Utilisation des bactériophages pour le contrôle de
Staphylococcus aureus dans les produits laitiers
Thèse LYNN EL HADDAD Doctorat en microbiologie Philosophiae doctor (Ph. D.) Québec, Canada © Lynn El Haddad, 2014
iii
Résumé
Staphylococcus aureus et ses entérotoxines constituent un risque pour l’industrie
alimentaire ainsi que pour les consommateurs. Environ la moitié des souches de S. aureus peuvent libérer des toxines menant à des symptômes de nausées, de diarrhées et de vomissements chez la personne ayant ingéré un produit contaminé. Une des solutions envisagées pour éliminer S. aureus et éviter la production d’entérotoxines, est l’utilisation d’un cocktail de phages.
Au cours de ce projet de doctorat, trois objectifs ont été développés afin de poursuivre une stratégie de sélection d’un cocktail de phages anti-S. aureus. Tout d’abord, deux phages isolés du milieu laitier, adaptés à celui-ci et respectant des critères de sélection, ont été caractérisés. Ensuite, afin d’éviter un transfert de facteurs de virulence, des myophages anti-S. aureus ont été produits sur une souche de Staphylococcus xylosus, une espèce non-pathogène utilisée en transformation alimentaire et ont été caractérisés afin de confirmer leur identité lorsqu’amplifiés sur les deux espèces. D’un autre côté, l’efficacité contre une panoplie de souches de S. aureus isolées de sources distinctes, l’absence de gènes de virulence dans les génomes phagiques et la résistance à différentes conditions environnementales ont permis de sélectionner des phages différents. Ceux-ci ont fait l’objet de deux cocktails de phages efficaces menant à une réduction significative de la concentration de S. aureus dans des fromages de type Cheddar produits en laboratoire. De plus, ces phages n’ont pas déclenché une surproduction d’entérotoxines staphylococciques C, confirmant la sécurité de leur utilisation dans le milieu laitier. Enfin, la conservation des phages sous forme encapsulée et congelée dans des microbilles de gel d’alginate/calcium semble une approche à approfondir.
Les phages staphylococciques virulents analysés dans cette thèse constituent d’excellents agents de biocontrôle étant non nocifs et infectant spécifiquement une espèce bactérienne donnée. Ayant confirmé l’efficacité de deux cocktails de phages, l’utilisation du produit phagique permettra de diminuer le risque d’apparition de souches bactériennes résistantes aux phages. De plus, entreprendre une mise à jour constante du cocktail phagique permettra de réduire la contamination causée par S. aureus, préservant ainsi l’innocuité et la qualité des aliments.
v
Abstract
Staphylococcus aureus and its enterotoxins pose a risk to the food industry and for
consumers. Approximately half of the S. aureus strains can release toxins leading to symptoms of nausea, diarrhea and vomiting to the person who ingests a contaminated product. One emerging solution to eliminating and preventing S. aureus enterotoxin production is the use of a phage cocktail.
Through this doctoral project, three objectives were developed to pursue a strategy for selecting an anti-S. aureus phage cocktail based on well-defined criteria. First, a methodology was developed leading to the isolation and characterization of two phages from raw milk. Then, in order to avoid a transfer of virulence factors, anti-staphylococcal myophages were produced on a strain of Staphylococcus xylosus, a non-pathogenic species used in food processing. Their genomic identity when propagated separately on the two species was confirmed. Moreover, the host range of these phages was tested against a panel of S. aureus strains isolated from different sources. Their genome was analyzed to confirm the lack of virulence genes. In addition, their resistance to different environmental conditions was used to select different phages for application purposes. These data helped design two efficient phage cocktails leading to a significant drop of S. aureus concentration in small-scale laboratory-based Cheddar cheeses. In addition, they did not trigger the overproduction of staphylococcal enterotoxin C confirming the safety of their use in the dairy environment. Finally, in the aim of commercializing the product, conservation methods were investigated and the encapsulation and freeze of phages in micro-beads consisting of alginate/calcium gel particles appear promising.
