’informations pré-signalées. Dans leur étude le SP fournissait une information totale, partielle, ou nulle sur la direction (flexion, extension), ou la force de la réponse (faible, forte). Contrairement au TR, l’amplitude de l’onde CNV augmentait avec le nombre des paramètres pré-signalés (Ulrich, Leuthold, & Sommer, 1998 ; Leuthold & Jentzsch, 2001).-
Dans d’autres études les auteurs se sont intéressés à d’autres composantes comme : ADAN (en anglais ; anterior directing attention negativity) et LDAP (en anglais ; late directing attention positivity). Généralement ces deux composantes sont analysées dans les études portant sur l’orientation de l’attention spatiale (e.g., Harter, Miller, Price, LaLonde, & Keyes, 1989 ; Hopf & Mangun, 2000 ; Eimer, Van Velzen, Forster, & Driver, 2003). En fait, lors de l’orientation de l’attention suite à la présentation d’une amorce spatiale, la composante ADAN présente 350 ms-600 ms, après la présentation de l’amorce, une augmentation de la négativité au-dessus des électrodes antérieures situées au côté controlatéral de l’amorce. Cette négativité est suivie après 600ms-900ms par la composante LDAN qui représente une grande positivité postérieure controlatérale au côté pré-signalé par l’amorce. Certaines étudespsychophysiologiques en utilisant la technique de pré-signalisation ont montré l’existence de ces deux composantes pendant la PP dans les tâches qui impliquent la préparation d’une réponse manuelle. Comme par exemple dans un simple mouvement de lever le doigt, Eimer, Foster, Van Velsen et Prabhu, (2005) et Eimer et Van Velzen, (2006) ont trouvé ces deux composantes lorsque l’amorce spécifie la main requise. Il en est de même de Gherri, Van Velzen et Eimer (2007) dans la préparation des réponses plus complexes (un mouvement de pointage) pour lesquelles différents paramètres doivent être spécifiés de façon indépendante : la main et la direction. Les auteurs ont montré que seule l’amplitude de la composante LDAP augmentait lorsque le SP fournissait des informations totales ou une information partielle sur la main impliquée dans la réponse, comparativement au cas où ce SP fournissait une information sur la direction.
Comme nous l’avons vu précédemment, la préparation motrice permet à certaines opérations de traitement de l’information d’avoir lieu en avance, ce qui évite qu’elles ne se déroulent après l’apparition du stimulus (pendant le TR). En conséquence, la préparation apporte un gros avantage pour la performance de notre action.
De façon intéressante, la préparation ne fait pas uniquement appel à des mécanismes d’amplification ou d’anticipation. Elle fait également appel à des mécanismes inhibiteurs. En effet, parallèlement à l’activation, la préparation requiert des processus d’inhibition afin d’éviter l’initiation de réponses erronées (Hasbroucq, Kaneko, Akamatsu, & Possamaï, 1997).
