ACCLIMATATION HIVERNALE
CHEZ
TII\ PETIT ENDOTHERME
:LA
MÉSANGE
À
rÊrp
NOIRE (poECrLE
ATMCA?TLLUS)
THESE
pnÉseNrÉp
COMME
EXI GENCEPARTIELLE
DU DOCTORAT
ENBIOLOGIE EXTENSIONNÉ
DE L'UNTVERSTTÉDU
QUÉBECÀ
VONrnÉel
PAR
MAGALI
PETIT<Savoir pour prévoir, afin
depouvoir>
Auguste Comte
UNIVERSITÉ DU QUÉBEC À RIMOUSKI
Service de la bibliothèque
Avertissement
La diffusion de ce mémoire ou de cette thèse se fait dans le respect des droits de son auteur,
qui a signé le formulaire « Autorisation de reproduire et de diffuser un rapport, un
mémoire ou une thèse ». En signant ce formulaire, l’auteur concède à l’Université du
Québec à Rimouski une licence non exclusive d’utilisation et de publication de la totalité
ou d’une partie importante de son travail de recherche pour des fins pédagogiques et non
commerciales. Plus précisément, l’auteur autorise l’Université du Québec à Rimouski à
reproduire, diffuser, prêter, distribuer ou vendre des copies de son travail de recherche à des
fins non commerciales sur quelque support que ce soit, y compris l’Internet. Cette licence et
cette autorisation n’entraînent pas une renonciation de la part de l’auteur à ses droits
moraux ni à ses droits de propriété intellectuelle. Sauf entente contraire, l’auteur conserve
la liberté de diffuser et de commercialiser ou non ce travail dont il possède un exemplaire.
A
toutes les personnesqui
m'ont
accompagnée, aidée, encouragéetout au long de
cevoyage et sans qui
je
n'aurais probablement pas parcouru tout ce chemin, merci.Je remercie les membres de mon comité de thèse: Piero Calosi (membre inteme), François
Criscuolo (membre exteme) et Jonathan Verreault (président du
jury)
pour avoir pris le temps delire
cette thèse et pour leurs commentaires qui en ont amélioré la pertinence et la clarté.Merci
à toutes les personnesqui m'ont
permisd'accomplir
cetravail,
que cesoit
sur leterrain, au laboratoire
ou
au bureau.Merci Yves
Turcottepour m'avoir
apprisà
démailler.Merci André
Laberge etla
corporation dela FER
Macpèspour
l'accès aupavillon. Merci
Louise
Amyot,
PaulineDubé, Danie
Massé, Jonathan Coudéet Pierre Rioux pour
votre disponibilité et votre aide. MerciAlain
Caron, Quinn Fletcher et Pierre Legagneux pour votretemps et vos précieux conseils.
Plus particulièrement,
je
remercie mon directeur de recherche, François Yézina. François,merci
dem'avoir
fait
confiancepour participer au
succès deton
nouveau labo.Merci
dem'avoir
encouragéeà
développer mes propres intérêtsde
recherche,de m'avoir
si
bienencadrée, écoutée, guidée.
Merci
pour
ton
incroyable disponibilité,
ton
enthousiasme, tamotivation
et ta curiosité de tous les instantsqui
ont conduit à d'incessantes et intéressantesdiscussions au bureau, autour
d'un
café ou d'un bon repas,Merci
à mes amis quim'ont
aidée dans letravail
et surtout à avoir unevie
sociale malgréla
somme detravail
que requiert l'aboutissementd'un
doctorat.Un
grand merci à Agnès etClément avec qui
j'ai
commencé cette aventure.Merci
Agnès et Clément pour votre soutien,votre
aide, nos rires, notre amitié.Merci
Judith,Félix
et Rosaliepour m'avoir
accueillie etaccompagnée
depuis
le
début.
Merci
Fanny,
Laurence, Sabrina,Grisel, Karine,
Audrey,Lyette,
Emily,
Loli,
Jesi,
Nacho, Pablo, Adriano,
Amin et
tous
les
autres
pour
nosdiscussions, nos soirées, nos rires et tous les moments passés ensemble.
Merci
aussi à toutes ces personnes quej'ai
croisées au détour d'une cuisineI
ou d'un rassemblementl.
Merci
àma famille
(père, frère, sæur, belle-sæur, nièce, nevetrx, tante, cousine, cousin)qui, malgré
la
distance, a toujours su rester présente.Merci
Popapour
...
tout.Merci Lud
etDèd pour toutes ces années de soutien, d'encouragements et de rigolades. Je n'aurai pu rêver
meilleurs frère et sæur.
Merci
Mathilde, Dylan,Alex,
Théo et Tom, mes nièce et neveux qui,bien
qu'ayant grandi sansmoi,
savent toujours me montrerqu'ils n'oublient
pasleur
tata.TABLE
DES
MATIÈnns
INTRODUCTION GENERALE
I.1
Modifications de I'environnement et ajustements phénotypiques...1I.2
Budget énergétique et allocation desressources...,...
...2L3
L'hiver
sous les hautes latitudes, une saison contraignante...,.,...,.3L4
Lathermorégulation,...
...4L4.l
Lazone
dethermoneutralité...
...4I.4.2
Les réponses comportementales aux froidestempératures
...5L4.3
Les réponses physiologiques aux froidestempératures
...,...6L5
L'acclimatalion
hivernale chez les endothermes de petitetaille:
connaissances actuelles et objectifs de lathèse
...,.7I.5.1
Les patrons d'ajustements métaboliques intra-saisonniers ...8L5.2
Les causes exogènes des ajustementshivernaux...,...
...9I.5.3
Les mécanismes endogènes des ajustements hivernaux...,...11L5.4
Les conséquences des ajustements hivernaux sur la valeur séIective...13I.6
Méthodologiegénérale..
...14I.6.1
Modèle d'étude : la mésange à tête noire (Poecile atricapillus)...15I.6.2
Sited'étude
...17L6.3
Protocole de capture-marquage-recapfure(CMR)
...201.6.4
Protocole expérimental : le < feather clipping ))...
...22
CHAPITRE
I
La
flexibilité
intra-saisonnière
de
la
performance métabolique révèle
un
découplage
entre le
métabolisme basal
et
la
capacité
thermogénique
chez
lesoiseaux
231.1 Résumé...
.."'.'."26
1.2 Abstract...
...'....'.27
1.3 Introduction
...281
.4
Material andmethods
... .. ... . . . ' .... 3I
1.4.1
Capture andhandling
...311.4.2
Respirometry...
'..'.'....'32
1.4.3
Sexingindividuals...
'..."33
1.4.4
Statisticalanalvsis
.'...'.331.5 Results...
'...341.5.1
Temperature...
'.'.'....34
1.5.2 Inter-seasonal variation in body mass and metabolic performance...35
1
.5.3
Intra-seasonal variation of body mass and metabolic performance... 3 8 1.5.4RelationshipbetweenBMRandMsum
...'.'..'.'...'.41
I.6 Discussion
...411.6.1
Inter-seasonalvariationinmetabolicperformance.
...'....'.'..411.6.2
Intra-seasonal variation in metabolic performance and the uncoupling ofBMR
andMsum...
'..'..'...'...".'42
1.6.3 Intra-individual variation in winter metabolicperformance
.'...."..'..45
1.6.4 Is metabolic expansibility a meaningful
variable?
'...'.46
CHAPITRE
Normes
deréaction
en conditions
naturelles
: comment
lespetits
endothermes
ajustent-ils leur performance
métabolique aux
variations
météorologiques
?
472.1
Résumé...
...502.2
Abstract...
...'....512.3
Introduction
...'....522.4
Materials andmethods
...'."'55
2.4.1
Capture andhandling
...552.4.2
Respirometry,...