Staphylococcal virulent phages seem to be excellent biocontrol agents, being harmless and infecting specifically a given bacterial species. Having confirmed the efficacy of two phage cocktails, the use of the phage product could help decreasing the risk of emergence of phage resistant bacteria. Furthermore, undertaking a constant update of the phage cocktail could also help reducing contamination caused by S. aureus and thereby preserving safety and food quality.
vii
Table des matières
Résumé ... iii
Abstract ... v
Table des matières ... vii
Liste des tableaux ... ix
Liste des figures ... xi
Liste des abréviations ... xiii
Avant-propos ... xv
Chapitre 1. Introduction ... 1
Avant-propos... 1
Staphylococcus aureus, un danger médical et alimentaire ... 1
Danger médical ... 2
Danger alimentaire ... 8
Identification du problème ... 12
Les bactériophages, agents de biocontrôle potentiels ... 14
Historique des phages... 14
Classification des phages ... 15
Cycles lytique et lysogénique ... 17
Critères de sélection des phages destinés à des fins de biocontrôle ... 20
Un phage strictement virulent ...21
Un phage ayant un génome dépourvu de tout facteur de virulence ...22
Un phage polyvalent ...23
Un phage efficace dans tout type de matrice ...24
Un phage ayant une activité stable et une bonne conservation à long terme ...25
Cocktails de phages ... 28
Applications phagiques ... 31
Thérapie par les phages ... 31
Assainissement par les phages ... 33
Biocontrôle et bio-préservation par les phages ... 35
Problématique, hypothèses et objectifs du projet ... 41
Chapitre 2. Characterization of a novel Panton-Valentine leukocidin-encoding staphylococcal phage and its natural PVL-lacking variant ... 43
Résumé ... 43
Avant-propos... 43
Contribution des auteurs ... 43
Publication ... 43
viii
Short-form paper ... 44
Acknowledgments ... 52
References ... 53
Supplemental materials ... 56
Chapitre 3. Improving the safety of a Staphylococcus aureus polyvalent phage by their production on a Staphylococcus xylosus strain ... 61
Résumé ... 611
Avant-propos ... 61
Contribution des auteurs ... 61
Publication ... 62
Abstract ... 62
Introduction ... 63
Materials and Methods ... 64
Results and Discussion ... 69
Acknowledgments ... 77
References ... 78
Supplemental material ... 81
Chapitre 4. Efficacy of two Staphylococcus aureus phage cocktails in a small-scale laboratory-based cheese production ... 89
Résumé ... 89
Avant-propos ... 89
Contribution des auteurs ... 89
Publication ... 90
Abstract ... 90
Introduction ... 90
Materials and Methods ... 92
Results and Discussion ... 98
Acknowledgments ... 109
References ... 109
Chapitre 5. Discussion, conclusion et perspectives ... 113
Problématique de S. aureus ... 113
Stratégie de sélection et d’élaboration de bons agents de biocontrôle staphylococciques ... 114
Évaluation de l’efficacité et de la sécurité de deux cocktails de phages staphylococciques ... 119
Conclusions et perspectives ... 121
Bibliographie ... 123
Liste des tableaux
Tableau 1.1. Protéines extracellulaires de S. aureus ... 6 Table S2.1. ORF identification, putative function, and comparison of LH1 genome with sequences available in public databases ... 56 Table 3.1. Genotyping of the 56 S. aureus strains used in this study. ... 66 Table 3.2. Codon usage of S. aureus JH1 and phages G1 and ISP for the amino acids encoded by the Team1 tRNAs. ... 73 Table 3.3. Host range of phages Team1, phi812, and K propagated on S. aureus SA812 and on S. xylosus SMQ121 ... 736 Table S3.1. ORF identification, putative function, and comparison of Team1 genome with sequences available in public databases ... 82 Table 4.1. Phages used in this study ... 93 Table 4.2. Log10 reduction of phage concentration after heat treatment at 73ºC for 20
seconds ... 98 Table 4.3. EOP values of the 5 phages infecting 57 strains of S. aureus and one strain of S.