I.5. Inhibition
Dans les tâches de TRC, le terme d’inhibition est utilisé en référence aux mécanismes ou aux processus qui se mettent en place pour supprimer la réponse comportementale dominante lorsque cette réponse est anticipée de façon erronée. Différents modèles tentent d’expliquer les mécanismes sous-jacents à l’exécution de la réponse motrice requise, comme par exemple la théorie des accumulateurs de McClelland (1979). Le modèle proposé dans cette théorie comprend deux niveaux : un niveau constitué d’accumulateurs d’activation correspondant à chacun des stimuli et un niveau composé d’accumulateurs correspondant à chacune des réponses motrices possibles. Au cours de TR tous les accumulateurs sont activés par l’information apportée par les stimuli, bien qu’une seule réponse soit requise. La réponse
compétition de course. Cette compétition a été expliquée par le modèle de course (race model ; figure 7A ; Audley, & Pike, 1965 ; Vickers, 1970 ; Cohen, Dunber, & McClelland, 1990). Dans ce modèle les deux accumulateurs sont indépendants et le TR correspond au temps nécessaire à un accumulateur pour atteindre cette valeur seuil. Mais, il est difficile, sans introduire de la notion d’inhibition, d’expliquer les processus impliqués dans l’exécution de la réponse dans des tâches dites de « conflit » (Stroop, Simon, tâche d’Ericksen) ou dans des tâches de TRC où plus de deux réponses sont possibles (Usher & MacClelland, 2001 ; Burle, Vidal, Tandonnet, & Hasbroucq ; 2004). En effet, la notion d’inhibition est introduite comme un facteur fondamental dans ces types de tâche. À la suite de la définition des trois tâches de conflit citées précédemment, le rôle de l’inhibition dans ces tâches a été précisé. Dans les tâches de conflit comme la tâche de Stroop (Stroop, 1935), la tâche Simon (Simon, 1990) ou les tâches d’Eriksen (Eriksen & Eriksen, 1974), le stimulus qui provoque la réponse est bidimensionnel ; l’une des deux dimensions est pertinente avec la réponse requise et l’autre ne l’est pas.
Dans la tâche de Stroop, le sujet doit nommer la couleur d’un mot présenté. Lorsque la tâche est pertinente ; la couleur correspond au mot présenté (e.g., le mot rouge est écrit en couleur rouge), et lorsque la tâche est non pertinente la couleur ne correspond pas au mot (e.g., le mot rouge est présenté en bleu).
Dans la tâche de Simon, le sujet doit répondre avec la main droite ou avec la main gauche selon la couleur du SR qui est présenté soit à droite soit à gauche, (e.g., répondre avec la main droite pour un stimulus vert et répondre avec la main gauche pour un stimulus bleu). Si le stimulus vert est présenté du côté droit alors la condition est congruente. Par contre, si le stimulus bleu est présenté du côté droit alors la condition est non congruente.
Dans la tâche d’Eriksen, le sujet doit répondre par sa main droite ou gauche selon l’identification de la cible (A ou H). Cette cible est accompagnée par des distracteurs de chaque côté. Lorsque la cible et les distracteurs sont identiques, alors la condition est congruente (e.g., AAA) par contre lorsqu’ils ne sont pas identiques alors la condition est non congruente (e.g., HAH). Dans ces trois types de tâches, le temps de réaction est plus long dans le cas où la réponse correspond à la dimension ou à la condition non congruente. Cette augmentation de TR est interprétée comme étant due à des processus d’inhibition qui empêcherait l’exécution de la réponse incorrecte. Autrement dit, dans la condition non congruente, la dimension non congruente (e.g., le mot rouge est présenté en bleu, le stimulus vert est présenté à gauche, le cas HAH) active la réponse incorrecte. L’activation de la réponse incorrecte doit être inhibée pour exécuter la réponse correcte ; donc cette activation
sera en compétition avec la réponse correcte. Plus la réponse incorrecte est activée plus on a besoin du temps pour l’inhiber, par conséquent, plus le TR est long.
La notion d’inhibition est alors importante pour compléter le modèle des accumulateurs. Burle et collaborateurs (2004) décrivent principalement deux modèles de processus d’inhibition qui divergent sur le niveau du traitement responsable de l’inhibition.