...562.4.3
Weatherdata
...572.4.4
Sexingindividuals
...,....582.4.5
Statisticalanalysis
...582.5 Results
...602.6 Discussion
...632.6.1
BMR.
...642.6.2
Msum
...65CHAPITRE
III
Les
manipulations
phénotypiques
confïrment
I'implication des
muscles
pectoraux et
de lohématocrite dans
la
capacité
thermogénique maximale
chezles
oiseaux
7l
3.1 Résumé...
...743.2 Abstract
...,...753.3 Introduction
...763.4
Materials andmethods
.,...783.4.1
Capture andhandling
...,...783.4.2
Respirometry...
...803.4.3
Experimentaltreatments
...813.4.4
Statisticalanalvsis
...813.5 Results
...,....,..843.5.1
Treatnenteffect...
...843.5.2
Efiect of pectoral muscle size and hæmatocrit on metabolism...863.6 Discussion
...89CHAPITRE
IV
93
Comment Ia
flexibilité
de
la
composition
corporelle reflète-t-elle
lesvariations
de
la
performance métabolique
chez
un petit
passereau
hivernant
sous
leshautes
latitudes
?
934.1 Résumé..,....
...,,964.2 Abstract...
...974.3 Introduction
...,.984.4
Materials andmethods
... 1004.4.1
Caplure and handling...
... 1004.4.2
Respirometry...
...1004.4.3
Organmass...
...L024.4.4
Statisticalanalvsis
... 1034.5 Resu1ts...
...1064.5.1
Seasonal variation in body mass and metabolicperformance
... 1064.5.2
Seasonal variation in lean dry organmass...
... 1094.6 Discussion
... I 134.6.1
Seasonal variationinbody
mass and metabolicperformance
...1144.6.2 Yaiarlon
in organ mass betweensexes...
... 1154.6.3
Yat'ratton in organ mass and relationships withBÀ/ffi.
..,... 1154.6.4
Yariation in organ mass and relationships withMsum..
...117CHAPITRE
V
tzl
L'augmentation hivernale de
la
capacité
intra-hivernale
desoiseaux
depetite
taille
thermogénique
favorise
la
survie
t2l
5.1 Résumé...
...1245.2 Abstract...
...I25
5.3 Introduction
...1265.4
Materials andmethods
...1.275.4.1
Capture andhandling
...1275.4.2
Respirometry...
...1285.5
5.6
5.4.4
Statisticalanalysis
...130Results
...1335.5.1 Within winter
survival...
...133Discussion
...141CONCLUSION
CÉNÉNAT,N
147
C.1 Ajustements de la performance métabolique au sein deI'hiver....
...I49
C.2
Facteurs météorologiques liés aux variations métaboliques...150C.3
Contribution de la composition corporelle aux ajustements métaboliques ...,...153C.4
Conséquences des ajustements métaboliques hivernaux sur la survie...,...156C.5
Syrthèse
...157C.5.1 Les ajustements du métabolisme de
base
...158C.5.2Lesajustementsdelacapacitéthermogénique...
...160C.6
Critiques del'étude
...162C.6.1 Contexte et
orisinalité...
...162C.6.2
Lim'ites...
...164C.7
Perspectives derecherche
...169C.7.1 Étude en conditions
expérimentales...
...170C.7.2
Étudeintégrée
...1'72 C.7.3 Étude à longterme....,...
...172^-, A. C.7.4 Etude en physiologie de la
conservation
...174ANNEXES
t77
LISTE
DES
TABLEAUX
Table
1.1Monthly minimal, mean and maximal
temperatures(oC)
recordedby
weatherstation
within
the study area.,...
...35Table2.I
Sample sizes presented relative to the number of capture per bird. ...56Table
2.2 Best model explaining
massand sex
independentMsum variations.
Residualminimal
absolutehumidity was extracted
from
a
polynomial
regression andcontrols for the effect
of
ambient temperature....
...62Table 3.1 Mean minimal
ambient temperature(+
s.e.m.) recordedhourly over the
studyperiod...
...79Table 4.1
Matrix
of
correlation showingall
comparisons between leandry
massesof
organgroups.
.,...105Table 5.1
Model
selection testingfor
the effectof
period and group (i.e. cohort and age) onthe return rate
of both
cohorts duringwinter.
(Worc":weight of
the model, Par :number of parameters, Dev.:
Deviance)...
...134 Table 5.2Model
selection testingfor
theeffect of
mean size-independentbody
mass(M,),
haematocrit(Hct),
residual basal metabolic rate(resBMR)
and residual summit metabolic rate (resMsum) on the return rate of both cohorts during winter. (Wor"": weight of the model, Par : number of parameters, Dev.: Deviance) ...135 Table 5.3Model
selection testingfor
theeffect of
mean size-independentbody
mass(M,),
haematocrit (Hct), basal metabolic rate(BMR)
and summit metabolic rate (Msum)on the return rate of both cohorts during winter. (Worc": weight of the model, Par :
number of parameters, Dev.:
Deviance).,,
... 137Table 5.4
Model
selection testing for the effectof
event and age on the retum rateof
cohortI
betweenyears. (QAICc: modified
AICc,
Wqarc":
weight
of
the model,
Par :number of parameters, QDev.:
deviance)
...139Table
5.5Model
selection testingfor the
effectof
mean size-independent body mass(M,),
haematocrit(Hct),
residual basal metabolic rate(resBMR)
and residual summit metabolic rate (resMsum)on the
return rateof
cohort 1
among years. (QAICc:modified AICc,
Wqarc" : weightof
the model, Par : number of parameters, QDev.:Deviance)...
...,.,...140Table
A.l
Inter-seasonalvariation
in
body
mass,basal metabolic
rate
(BMR),
summit metabolic rate(Msum)
and metabolicexpansibility (ME),
Data arc least squaremeans
controlling
for
year, season and sex (andtime
of
capturefor body
mass)with
birdID
as randomparameter.
...177TabIe A.2lntra-seasonal
variation
in
body
mass,basal metabolic
rate
(BMR),
summit metabolic rate(Msum)
and metabolicexpansibility (ME). Data
are least squareand
ME, with bird
ID
as random parameter.Body
mass analysis also includedtime of capture as
covariate...,
.'...'....'..'...177
Table
A.3 Full
model explaining
mass and sex independent basalmetabolic rate (BMR)
variation. ResidualBMR
were extractedfrom
anANCOVA controlling for
bodymass
and
sex. Residual
minimal
absolute
humidity
was
extracted
from
apolynomial regression and controls for the effect of ambient temperature... 178
Table
A.4 Full
model explaining mass and sex independent summit metabolic rate (Msum) variation. Residual Msum were extractedfrom
anANCOVA controlling for
bodymass
and
sex. Residual
minimal
absolute
humidity was
extracted
from
apolynomial regression and controls for the effect of ambient temperature...I79
Table
A.5
Basal metabolic rate (BÀrR) monthly variation. Least square means ofBMR
were calculatedfrom a
linearmixed
effect modelincluding body
mass and month asfixed parameters and bird
ID
as randomvariable.
.'..' 180Table
A.6
Summit metabolic rate(Msum)
monthly variation. Least square meansof
Msumwere
calculatedfrom
an
ANCOVA
including body
mass andmonth
as fixedparameters.
...'...'...'...'.
180Table
A.7
Black-capped chickadees' capture markage recapture(CMR)
data collected overthree periods during
winter
1for
cohort 1 and winter 2for
cohort 2,...181Table A.8 Black-capped chickadees' capture-markage-recapture
(CMR)
data collected duringFigure
I.1
Réponses compensatoires. Trois processus permettent auxindividus
des'ajusterà
leur
environnement(Env.):
1)
I'adaptation
génétiqueentre
générations,2)
laplasticité
phénotypiqueentre les
stadesjuvénile (Juv.)
et
adulte
(Ad.) et
3)
laflexibilité
phénotypique
chez
les
adultes
en
réponse
aux
variationsenvironnementales.