xylosus SMQ121. ... 101
xi
Liste des figures
Figure 1.1. Fonctions des protéines de la paroi cellulaire de S. aureus. ... 5
Figure 1.2. Les morphotypes des bactériophages. ... 16
Figure 1.3. Représentation du réseau entre les phages de staphylocoques basée sur le contenu protéomique. ... 17
Figure 1.4. Cycles lytiques et lysogéniques des phages virulents et tempérés. ... 18
Figure 1.5. Micrographes par microscopie électronique à transmission de microsphères d’alginate et d’alginate/carbonate de calcium remplies du phage K. ... 26
Figure 1.6. Exemple de l’efficacité des cocktails de phages. ... 29
Figure 1.7. Effet d’un cocktail de phages (phiIPLA35 et phiIPLA88) sur la croissance de S. aureus Sa9 dans un fromage à pâte pressée durant l’affinage. ... 39
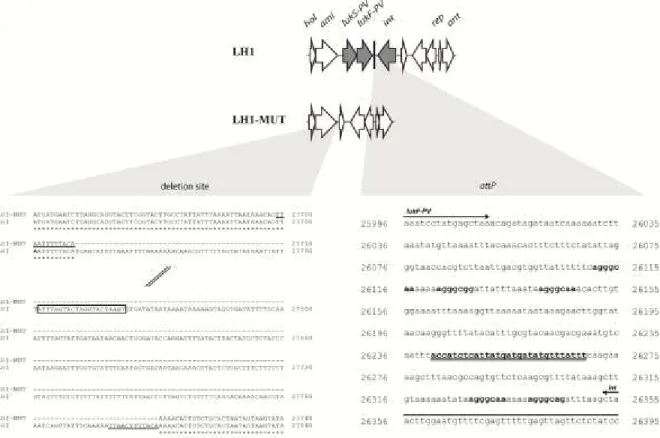
Figure 2.1. Genome alignments of LH1 and LH1-MUT, proteins bands shown on a SDS-PAGE, and identification of the different structural proteins. ... 48
Figure 2.2. Part of the LH1 genome showing the deleted region in LH1-MUT genome and the nucleotide sequence of the attachment site attP. ... 50
Figure S2.1. Electron micrograph of S. aureus phage LH1. ... 57
Figure S2.2. Genome alignments of all PVL-carrying phages available including LH1. ... 58
Figure S2.3. Comparison of staphylococcal phage complete proteome ... 59
Figure 3.1. Electron micrograph of phage Team1. ... 69
Figure 3.2. One-step growth curves of both Team1-SA812 and Team1-SMQ121 ... 70
Figure 3.3. Circular genome comparison of phages Team1, G1, and ISP. ... 72
Figure 4.1. Monitoring phage concentration in Cheddar cheese curd at pH 5.5, stored at 4ºC for 28 days... 99
Figure 4.2. Monitoring staphylococcal and phage counts over time during a small-scale, laboratory-based Cheddar cheese production ... 104
Figure 4.3. Monitoring phage concentration with or without encapsulation in beads in fresh, freezed, and lyophilised states. ... 107
Figure 4.4. Free phage and encapsulated phage concentration monitoring during 3 months of storage at 4ºC and -20ºC ... 108
xiii
Liste des abréviations
ADN acide désoxyribonucléique
ADNdb acide désoxyribonucléique double brin ADNsb acide désoxyribonucléique simple brin ARN acide ribonucléique
ARNm acide ribonucléique messager ARNr acide ribonucléique ribosomique ARNt acide ribonucléique de transfert
Aw activité de l’eau
BIM mutant bactérien résistant aux bactériophages (bacteriophage- insensitive mutant)
CA-SARM S. aureus résistant à la méthicilline associé à la communauté
CC complexe clonal
CI Intervalle de confiance (confidence interval) CoNS staphylocoque à coagulase négative
CoPS staphylocoque à coagulase positive
Db double brin
ES entérotoxine
ET exfoliatine
EOP efficiency of plaquing
FDA food and drug administration
G + C contenu en guanine et cytosine de l’ADN
HA-SARM S. aureus résistant à la méthicilline acquis dans les hôpitaux
Kpb kilo paires de bases
LukS-PV sous-unité S de la toxine PVL LukF-PV sous-unité F de la toxine PVL
MAPAQ ministère de l’agriculture, des pêcheries et de l’alimentation du Québec
µl microlitre
ml millilitre
MLST multi-locus sequence type
MOI multiplicité d’infection (multiplicity of infection)
MSCRAMM protéines de sa paroi cellulaire (microbial surface components recognizing adhesive matrix molecules)
nm nanomètre
ORF cadre de lecture ouvert (open reading frame)
Pb paire de bases
PBP protéine liée à la Pénicilline (Penicillin-binding protein)
PCR réaction en chaîne de la polymérase (polymerase chain reaction) PVL Panton-Valentine leucocidine
PSM moduline soluble au phénol (phenol-soluble modulin) RCRMB réseau canadien de recherche sur la mammite bovine SARM S. aureus résistant à la méthicilline
Sb simple brin
SCC cassette chromosomique staphylococcique SCV petite variante de colonie (small colony variant)
xiv
SDS-PAGE électrophorèse sur gel de polyacrylamide contenant du sodium (sodium dodecyl sulfate polyacrylamide gel electrophoresis)
ST séquence type
TSA tryptic soy agar
TSB tryptic soy broth
TSST toxine du syndrome de choc toxique UFC unité formatrice de colonie
UFP unité formatrice de plaque UHT ultra-haute température
xv
Avant-propos
Résultats trouvés. Conclusions retirées. Article publié. Thèse déposée. Une succession de faits résumant quatre longues années de travail soulignées par une réussite bien méritée. Je dois ce succès à plusieurs personnes qui m’ont inspirées, encouragées et fortifiées au cours de ces années.