Le premier est « l’inhibition latérale » : c’est l’inhibition générée par les accumulateurs eux-mêmes, autrement dit, par le niveau moteur. L’accumulateur correspondant à la réponse motrice requise reçoit une activation ; à ce niveau, cet accumulateur inhibe les autres accumulateurs (inhibe les réponses associées aux autres accumulateurs), donc la réponse sera exécutée lorsque le niveau d’activation d’accumulateur atteint la valeur du seuil prédéfini (figure 7B). Le deuxième modèle est « l’inhibition proactive » qui provient d’un niveau du traitement supérieur au traitement moteur. Chaque accumulateur reçoit à la fois des signaux excitateurs et inhibiteurs. C’est la somme de ces signaux qui va déterminer le niveau d’activation d’un accumulateur, et la réponse sera exécutée lorsque le niveau d’activation d’un des accumulateurs arrive au seuil prédéfini (figure 7C). Les auteurs ont cherché dans leur article à définir à la fois les sites générateurs de l’inhibition et les sites d’application de l’inhibition dans le but de tester les modèles d’inhibition latérale et proactive. Une tâche de TRC se caractérise d’abord par une activation bilatérale du cortex moteur primaire puis par l’inhibition des régions non impliquées dans la réponse requise. La moelle épinière ainsi que le cortex moteur primaire représentent les sites d’action de l’inhibition. Mais deux régions cérébrales sont essentiellement responsables de cette inhibition : le cortex cingulaire antérieur (CCA) et l’aire motrice supplémentaire (AMS). Cela conforte l’hypothèse d’un modèle d’inhibition proactif, car l’inhibition est contrôlée par des régions pré-motrices.
Les processus d’inhibition sont multiples. Certains auteurs ont tenté de catégoriser ces processus et de déterminer si ces niveaux d’inhibition sont contrôlés par des centres multiples. Logan (1994) défend l’existence de deux processus d’inhibition : un processus rapide et « non sélectif » inhibant toutes les réponses comme dans le cas de la tâche Stop (e.g. Van den Wildenberg, Van Boxtel, Van der Molen, & Logan, 2002, 2003 ; Van Boxtel, Van der Molen, Jennings, & Brunia 2001) ; Un processus plus lent permettant une inhibition « sélective », comme par exemple dans les tâches d’Ericksen où les sujets doivent inhiber la réponse activée par des distracteurs si cette dernière n’est pas compatible avec la réponse attendue. Dans la tâche Stop le sujet est soumis à une tâche de TR classique, un signal stop peut être présenté pour ne pas répondre. Le sujet doit alors interrompre la réponse qu’il était sur le point ou en train d’exécuter. Ces processus d’inhibition sont considérés comme non spécifiques car ils stoppent toute l’activité. Ce point de vue s’appuie sur certaines données de la littérature dans le domaine du contrôle moteur qui envisagent deux processus différents dans le contrôle du mouvement. Les études de De Jong, Coles, Logan et Gratton (1990) et De Jong, Coles et Logan (1995) proposent que l’inhibition du mouvement peut opérer soit en affectant l’élaboration des commandes motrices au niveau central, soit en affectant la sortie de la commande motrice vers la périphérie. Les auteurs distinguent donc deux processus d’inhibition : un processus d’inhibition sélectif contrôlé par les régions corticales et un processus périphérique rapide, non sélectif, contrôlé par les régions sous-corticales et fonctionnant en tout ou rien.