Différentes formes
représentent
différents
individus
etdifférentes couleurs représentent differents
phénofypes..
...2 FigureL2
Représentation du budget énergétique selon le modèle dubaril.
Après digestion etabsorption, I'assimilation des nutriments apporte l'énergie nécessaire à l'organisme
(le
baril)
pour répondre aux dépenses énergétiques (les robinets) associées au travail musculaire, à la croissance des tissus et à la production de chaleur. D'après Weiner(ree2).
...3Figure
L3
Relation entre la température ambiante et le métabolismed'un
animal endotherme.Dans la zone de thermoneutralité, un
individu
adulte, au repos et à jeun exprime sonmétabolisme de base (BIvfR). Hors de la
TNZ,
le métabolisme augmente. Quand latempérature
ambiante
devient
trop
froide, I'animal atteint
sa
capacité thermogénique maximale mesurablevia
son métabolisme maximal, le Msum.A
ce stade,l'individu
entre en hypothermie et son métabolisme diminue.Modifié
d'après(lVillmer
eta1.,2005).
...5Figure
I.4
Echangesde
chaleur entreun
animal
et
son
environnement.Les
transferts dechaleur
se
font par
conduction, convection,
évapotranspirationet
radiation.@
Masali
Pe1it...,
...6Figure L5 Mésange à tête noire en hiver, forêt Macpès. O François Yéztrn.
Figure
I.6 Distribution
dela
mésange à tête noire,La
répartition de la mésange à tête noires'étend entre 37oN (Colorado)
et
67oN (Alaska) (gris).
Au
site d'étude
(flèche rouge), àRimouski
(Québec, Canada),les mésanges sont près de lalimite
nord-est deleur
aire dedistribution.
(Source Environnement Canada : http://www.ec.cg.ca). ...,...l6
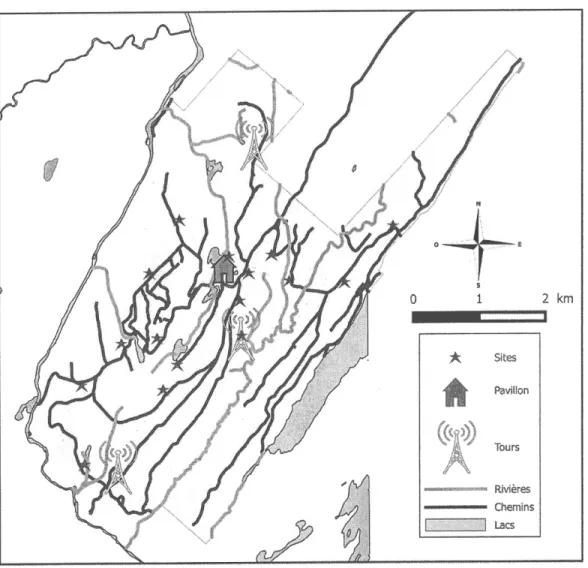
Figure I.7 Forêt d'enseignement et de recherche (FER) Macpès. Sont représentées les trois toursmétéorologiques
(aune), le pavillon
de recherche etles
16 sites de capture (étoiles rouges) utilisés pour cetteétude...
,... 18Figure
L9
Sites de capture. Les mésanges, attirées par les graines de toumesol, entrent dans latrappe et y sont piégées par la porte coulissante. O Magali Petit. ... ...20 Figure
LlO
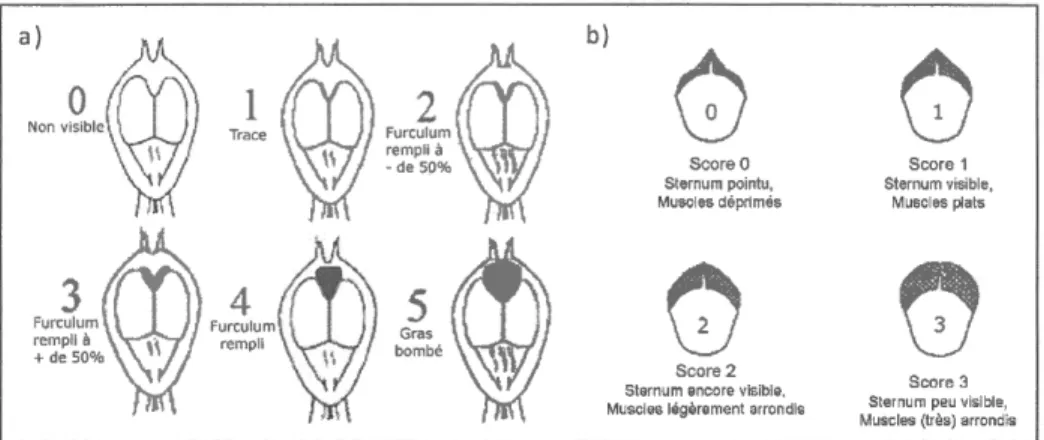
Mesures morphométriques. a)Indice
de grasfurculaire
sur une échellede0 à5
(d'aprèsGosler,
1996).b) Taille
des musclespectorarx
sur une échelle de0 à
3Figure I.1
I
Système de respirométrieutilisé
pour mesurer le métabolisme des mésanges à laFER Macpès. @ Magali
Petit...
'.'.'...21Figure L12
Aile
et queue dont les primaires (P1, P3,P5,P7
et P9) et les rectrices (R2 et R4)furent coupées le long du rachis. @ Magali
Petit'."...'.
..."..'."...'.22
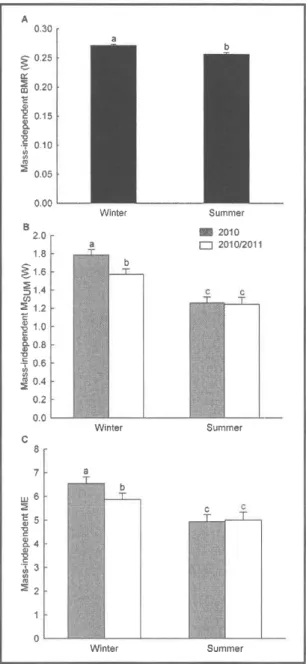
Figure 1 .1 lnter-seasonal variationof
mass-independentBMR,
Msum andME.
Data are least square meansfor BMR (A),
Msum (B) andME
(C) controlling for year, season, sexand body
masswith bird
ID
as random
parameter.Different letters
indicatesignificant difference between
seasons.
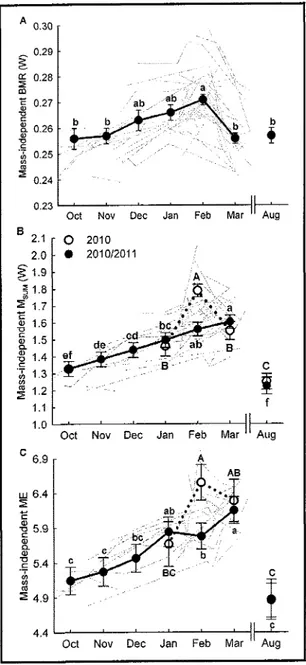
...'...'..'.'...'...37
Figure
1.2 Intra-seasonal changesin body
mass. Data are least square meansof
body masscontrolling
for
year, month, timeof
capture, sexwith
birdID
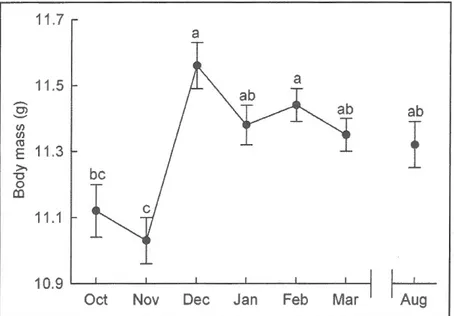
as random parameter.Different letters represent significant difference between months...,,...38
Figure
1.3Monthly variation at the population
andindividual levels
in
mass-independentBMR,
Msum andME.