Je me permets tout d’abord de remercier mon directeur de thèse, Prof Sylvain Moineau, sans qui cette étape n’aurait pas été accomplie. Merci Sylvain pour tes conseils indispensables, pour ta présence inconditionnelle et pour la confiance que tu m’as accordée. C’est grâce à toi que mon enthousiasme, ma curiosité et mon esprit scientifiques se sont développés et m’ont permis d’atteindre cette étape aujourd’hui.
Je tiens aussi à remercier mon co-directeur de thèse ainsi que mon comité aviseur, Prof Steve Labrie, Dr Claude Champagne et Dr Daniel St-Gelais. Merci pour votre confiance en mon potentiel, pour vos mots encourageants, vos expertises respectives et surtout pour avoir été présents tout au long de mon cheminement scientifique. Je remercie aussi Dr Lawrence Goodridge d’avoir accepté le rôle d’examinateur externe.
Je voudrais aussi remercier les organismes subventionnaires, FQRNT, MAPAQ, AAC et Novalait qui m’ont permis de réaliser ce parcours.
Un grand merci à mes collègues de travail, passés et présents. Merci pour votre simple présence qui a rendu l’environnement de travail chaleureux et joyeux. Je voudrais surtout remercier ma collègue de travail, mon amie depuis mon premier jour au Canada, Siham. Merci pour tes conseils fraternels, merci d’avoir partagé ma joie et mes moments stressants. Merci tout simplement d’avoir été ma grande sœur durant mon parcours au Canada.
Je voudrais aussi remercier tous mes amis et ma famille libanaise au Québec ainsi que ma famille québécoise. La liste est tellement longue que je ne pourrais citer tout le monde un par un. Je suis bénie de vous avoir tous dans ma vie. Vous m’avez tous submergée d’amour. Votre présence m’a réconfortée et m’a permis de surmonter toutes les difficultés rencontrées.
xvi
Merci à mon adorable frère Alain. J’ai passé deux années complètes à tes côtés et j’ai appris à mieux te connaitre. Merci d’avoir supporté mes crises de folie et mon stress, surtout en fin de parcours doctoral. Tu as toujours su quoi faire et quoi dire durant ces moments et je te suis à jamais reconnaissante. Je remercie aussi mon frère Nicolas et ma sœur Léa qui, malgré les distances, ont été présents chaque jour à mes côtés. Je vous aime beaucoup vous trois!
Je tiens à remercier du fond du cœur mes parents, Paula et Sami. Vous êtes et vous resterez mes idoles. Vous êtes l’exemple des parents forts, consacrés à offrir le meilleur des vies à leurs enfants même si ceci conduit à un éloignement géographique. Merci d’avoir eu confiance en moi et d’avoir eu le courage de m’envoyer seule à un pays si loin. Je vous dédie mon succès et j’espère que vous êtes fiers de moi. Je vous aime!
Un grand merci à mon amour, à l’homme de ma vie, Georges. Tu as tellement enduré pendant toutes ces années à cause des distances géographiques entre nous. Je te remercie d’avoir toujours été là pour moi et d’avoir partagé mes moments de stress, de solitude et de joie. Merci d’avoir toujours eu confiance en moi et en mon potentiel. La fin approche et notre vie ensemble commencera bientôt! Je t’aime!
Je remercie Dieu chaque jour de m’avoir entourée de personnes si précieuses. C’est grâce à vous tous que j’achève cette étape de vie avec succès et que je quitte le Canada en gardant plein de beaux souvenirs inoubliables au fond de mon cœur.
When you reach the end of what you should know, you will be at the beginning of what you should sense.