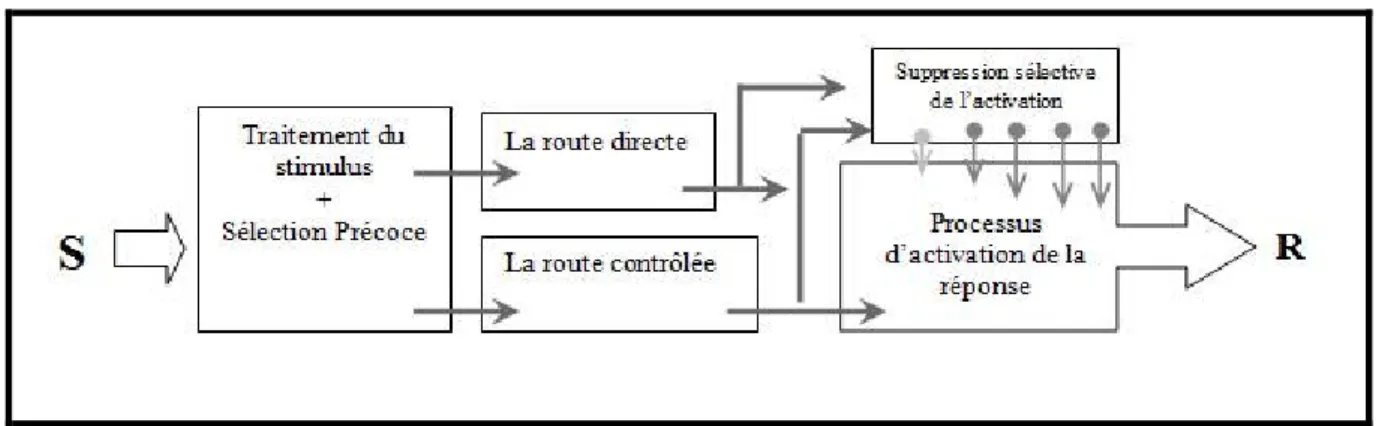
Certains auteurs se sont basés sur d’autres modèles pour expliquer la diminution de la performance dans la tâche de TRC et les tâches de conflit. Par exemple, Ridderinkhof en 2002 pour justifier son hypothèse « activation-suppression » s’est appuyé sur le modèle de double-routes. Dans ce modèle on distingue deux routes (voir, Kornblum, Hasbroucq, & Osman, 1990) ; une route délibérée et contrôlée active le code de la réponse correcte selon la consigne (la traduction S-R correspondant ou non correspondant), l’autre rapide et directe active le code de la réponse qui correspond au stimulus et son programme moteur (indépendant de la consigne de la traduction S-R) : par exemple, une flèche pointée vers le côté droit active la réponse qui implique la main droite, même si la consigne demande la réponse par la main gauche lorsque la flèche indique le côté droit. Lorsque les deux codes sont congruents, le programme moteur qui était activé par la route directe, est exécuté rapidement. Par contre, lorsqu’ils ne sont pas congruents, le programme moteur doit être abandonné en faveur de l’autre programme moteur qui en conséquence a besoin de temps pour être exécuté. Ridderinkhof a introduit dans ce modèle des processus de suppression sélectifs de cette
activation directe (figure 8). Cet auteur a utilisé dans une tâche de Simon l’analyse des distributions du TR et la technique de delta plot pour étudier le mécanisme de suppression de la réponse activée. Dans ce cas, le delta plot représente la taille de l’effet Simon. Dans une tâche de Simon, une route traite la position non pertinente du stimulus, alors que l’autre route traite la couleur pertinente du stimulus. Le traitement de la position est rapide et active automatiquement la réponse correspondante (la route directe). Par contre, le traitement de la seconde route permet d’activer la réponse qui correspond à la couleur du stimulus selon la consigne donnée (la route contrôlée). Ridderinkhof suggère l’existence d’une suppression sélective de la réponse activée directement par la dimension non congruente du stimulus. Cette suppression sélective aurait besoin de temps pour être élaborée. Ainsi, lorsque le TR est court, ce mécanisme n’a pas suffisamment de temps pour se mettre en place. Dans ce cas, la réponse correcte est complètement supprimée par l’activation de la réponse incorrecte. Par contre, lorsque le TR est long, l’activation de la réponse incorrecte est supprimée, ce qui réduit l’effet de Simon avec le temps (la pente de delta plot est plus négative).
Figure 8 : Représente une extension du modèle double routes avec les processus de la suppression sélective.