Population data (blackline
and dots) are least square meansfor BMR (A), Msum (B)
andME (C) controlling for
year, month, sex, body massand
bird
ID
as random
parameter.In
B
and
C
-
dotted
line,
open dots
anduppercase:
l"
year;solid line, black
dots and lowercase: 2no year.Different
letters representsignificant
difference between monthswithin a
year. Predicted valuescalculated
from LMEs
(seetext
for
details) are used
to
visualize data
from individuals captured more than once (grey lines)....'...
...'...40Figure
1.4 Visual representationof
the monthly winter incrementin
mass-independentBMR
(A)
and Msum (B) relative to summer values. Data measuredin
August are used asthe summer phenotype reference
point (August:0To,
black star, dashed line)while
maximal levels ofBMR
and Msum recorded inwinter
are considered representativeof the "peak"
winter
phenotype (February:
100%).Monthly
changesin
metabolicperformance (values above
lines)
are presentedin
percentof
total
inter-seasonaldifference.
Dotted lines
representBMR
variations below the
summer referencepoint.
...,,.
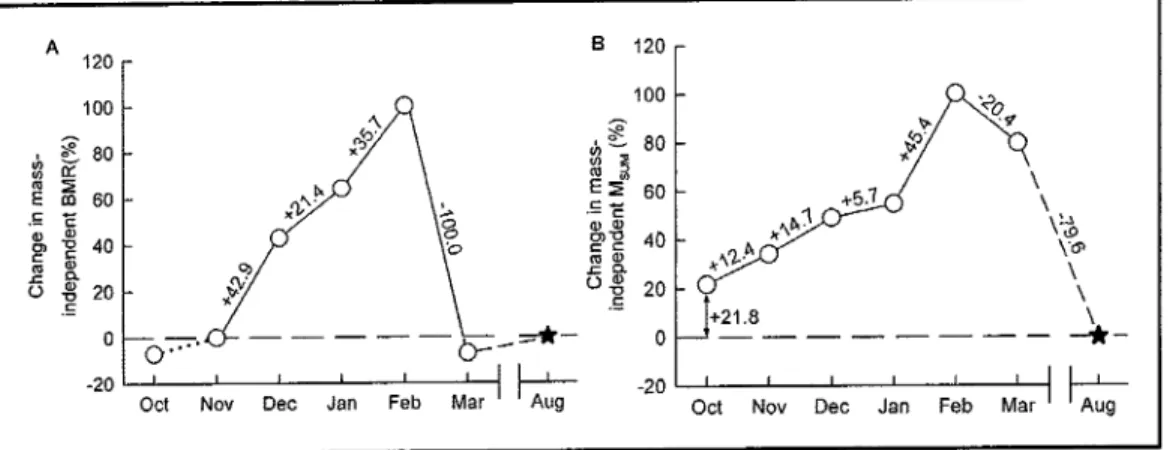
...".'..."""44
Figure 2.1 Reaction nonns accordingto
three scenarios.In
(a)individuals
express differentelevations and slopes,
in (b)
individuals
expressdifferent
elevationsbut
similarslopes and
in
(c) individuals express the same elevations and slopes, which reflect thepopulation reaction
nonn...,...
.'...'.53Figure 2.2 Relationship between mass and sex independent
BMR
and the natural rangeof
minimal
ambient temperature. ResidualBMR were
extractedfrom
anANCOVA
confolling
for body mass and sex. Population-average reaction norm is representedby
theblack line while
greylines
represent reactionnorns
of
individuals
caughtmore than
five
times....
...'..61Figure 2.3
Massand
sex independentMsum variation over the natural
rangeof
weatherconditions.
Data
are residual
Msum
presentedagainst
(a)
minimal
ambient temperature (min Ta) and (b) residual minimal absolute humidity.Black
dotsin
(b) represent data collected atmin
Ta < OoC, white dots at O'C<min Ta<16oC and grey dots atmin
Ta>16oC (seetext for
details). For both panels,black
lines are curvesfitted
by
loess(smoothness:0.85,
degree:
2).
ResidualMsum were
extractedhumidity
in
(a) orminimal
temperaturein
(b). Residualminimal
absolutehumidity
controlsfor
the effectof
ambient temperature and was extractedfrom
a polynomialregression.
Coefficients
of
determination
are
presented
for
independent relationships, see textfor
completemodel.
...,...63Figure 2.4 Monthly variations
in
minimal
temperature. DaTa are extremelow,
mean andextreme high values per month for
minimal
ambient temperature...65Figure 3.1 Capfures and recaptures
of
Black-capped chickadees. Eachindividual
capture isrepresented
by
a circle. Grey circles represent individuals that were not recaptured.Circlesjoined
by a line represent captures and recapturesfor
given individuals. ...78Figure 3 .2 Y ariation
in
body mass over the experimental period. DaIa are least square meansofbody
massfor
control and clipped Black-capped chickadees before and after thetreatment.
Least
square meanswere
extractedfrom
separatedLME
models pergroup
controlling for
period, body size and timeof
capture,with bird ID
as randomparameter,
Asterisk
indicatesthe significant
increasein
body
masswithin
thecontrol group between periods (:n:*{< p <
0.001).
...85Figure 3.3
Variation in
muscle score over the experimental period.Data
are median muscle scoresfor control
and clipped Black-capped chickadees measured before and after the treatment (as these are median values, there are no effor bars)...85Figure 3.4
Changesin
thermogenic capacity and haematocritin
Black-capped chickadeesover the experimental period. Data are least square means
of (A)
summit metabolicrate (Msum) and (B) haematocrit before and after the treatment. Least square means
were
extracted
from an
LME
model controlling
for
group, period and
theinteraction tetm group
x
period,with
birdID
as random parameter. Body mass wasalso included in the model as a covariate for
Msum.
...,...86 Figure 3.5 Relationship betweenMsum
and haematocritin
Black-capped chickadees. Dataare predicted Msum values plotted
against haematocrit. Predictedvalues
werecomputed
from a
LME
model controlling
for
group,period, the
interaction termgroup
x
period, muscle
scoreand body
massas
covariates,with bird
ID
as arandom
parameter.
...87Figure
3.6
Relationship between
Msum and
pectoral muscle score
in
Black-cappedchickadees.
Msum
data
areleast
square means extractedfrom
an
LME
modelcontrolling for
group, period, the interaction termgroup x
period,body
mass andhaematocrit as
a
covariates,with bird
ID
as random parameter.Different
lettersindicate significant differences (p <
0.05).
...88 Figure4.1
Seasonal variationin body
composition throughout the year. Data are least squaremean
of
leandry
massof
A)
liver,
B)
kidneys,C)
intestines,D)
gizzafi,
E)
size-independent
leg
muscles,
F)size-independent
pectoralis
muscles,
G) supracorqcalder,c muscles,H)
size-independent skeletal muscles,I)
heart, J) lungs,K)
brain and
L)
size-independent skin. Least square means were obtainedfrom
aGLM
including period, sex, period*sex as independent variables and body size as covariate.Figure
4.2
Seasonal variationin
body mass and lean dry body mass throughout the year' Data are least square mean obtainedfrom a GLM
including period, sex, relativetime
of
capture and period*sex as independent variables aswell
asbody
size as covariate.Different letters represent significant difference between
periods.