1
Chapitre 1. Introduction
Avant-propos
L’introduction de la thèse est divisée en trois sections. La première section décrit le microorganisme Staphylococcus aureus qui fait l’objet de cette thèse et sa problématique dans les produits laitiers. Ensuite, une deuxième section consiste à développer une des solutions de lutte contre S. aureus, soit les bactériophages virulents. Enfin, les applications médicales et alimentaires des phages anti-staphylocoques sont détaillées dans la dernière section.
Staphylococcus aureus, un danger médical et alimentaire
S. aureus appartient à la famille des Micrococcaceae et au genre Staphylococcus qui
regroupe 41 espèces reconnues jusqu’à présent (Gillaspy & Iandolo, 2009). C’est en l’an 1880 que la première description d’une infection suppurative par un staphylocoque a été observée en Écosse et a été nommée par le chirurgien Alexander Ogston (Ogston, 1984). L’adjectif aureus (ou "or" en latin) lui a été attribué en 1884 par Anton J. Rosenbach suite à la couleur formée par les colonies sur du milieu sang (Rosenbach, 1884).
Ce qui diffère les staphylocoques des autres membres de cette famille est leur contenu faible en G+C d’environ 32,8 % (Dlawer & Hiramatsu, 2004), leur tolérance élevée au sel dans le milieu (jusqu’à 20 %) ainsi que leur résistance à la digestion par le lysozyme grâce à son taux élevé d’O-acétylation au niveau de la couche de peptidoglycanes (Gillaspy & Iandolo, 2009).
S. aureus est un coque à Gram positif d’environ 0,7 à 1,2 micromètre de diamètre qui se
trouve seul, en paires ou en grappe dans divers milieux liquides et solides. Cette bactérie aérobie ou anaérobie facultative a une température optimale de croissance de 37 ºC lui conférant le caractère de mésophile.
S. aureus est considérée avant tout comme une bactérie commensale. En effet, elle fait
partie de la microflore normale de la peau, du tractus intestinal et du nasopharynx. Dix à 35 % de la population générale sont des porteurs sains permanents et 60 % des individus
2
sont des porteurs sains temporaires de S. aureus (Edwards & Massey, 2011). Cependant, S.
aureus peut se retrouver dans d’autres niches écologiques dont la terre, l’air, l’eau et les
aliments dont les produits laitiers. Certaines souches infectent les mammifères incluant l’humain. C’est le staphylocoque à coagulase positif le plus isolé d’infections humaines puisqu’il est capable de produire l’enzyme menant à la coagulation du plasma sanguin. Lorsque cette bactérie se transmet d’un individu malade à l’autre, change d’habitat normal ou est exposée à une situation de stress, elle s’attache, colonise et déchaîne ses facteurs de virulence multiples. De ce fait, S. aureus présente un danger de santé publique, que ce soit au niveau médical ou alimentaire.
Danger médical
S. aureus est connue par sa prévalence dans les lieux hospitaliers et son transfert rapide
dans la communauté. En effet, environ 1 200 000 cas d’infections par S. aureus/an sont notés dans les hôpitaux aux États-Unis (www.cdc.org/) ainsi que 28,4 cas d’infections par
S. aureus/100 000 habitants par an dans la région de Calgary au Canada (Laupland et al.,
2003).
Le problème de santé publique majeur relié à S. aureus et présent partout dans le monde est celui de sa résistance aux antibiotiques. À cause d’une exposition fréquente à différents types d’antibiotiques, ce pathogène est de plus en plus résistants à ceux-ci, dont notamment à la famille des bêta-lactames (méthicilline, pénicilline, etc.). Ces souches pathogènes prennent le nom de SARMs (S. aureus résistants à la méthicilline). Cette résistance est à l’origine de la présence d’un gène nommé mecA localisé dans la cassette chromosomique staphylococcique mec (ou SCCmec). Le gène mecA encode une protéine membranaire appelée PBP-2a. Celle-ci confère une faible affinité aux antibiotiques de la famille des bêta-lactame contrairement à la protéine PBP, empêchant ainsi l’affaiblissement de la paroi bactérienne par les antibiotiques, la lyse et la mort cellulaires (Conly & Johnston, 2003; Lim & Strynadka, 2002; Lowy, 2003).