Des résultats expérimentaux confirment le modèle d’activation-suppression. Burle, Possama , Vidal, Bonnet et Hasbroucq (2002) ont utilisé le même modèle pour expliquer leurs résultats ; dans une tâche de Simon, le degré d’activation est mesuré par la présence d’une ébauche d’erreur (voir la section I.6.2.2). Ils ont montré que le degré d’inhibition de la réponse incorrecte est plus important dans les essais qui contiennent des activités incorrectes (ébauche d’erreur) par rapport aux essais sans ébauche d’erreur. Donc dans la condition incongruente plus la réponse incorrecte est activée plus la suppression de cette réponse
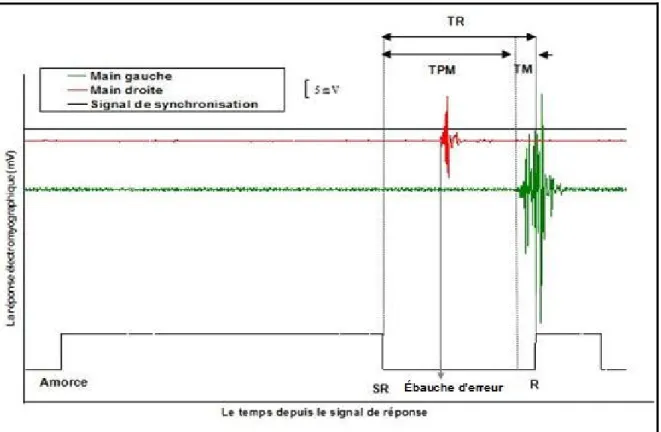
Les expériences présentées dans le cadre de ce mémoire reposent sur différentes méthodes d’inférence : l’analyse du TR moyen et l’analyse des erreurs, l’analyse des distributions du TR, le fractionnement du TR en deux composantes (temps pré-moteur, temps moteur), et l’analyse des ébauches d’erreur (voir figure 12).
I.6. Différentes méthodes d’analyse utilisées dans ce travail
I.6.1
Analyse des distributions de temps de réaction
L’analyse des distributions de TR est un outil particulièrement intéressant pour étudier les processus de traitement de l’information car il permet de décrire précisément la façon dont un facteur expérimental influence les performances cognitives. Les informations recueillies par l’intermédiaire de cette méthode sont beaucoup plus fines que de simples indices de tendance centrale tels que la moyenne ou la médiane. En effet, l’analyse des distributions de TR offre une vision globale de l’ensemble des TR recueillis au cours de différentes conditions expérimentales, elle permet de comprendre certains aspects ne pouvant être mis en évidence avec des indices de tendance centrale. La manipulation d’un facteur expérimental peut provoquer un changement de la forme de la distribution (figure 9A versus figure 9B) ou/et un décalage de la courbe (Figures adaptées de Spieler, Balota, & Faust, 2000 ; figure 9A versus figure 9C / figure 9D). Dans ces cas, la moyenne peut ne pas être affectée par ces changements et l’absence de résultat ne reflète pas une absence d’effet du facteur manipulé mais l’impossibilité de mettre en évidence cet effet en utilisant uniquement un indice de tendance centrale.
Figure 9 : Changements équivalents au niveau du TR moyen résultant soit d’un allongement de la forme de la distribution (B), soit d’un décalage de la distribution (C), et des effets compensés (D).
Deux techniques d’analyse sont généralement utilisées pour appréhender la forme des distributions de TR. La première technique consiste à ajuster une fonction de probabilité théorique aux données expérimentales individuelles collectées essai par essai (Luce, 1986). La deuxième technique d’analyse consiste à analyser les distributions moyennes en utilisant la technique de vincentisation’ élaborée par Vincent (1912). C’est la technique de vincentisation qui sera utilisée dans la première étude de ce manuscrit.