...107Figure 4.3
Metabolic variations throughout the year. Least square meanof
A)
BMR
and B)Msum were obtained
from
a
GLM
including period,
sex
and
periodxsex asindependent
variables. Different letters
representsignificant difference
befween periôds.LDM
: lean drym4ss...
... 109Figure 4.4 Seasonal variation in organ groups throughout the year. Least square mean of
A)
size-independent digestive organ,
B)
cardiopulmonary organ, C) size-independent muscle,D) brain and E) size-independent skin lean dry masses as well as of F) fat and G) size-independent water content. Least square means were obtained from a
GLM
includingperiôd, sex, period*sex as independent variables (and size as covariate
for
digestiveorgans, muscles,
skin
and water
content).Different letters
represent significant diiference betweenperiods.
...'.,'..'."""'
111Figure 4.5 Contribution of body components to metabolic performance throughout the year. Bars
represent the 12 adjusted
from
the best model explaining size-independentBMR
orwhole
Msum for
each period (see methods). Organs contributingto
the models aregiven in bars. Organs between ( ) were retained in the models but were not significant. Summer is not represented since
BMR
and Msum were not foundto
relate to bodycomponents during this
period.
.'.'... 113Figure 5.1 Diagram representing both the apparent survival ((D) and the encounter probabilities
(p) within winter. Apparent survival and encounter probability are constant. ... 133
Figure 5.2 Relationship between
winter
apparentsurvival
and a)winter
uncorrected Msum,with
survival controlledfor
the effect of hæmatocrit and b)winter
raw Msum,with
survival controlled for the effect ofBMR
(see textfor
details),
...136Figure
5.3
Diagram
representingboth the
apparent survival
(@) and
the
encounterprobabilities
(p)
among years.Apparent survival was
constantwhile
encounterprobabilities decreased between the end
of
awinter
and the beginningof
the nextone...
....138Figure C.1 Condition, qualité et valeur sélective. L'ensemble des paramètres physiologiques
détermine
la
condition physiologique
mais
seuls certains
de
ces
paramètresLISTE
DES
ABBREVIATIONS
BMR
CMR
DEE FERGLM
HCT
LCT
LME
LRT
MgMMR
Msum T5TNZ
Basal metabolic rate
Capture marquage recapture
Daily energetic expenditwe
Forêt d'enseignement et de recherche
General linear model
Hématocrite
Lower
critical
temperatureLinear mixed effect
Likelihood ratio test
Body mass
Maximal metabolic rate
Summit metabolic rate
Body temperature
nÉsuruÉ
Les
changementsglobaux
sont
associésà
une
augmentationde
la
stochasticité
del'environnement pouvant affecter
la
phénologie,la
démographie etla
variabilité génétique des espèces.La flexibilité
phénotypique permet aux organismes d'ajuster leur phénotypeà
courtterme,
ce qui
dewait
leur
conferer
une
certaine
capacitéà
affronter
les
fluctuations environnementales. Dans ce contexte,il
est donc important d'étudier\a
capacité de réponse desindividus aux
variationsde leur
environnementafin de
comprendreI'effet
des changementsenvirorurementaux sur
la
dynamique des populations. Cependant, nous manquons d'étude enconditions naturelles sur
la
capacité d'ajustements à court terme de paramètres physiologiqueschez
les
endothermes.Avec
cette thèsede
doctorat,nous
avonsutilisé une
population de mésangesà
tête noire (Poecileatricapillus)
résidant au Québecpour
étudier les ajustementshivernaux du métabolisme de base
(BI\R)
et dela
capacité thermogénique maximale (Msum) enconditions naturelles. Plus particulièrement,
les
objectifsde
cette thèse étaientde
methe enévidence
l)
les patrons d'ajustements duBMR
et du Msum (chapitre1),2)
la relation entre lesconditions
météorologiqueset la
performance métabolique(chapitre
2), 3)
l'effet de
la composition corporelle sur leBMR
et le Msum (chapitres 3 et 4) et 4) le lien entre le phénotypehivemal et la valeur sélective (chapitre 5).
Avec le chapitre 1, nous avons obseryé, pour la première fois en milieu naturel, qu'un oiseau de
petite
taille
est capable d'ajuster son métabolismehivemal à
court terme.Nos
résultats ontmontré
une
augmentationhivemale
du
BMR
(6%)
et du
Msum (34%)
et
ont
révélé undécouplage
temporel
enfe
ces deux paramèîes.
Alors
que
I'augmentationdu BMR
necommençait qu'en novembre pour revenir à un niveau estival dès mars, le Msum commençait à augmenter dès la
fin
de l'été et restait élevé en mars. Ce décalage temporel entreBMR
et Msumsuggère que les deux paramètres métaboliques répondent à des contraintes hivemales différentes.
Avec
le
chapitre 2, nous avons décrit, pour la première fois chez une espèce aviaire en liberté,les normes de réaction
du BMR et du Msum
en fonctiondu
gradient nafurel des conditions météorologiques. Les résultats ont révélé que le métabolisme de base différait enûe les individuset
était
faiblementet
négativementrelié
à la
tempérahreminimale. Quant
à la
capacitéthermogénique,
nos
donnéesont montré que les
mésanges ajustaientleur Msum
avec latempérature minimale selon une courbe sigmoïde, augmentant linéairement entre 24"C et -10oC,
et avec
l'humidité
absolue selon une courbe enU.
Ces résultats impliquent que les coûts de maintenance seraient surtout individu-dépendants et ne seraient que faiblement influencés par lesconditions
météorologiquesalors
que
Ia
capacité
thermogéniqueserait,
au
contraire,principalement
liée aux
conditions
météorologiques, notammentla
tempérafure.Avec
lechapitre 3, en manipulant la taille des muscles par un protocole de réduction de surface alaire,
nous avons démontré, pour
la
premièrefois
enmilieu
naturel,le lien
de cause à effet enffe lataille
des muscles pectorauxet la
capacité thermogénique maximale.Nos
résultatsont
aussimontré que le Msum était positivement conélé à I'hématocrite. Cette étude démontre que chez
un
oiseaude petite taille en
conditions naturelles,la
thermogénèsepar
frissonnement est supportée par les muscles pectoraux et probablement parla
capacité de transport de l'oxygène.Le
chapitre
4
nousa
permis d'étudierle
lien
entrela
variationde
masse des tissuset
les ajustements duBMR
et du Msum au cours de I'année. Nos résultats ont montré quLe 64%o de lavariation annuelle du Msum était expliquée par les changements de masse des muscles et des
organes cardio-pulmonaires
alors
que 35o dela
variabilité annuelledu BMR
étaitliée à
la masse des muscles squelettiques et des organes excréteurs (foie+reins). Finalement, les résultatsùt
chapitre 5 ont montré que, chez un petit passereau résident, la survie intra-hivemale était liée au Msum suivant une courbe sigmoide alors quela
survie à long terme était indépendante duphénotype hivemal. Cette étude
met
en évidence,pour la
premièrefois
chez une populationnaturelle d'oiseaux, une relation positive
enfe
le métabolisme hivernal et la valeur sélective.Mots-clefs
:
flexibilité
phénotypique; endotherme;
acclimatationhivernale;
performance métabolique ; oiseau ; métabolisme de base;
capacité thermogénique maximale.ABSTRACT
Global
changes are associatedwith
an
increasein
weather stochasticity,which could
affect phenology, demography and geneticvariability of
species. Phenotypicflexibility,
which allowsindividuals
to
adjusttheir
physiologyrapidly
to
variationsof
their
habitat should provide acertain capacity
to buffer
environmental fluctuations.In
this context,it
is
therefore crucial to study response capacity of individuals to surrounding variationsin
order to understand the effectof
environmental changes on population dynamics. However, as far as we know, there is a lackof
study
on
capacityfor
short-term adjustmentsof
physiological parametersin
free-livingendotherms.
With
this
thesis,we
useda
population
of
Black-capped chickadees (Poecileatricapillus)
living in
Quebecto
studywinter
adjustmentsof
basal metabolic rate(BMR)
andmaximal thermogenic capacity (lvtsum)
in
natural conditions.More
specifically, the objectivesof
this thesis wereto highlight
1) the intra-seasonal pattemof BMR
andMsum
adjustments(chapter
1),2)
the relationship between weather conditions and metabolic performance (chapter2), 3)
the effectof
body composition onBMR
and Msum (chapters3
and4)
and4)
thelink
between winter phenotype and fitress (chapter
fl.
With
chapter 1, we showed for the first time in natural conditions that this small avian species isable to adjust its winter metabolism on a short time scale. Results revealed a winter increase in
BMR
(6%) and Msum (34%) and a temporal mismatch between these parameters. WhileBMR
beganits
seasonal increasein
November and came backat a
summer-like value as soon asMarch, Msum began to increase at the end of summer and stayed high in March. This difference
in the timing of variation in winter
BMR
and Msum suggests that the two parameters respond todifferent winter constraints.
With
chapter 2, we described, for the first timein
a freeJiving birdspecies, reaction norms
of
BMR
andMsum
across the natural rangeof
weather variations. Results showed thatBMR
varied between individuals and was weakly and negatively related tominimal
temperature. Data revealed that Black-capped chickadees adjustedtheir Msum with
minimal temperaturefollowing a
sigmoid curve, increasing linearly between 24oC and -10oC, andwith
absolute humidityfollowing
a U-shape relationship. These results suggest that weatherconditions have
minimal
effectson
maintenance costs,which would mainly be
individual-dependent.
In
contrast, thermogenic capacitywould mainly
respondto
weather conditions, especially temperature.With
chapter 3, using a feather clipping protocol to manipulate musclesize,
we
demonstrated,for
the first time
in
natural conditions,the
causaleffect
of
pectoral muscle size on maximal thermogenic capacity. Results also highlighted that Msum is positivelycorrelated
with
hæmatocrit. These findings therefore demonstrate thatin
a small-bodiedfree-living
bird, shivering thermogenesis is supportedby
pectoral muscle size and probably also by oxygen carryingcapacity.In
Chapter y',we
studied the relationship between changesin
bodycomposition and metabolic performance
over the
year. Results showed that 640Âof
Msumvariations throughout the year were explained
by
changes in body muscles and cardiopulmonary organswhile 35%
of
the
annualBMR variability
was relatedto
variationsin
massof
body muscles and excretory organs (liver+kidney).Finally,
resultsof
chapter5
showed that,in
asmall
resident passerine,within winter survival was
relatedto
Msum following
a
sigmoidrevealed,
for
the
first
time
in
free-living birds,
a
positive
relationship between winterthermogenic capacity and fiûress.
Keywords:phenotypic
flexibility;
endotherm;winter
acclimatization; metabolic performance;INTRODIJCTION
GE.NtrRALE,
I.l
Modilications
de
I'environnement
et a.justements phénotypiques
Outre
un
réchaufïèmentglobal
et
Lule élévationdu
niveau des océans.les
changetlentsclimatiques s'accompâgneut d'évéuements météorologiques extrêmes
(e.9.
sécheresse. pluie velglaçaute) de ph"rs en grh,rs fréqueuts (fPCC,20l3).
Les populatious aniurales dont les indiviclusexprimenl
une
capacité
cle réponse phénolypique
(i.e.
physiologique,
comportemenLale)insufl'isante pour
fàile
fàce à I'augmentation cle stochasticité cle l'envirolmernellt pounaient ainsiêtre défàvorisées ou aurenées à disparaîtle (Canale
&
Heruy,2010;
Forcada et a1..2008). Pourfàile
fàce arLx varial-ions envirorlrementales. les organisrres pellvent s'a-juster àtlois
échelles detenrps (Seebaclrer
&
Frarrklin. 20il2) (tigurc
T.l).
À
l'éclrelle
cles gérrératiorrs. I'adaptatiorr génétique pernlet aux populations de s'adapter aux charrgements de leur lrabitat (HotTiranrr&
Sgrô,20ll).
Au
cours de la croissance. la plasticité phénotypiqLre penllet à un inclivicil-r.iuvénile cle dér,elopperur
phénotype adulte acléquatà
sonmilieu
(Traviset
al..
1999).Dutant la
vieadr-rlte, la
flexibilité
phénotypique llermet à un indiviclu d'a-juster son phénotype de f'açon rapide et réversible aux variations de son euvironnement (Piersma&
Drent, 2003). Par conséquent, lemaintien cles populations clans leur airc cle répartition acnrelle dépend cle leur capacité à r'epondre
nou
seulementanx
changeurents climatiquesà long
terme,vla
des mécanismes d'adaptatiolt.r.nais aussi aux changeurents à cor.rrt tenne. vra
la flexibilité
phénotypiqr-re. Les individr-rs ayallt une capacité cl'ajustement insufllsante pour suivre la vil.esse des 1'lLrctuations euvironnemenlalesn'auraieut cl'autres
choix
que cle se cléplacer vers d'autres habitatspour
survivre (Canale&
Herny.2010;
Reeclet a1.,2010; Van De Pol et a1.,2010).
Il
esL donc essentield'étudier
lacapacité
cl'ajusterrrentcles
orgallisrnes
pour
comprendre
I'efTet
de la
variatiot't
deI'enviLoruremellt sur leur valeur sélective, sur la clynaurique des populations et sltr le devenir des esnèces.
,
Juv.1
Ad.1
Ad.1l-re
Env. A Env. B Juv.2ô
\
I I I Ie
Env.c
Env. DAd.2
Ad.2Figure
I.1
Réponses colnpellsatoires.Trois
processLrs perfiIettentalrx
indivrdris
de
s'ajusterà
leur
enl,ironnernent
(Env.): l)
I'aclaptation génétique
entre
générations.
2) la
plasticité phénotypique entre les stadesjuvénile
(Juv.) et actulte(Ad.)
et3)
lailexibilité
phénotypique chez les adultes en réponse au-r variationseuvironrreureutales. Ditïér'errtes
fbrnres
repr'ésentent differerrtsi ndiv idus et di tTérerrtes coulerLrs leprésenter.r t ditTéren ts ph énotypes.
1.2
Budget énergétique et
allocation
des ressources.
La
r,aleur sélectived'urr
inctividn dépend principalement cle sa capacitéà
acquérir chaqltejour
s-rtïisamllletlt
d'énergiepour
assurel ses acti\.ités(e.g.
therrnorégulation, reproduction)(Brodin.2007) (figure
1.2).Les
ressources énergétiqr-resprovienueut
de
I'alimentatiott
etdépendent de
la
qualité dela
noun'iture (e.g. contenu lipicliqure), de la capacité deI'animal
àacquérir
et
ingérer cette uourriftre aiusi
quede sa
capacitéà en assimiler l'énergie.
Lesdépenses énelgériques quotidiennes. quant
à
elles, (DEE.pour
<daily
energy expenditure >)englobent ['enseurble
clesactivités
réaliséespar
Lulincliviclu (e.g. locomotion.
digestion. reproduction, thermorégulation) (Piersma. 2002; Rickle fs e tal..
1 996).pour répondre aux dépenses énergétiques (les robinets) associées au
travail
musculaire, à lacroissance des tissus et à la production de chaleur. D'après Weiner (1992).