Cette technique proposée par Vincent (1912) puis développée par Ratcliff (1979) repose sur l’estimation d’une distribution moyenne représentative des distributions individuelles. Cette technique d’analyse présente, d’une part, l’avantage d’aborder la forme de la distribution sans présumer de la fonction de probabilité qui décrirait le mieux la distribution
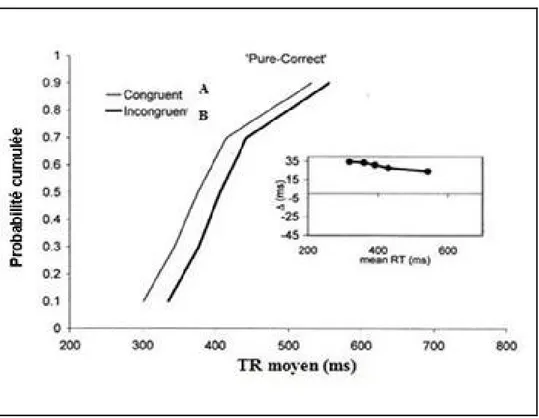
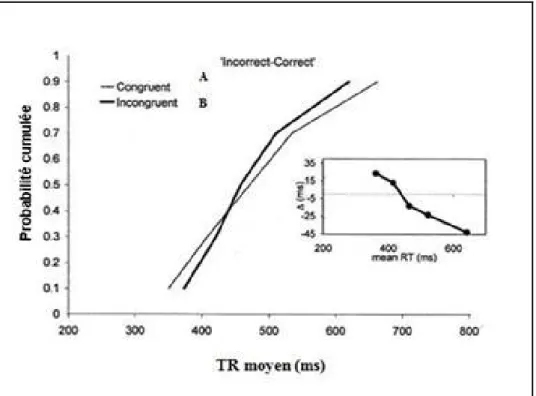
Ratcliff (1979) a montré que pour les distributions théoriques qui s’ajustent le mieux aux données de TR, la vincentisation permet d’obtenir une bonne estimation de la distribution de TR. Cette technique consiste à classer les TR pour chaque sujet et dans chaque condition expérimentale par ordre croissant (par exemple conditions A et B) (grandes figures 10,11), puis à décomposer la distribution de TR en classes de même effectif (quantiles). Le calcul du TR moyen de chaque quantile permet d’obtenir une distribution vincentisée individuelle pour chaque sujet, ainsi qu’une distribution vincentisée de groupe en moyennant les distributions individuelles quantile par quantile. On notera que, en général, il n’est pas possible d’obtenir une distribution de groupe simplement en mélangeant les distributions individuelles, car il n’est pas correct de moyenner simplement les ordonnées des distributions individuelles pour obtenir une distribution moyenne. Généralement, les distributions de TR sont graphiquement présentées sous la forme de fonction de densité de probabilité (probability density function) ou sous la forme de fonction de densité cumulée (cumulative density function). Dans le cadre de ce travail, l’ensemble des représentations graphiques sera présenté sous la forme de fonction de densité cumulée, c’est-à-dire la probabilité cumulée d’apparition d’une valeur de TR en fonction de la latence.
Pour chaque distribution vincentisée, l’indice delta plot dans ce travail était calculé (petites figures 10, 11). Cet indice rend compte à la fois de l’amplitude et du sens de l’effet du facteur manipulé pour chaque quantile. Exprimé en millisecondes, l’indice delta représente l’écart entre les distributions de TR de deux conditions expérimentales rapporté à la moyenne du TR moyen obtenu pour ces deux conditions (Delta = TR1 – TR2 en fonction de (TR1 + TR2)/2). Cet indice reflète la dynamique de l’effet du facteur manipulé. Par exemple, lorsque les distributions A et B sont identiques, le delta plot est égal à 0 pour chacun des points. Lorsque la différence est non nulle, mais constante pour chacun des points, cela signifie que les deux distributions se différencient seulement d’un terme constant. Les cas les plus intéressants sont ceux où les fonctions cumulées convergent ou divergent, voire se croisent comme dans l’exemple ci-dessus (figure 11) ; la courbe de différence est alors croissante (distributions A et B divergentes) ou décroissante (distributions A et B convergentes). On en infère alors que des processus différents sont à l’œuvre dans les TR courts et dans les TR longs. Pour mieux comprendre l’intérêt de ce type d’analyse, on présente un exemple issu d’une étude réalisée par Burle et ses collaborateurs en 2002. Dans une tâche de Simon, et à l’aide des enregistrements électromyographiques et l’analyse des distributions de TR, les auteurs ont montré, que la pente de delta plot (entre les deux distributions de TR dans les conditions congruente et incongruente) est plus négative dans les essais où il y a double