La
somme des apports et des dépenses énergétiques constitue le budget énergétique. Plusun budget énergétique est
positif
plus lesindividus
peuvent être performantspuisqu'ils
ont plus d'énergie à allouer à leurs activités. Par contre, dans un environnement contraignant, oùles besoins énergétiques sont accrus et les ressources limitées, des individus peuvent avoir un budget énergétique négatif et donc trop peu d'énergie disponible pour maintenir toutes leurs
activités (e.g. défenses immunitaires), ce
qui
à long terme pourrait compromettre leur valeur sélective.Selon
le
principe d'allocation
des ressources(Daan
et
al.,
1990; Weathers&
Sullivan,
1993;Weiner,
1992), dansune situation
énergétiquementdifficile,
les
individusutilisent
des ajustements phénotypiquesafin
dediminuer
leurs dépensesetlou
d'augmenter leurs apports et rééquilibrer leur budget énergétique (Masman etal.,
1986).I.3
L'hiver
sous leshautes
latitudes, une saison
contraignante
L'hiver
en régions tempérées froides est considéré comme undéfi
énergétique majeur pour les espèces homéothermes résidentes (Cooper, 2000; Weathers&
Sullivan, 1993). Les froides températures obligentles
espècesqui
restent activesà
augmenterles
dépenses énergétiquesdestinées à leur thermorégulation afin de maintenir leur température corporelle stable (Cooper
&
Swanson, 1994; Liknes
&
Swanson, 1996) alors quela
courte durée dujour
etla
couverhreneigeuse entravent
leur
quête alimentaireet
donc potentiellement leurs apports énergétiquesMcNamara et
al.,
7990; Swanson, 2010). Pour surviwe à ces conditions hivemales exigeantes,les espèces résidentes ajustent
leur
phénotype,c'est
I'acclimatation hivemale. Ce processus, composé d'rur ensemble d'ajustements physiologiques et comportementaux, permet d'augmenterles
dépenses énergétiques destinéesà la
thermorégulationen dépit
d'un
accèslimité
à
la nourriture (Cooper&
Swanson, 1994;Swanson,l99la;
Swanson&
Thomas, 2007).t.4
La thermorégulation
1.4,1
Lazone
dethermoneutralité
La
zonede
thermoneutralité(TNZ
pour (thermoneutral zone>),
délimitéepar
les températures critiques inférieure(LCT
pour << lower critical temperature >) et supérieure (UCTpour
(
upper critcal temperature >) (figure I.3), correspond à la plage de températures ambiantesdans laquelle
un
animal
endothermen'a
besoinde foumir
aucuntravail
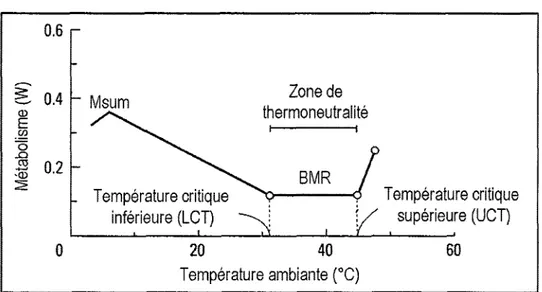
métabolique pourmaintenir sa température corporelle constante Ç1ill
et
al.,
2004).C'est
dans cette plage detempératures que
le
métabolismede
base(BMR
pour
<<basalmetabolic
rate >>)peut
êtremesuré.
Le BMR
se mesure chezun individu
adulte, àjeun,
à thermoneutralité et durant laphase
inactive
du
cycle circadien.
Le
métabolismede
base
représentedonc
l'énergieminimale
nécessaireà un individu
endothermepour
assurer sa maintenance physiologique(e.g,
battements cardiaques,
filtration
rénale) sans prendre
en
compte
les
dépensesénergétiques associées
au
développement,à la
digestion,
à la
thermorégulationet
auxactivités j oumalières (D aan et
al.,
1990; McKechnie, 2008).Un
animal exposé à une température hors de saTNZ
doit par contre ajuster sa physiologieetlou son
comportement(Hayes
&
Garland, 1995;
Seebacher,2009) pour maintenir
satempérature corporelle stable. Ces ajustements engendrent
une
augmentationdes
dépenses énergétiques et ainsi une augmentation du métabolisme au-dessus duBMR
(figureI.3)
(Bush el.(/)
6
-o
s
s.2 Température critique supôrieure (UCT)20
40 Température ambiante ("C) Température critique inférieure (LCT)Figure
I.3
Relation
entrela
tempérafure ambianteet le
métabolismed'un
animalendotherme. Dans la zone de thermoneutralité, un
individu
adulte, au repos et àjeun
exprime son métabolisme
de
base
(BMR). Hors
de
la
TNZ,
le
métabolisme augmente. Quandla
température ambiante devienttrop froide, l'animal
atteint sacapacité thermogénique
maximale
mesurablevia
son
métabolismemaximal,
leMsum.
A
ce
stade,I'individu
entreen
hypothermieet
son métabolisme diminue.Modifré
d'après(Willmer
et aL.,2005).I.4.2
Les réponses comportementales auxfroides températures
Les échanges de chaleur entre un animal et son environnement se
font
a) par conduction,directement entre
derx
matériaux en contact(Willmer
et aL.,2005); b) par convection viatn
fluide
(air
ou
eau)(Hlll
et
al.,
2004);
c)
par radiation
thermiquevra l'émission
d'ondes infrarouges(Hill
et aL.,2004) et d) par évapotranspiration(Randallet a|.,2002).
Exposés à defroides
températures,les
animaux perdent
de
la
chaleur
par
évapotranspiration, parconduction
lorsqu'ils
sont en contact avec une surfacefroide,
par convectionlorsqu'ils
sont exposés auvent et par radiation,
surtoutla
nuit
(figure I.4). Pour minimiser
les pertes etmaximiser les gains de
chaleur,les
animaux peuventajuster leur
comportementafin
demoduler les échanges de chaleur avec I'environnement. Par exemple, ils peuvent sélectionner
des micro-habitats à
I'abri
du vent pour réduire les pertes de chaleur par convection (Marsh&
Dawson, 1989; Scholander etal.,l950b;
Wolf
er aL.,1996) ou ils peuvent s'exposer au soleilpour
augmenterles
apportsde
chaleurpar radiation
(Doucette&
Geiser,2008;
Geiser&
Drury,2003),
environnement.
Les
transfertsconvection, évapotranspiration et radiation. O Magali Petit.
I.4.3
Les réponses physiologiques auxfroides
températuresLes
animaux peuvent
aussi
limiter les
pertes
de
chaleur
par
des
ajustementsphysiologiques. Par exemple, en augmentant leur isolation thermique (e.g. par piloérection) (Marsh
&
Dawson,
1989; Scholanderet
al.,
1950b; Swanson,l99la)
et
en augmentant lavasoconstriction des extrémités (Marsh
&
Dawson, 1989). De plus, toute activité produisant dela
chaleur,la
chaleur générée lors dela
locomotionou
dela
digestion participe aussi aumaintien
de
la
températurecorporelle chez
les
organismes endothermes,c'est
de
lathermorégulation par substitution (Cooper
&
Sonsthagen, 2007)'Lorsque les ajustements comportementaux et la thermorégulation par substitution ne suffisent plus à compenser les pertes de chaleur, les endothermes utilisent principalement la thermogénèse
par
frissonnementpour produire
activementde
la
chaleur
et
maintenir leur
températurecorporelle.
La
thermogénèsepar
frissonnement représentela
première sourcede
chaleurendogène chez tous les vertébrés endothermes (Hohtola, 2002). En réponse au froid, le système
nerveux
somatique déclenchedes
contractions
asynchroneset
soutenuesdes
muscles squelettiques antagonistes. Puisque les contractions sont aléatoires, le frissonnement permet auxchaleur jusqu'à atteindre sa capacité thermogénique maximale (Msum, pour (( summit metabolic
rate >>, figure I.3). Ensuite, si la température ambiante continue de diminuer, la thermogénèse ne
suffit plus à compenser les pertes de chaleur et
I'individu
entre en hypothermie, ce qui se traduit par une diminution du métabolisme. De cefait,
le Msum correspond au métabolisme maximal d'un individu exposé aufroid
(Dawson&
Smith, 1986; Marsh&
Dawson, 1989). Le Msum estmesuré en conditions expérimentales et est généralement atteint à des températures plus froides que celle rencontrées en nature. Cependant, parce que
le
Msum est corrélé àla
tolérance desindividus au
froid
(Swanson, 1990a; Swanson, 2001; Swanson&
Liknes,2006),
c'est un indicede
la
capacité thermogénique courammentutilisé
dansles
étudessur
la
thermorégulationMcKechnie &
Swanson,2010; Swanson,2010; Swanson et a1.,2012).L5
L'acclimatation hivernale
chez
les
endothermes
de
petite
taille:
connaissances actuelles et
objectifs
de la thèse
L'acclimatation hivernale chez
les
endothermesde
petite
taille
est
généralement caractérisée, relativementà l'été, par un
accroissement dela taille
des organes(Liu
&
Li,
2006; Zheng eta1.,2008),
par une augmentation du métabolisme de baseet
dela
capacité thermogénique (Cooper&
Swanson, 1994; Swanson&Yézina,2015;Zhao
et a/., 2010) ainsique
par
une
haussede I'activité
des
hormoneset
des
enzymesmises en
jeu
dans
lecatabolisme
des lipides, des glucides
et
des protéines(Dawson
et
al.,
1992; Liknes
&
Swanson, 2011a).
Ainsi,
pour affronter les températures hivemales, ces animaux ajustent leur physiologie (e.g. activité enzymatique) (Liknes&
Swanson,20lIa)
et leur morphologie (e.9.taille
des organes) (Liknes&
Swanson, 2011b).Bien
queI'acclimatation hivemale soit
étudiée depuis les années50
(Scholanderet
al., 1950a),notre
compréhensionde
ce
phénomèneest encore
à
développer.En
effet,
nosconnaissances actuelles se
limitent
surtout aux ajustements saisonniers et nous n'avons quepeu
d'informations
surla
capacité des endothermes à ajuster leur physiologie sur une courtepériode de temps.
Pour
approfondir nos coruraissances surla
capacité des endothermes dehivernales, nous avons analysé
la
capacité d'ajustement dela
mésange à têtenoîe
(Poecileatricapillus). Cette thèse regroupe
cinq
chapitres
mettant
en
évidence
les
patronsd'ajustements métaboliques intra-saisonniers
(chapitre
,l),
les
causes exogènesde
cesajustements métaboliques
(chapitre
2),
Ies
mécanismes endogènesqui
permettent
auxanimaux de réaliser ces
ajustements(chapitre
3
et
4)
ainsi que les
conséquences des ajustements phénotypiques hivernaux sur la valeur sélective des individus (chapitre 5).I.5.1
Lespatrons d'ajustements
métaboliquesintra-saisonniers
Les
changements climatiquessont
caractériséspar
rme
augmentationdes
événementsmétéorologiques extrêmes (e.g. innondation, sécheresse) @asterling et a1.,2000). Ces variations
environnementales à court-terme peuvent affecter la démographie, la phénologie @orcada et al.,
2008) et la variabilité génétique des espèces (Canale
&
Henry, 2010) en perturbant, par exemple, les dates de reproduction (Laaksonenet a1.,2006; Van Der Jeugd et a1.,2009) ou en favorisant les génotypes associés aux phénotypes les plus flexibles (Canale&
Henry, 2010). D'après lesmodèles climatiques, les événements extrêmes sont prévus pour être plus frequents, notamment
dans les régions à hautes latitudes (IPCC, 2007), surtout durant la saison hivernale (DesJarlais er
a1,,20I0).
Or, une augmentation dela
stochasticité des conditions hivemales dewaitfaire
del'hiver,
une saison déjà énergétiquement contraignante en région tempérée froide, une étape ducycle
annuel encoreplus
difiicile
à
affronter. Néanmoins, grâceà
leur
capacitéà
ajusterrapidement
et de
façon réversibleleur
physiologie(i.e. flexibilité
phénofypiqug Piersma&
Drent, 2003), les animaux pounaient être en mesure de faire face, jusqu'à une cefiaine limite, à
ces variations météorologiques imprévisibles (Canale
&
Henry,
2010).Afin
de comprendreI'effet
des
fluctuations hivernalessur
la
physiologie des individus
et la
dynamique despopulations (Vedder et
a|.,2013),
il
apparaît doncprimordial
d'étudier la capacité des espèces endothermes résidant en régions tempérées froides de s'ajuster aux variations rapides de leurenvironnement.
Nos connaissances sur I'acclimatation hivemale sont encore très limitées. En effet,
si
les ajustements aufroid
sônt étudiés depuis des décennies (Scholander et a1., 1950c), la plupart desétudes
sur l'acclimatation
hivernalen'ont
fait
qu'analyserles
variations inter-saisonnières (entre été et hiver) de paramètres physiologiques tels que leBMR
ôu le Msum (Cooper, 2002;Cooper
&
Swanson, 1994;Liknes
&
Swansgn, 1996; Swanson, 1991a).Ainsi, bien
que derécentes études se soient intéressées
aux variations
entreles
quatre saisons (Zheng eT al.,2008),
les
études
sur
l'acclimatation hivemale
n'ont
pour
f instant
pas
la
résolution temporelle nécessaire pour mettre en évidence les ajustements potentiellement utilisés par lesendothermes pour répondre aux variations rapides de I'environnement hivernal (McKechnie,
2008; McKechnie
&
Swanson,2010).A
notre connaissance, seules deux études (Broggiet al.,2007;
Swanson&
Olmstead, 1999)
ont
analysé
les
ajustementsà
court
terme
de
la performance métabolique hivernale en conditions naturelles. Swanson&
Olmstead (1999),avec des populations de mésanges
à tête noire,
dejuncos
ardoisés (Junco lryemalis)et
de bruants hudsoniens (Spizella arborea), et Broggi et al. (2007), avec une population de mésangescharbonnières (Parus major),
ont
observé que des oiseaux de petitetaille
augmentaient leur performance métabolique durantl'hiver
(i.e. en quelques semaines) en réponse à la diminutionde la température ambiante. Basé sur ces deux études,
il
sernble que la température ambiante aitun effet proximal sur la performance métabolique hivemale. Nous pouvons donc supposer que
les endothermes de petite
taille
ajustent leur métabolisme au cours deI'hiver
et atteigrrent leur métabolisme maximal durant la période la plus froide del'hiver.
Cependant, aucune étude n'aencore mis en évidence le patron d'ajustement du métabolisme hivemal chez les espèces résidant sous les hautes latitudes. De plus, bien que les ajustements métaboliques inter-saisonniers soient
interprétés comme des exemples de
flexibilité
phénotypique(,iknes
&
Swanson,20lla;Yézina
et
al.,20l1),
nous manquons de données inha-individuelles pour supporter cette assertion.Avec
le
chapitre
/,
nous avonsdonc
analyséles variations du
BMR et
du Msum
enconditions naturelles chez des mésanges
à
têtenoire
capturées plusieursfois au
cours deI'hiver afin
l)
de déterminer le patron hivemal duBMR
et du Msum au sein de la population et2) d'établir si ce patron reflète effectivement la flexibilité phénotypique individuelle.
1.5.2
Les causes exogènes des ajustementshivernaux
L'hiver
sous
les
hautes latitudes
représenteun défi
énergétiquepour
les
espècesendothermes