Aux États-Unis, il a été estimé que les infections par les SARMs sont plus létales que la mortalité associée au VIH/SIDA (Bancroft, 2007; Peschel & Otto, 2013). Le taux de maladies reliées aux SARMs est 17 fois plus élevé dans les hôpitaux du Canada depuis
3 l’année 1995 jusqu’à l’année 2010 (Public Health Agency of Canada, 2013). Celles-ci sont aussi très coûteuses. On note un coût total relié aux infections par les SARMs d’environ 33 à 85 millions de dollars dans les hôpitaux canadiens en l’an 2005 (Goetghebeur et al., 2007) avec une moyenne annuelle de 36 millions de dollars jusqu’en 2013 (Public Health Agency of Canada, 2013).
Il existe deux types de SARMs. Ceux acquis dans les hôpitaux (HA-SARM) et ceux associés aux disséminations dans la communauté (CA-SARM). Au Canada, 10 souches différentes majeures de SARMs ont été identifiées et isolées jusqu’à présent (Nichol et al., 2011). Les HA-SARMs sont nosocomiales et surviennent des plaies de patients infectés, de cathéters, d’une hospitalisation prolongée, mais aussi de la peau de porteurs sains (Furuya & Lowy, 2006). Ils sont résistants à plusieurs types d’antibiotiques (érythromycine, tétracyclines, etc.) alors que les CA-SARMs ne sont résistants qu’aux antibiotiques composés de bêta-lactames (Furuya & Lowy, 2006). Ils sont à l’origine de cas de morbidité et de mortalité (Chen et al., 2011) et se disséminent rapidement dans la communauté (Goetghebeur et al., 2007; Nichol et al., 2011).
S. aureus est responsable de trois principaux types de maladies. La première est
caractérisée par des infections cutanées et superficielles comme les impétigos, les furoncles, les ulcères, les blessures infectées et les abcès. Dans le cas des bovins, ce genre d’infection est appelé une mammite (infection des glandes mammaires bovines). Ces infections suppuratives sont traitées rapidement et facilement par l’incision et le drainage de l’infection accompagnés d’antibiotiques ou par l’application d’une pommade antibactérienne (Edwards & Massey, 2011; McCaig et al., 2006).
Le deuxième type de maladies rassemble les infections systémiques staphylococciques. Ces dernières constituent l’une des plus communes et sérieuses infections bactériennes dans le monde et peuvent causer la mort de l’individu suite à la multiplication et la propagation de S. aureus dans le sang causant une bactériémie et un choc septique létal (Thwaites & Gant, 2011). Une bactériémie aggravée mène à l’entrée de la bactérie dans les tissus environnants et peut causer une endocardite, des abcès au niveau des organes, une arthrite, une ostéomyélite vertébrale ou même une méningite (Edwards & Massey, 2011; Miller & Cho, 2011). Plusieurs études montrent la façon par laquelle S. aureus est capable de
4
s’adapter à son hôte et de fuir le système immunitaire (Edwards & Massey, 2011). Premièrement, la capsule polysaccharidique semble permettre à S. aureus de résister aux mécanismes de défense de l’hôte comme l’opsonisation et la phagocytose (Gillaspy & Iandolo, 2009). L’entrée et la persistance à l’intérieur des neutrophiles par phagocytose paraissent aider S. aureus à fuir le système immunitaire (Edwards & Massey, 2011). Deuxièmement, ce pathogène peut profiter de l’apport en fer en s’attachant aux globules rouges humains. Le fer étant un nutriment essentiel pour sa survie et sa virulence, il utilise son enzyme hémolysine pour s’emparer de l’hème et le dégrader en fer (Lowy, 2011).
Une caractéristique spécifique à S. aureus est sa capacité à envahir une cellule hôte non phagocytaire. Une fois à l’intérieur de ces cellules, S. aureus peut induire une apoptose ou une nécrose cellulaire en les endommageant par ses cytotoxines, ou bien, persister sous un état semi-dormant. Ce phénotype, appelé variante de petite colonie (ou small colony variant, SCV), est protégé contre les attaques du système immunitaire et contre l’effet des antibiotiques (Foster et al., 2014; Tuchscherr et al., 2010). Une étude a montré que les SCVs peuvent demeurer plusieurs semaines à l’intérieur de la cellule hôte. De plus, lorsque ces bactéries se « réveillent », elles reprennent leur virulence et infectent rapidement et dynamiquement d’autres cellules de l’hôte (Tuchscherr et al., 2011). Ce changement de phénotype semble essentiel pour le processus d’infection de S. aureus et est à l’origine des infections chroniques et récurrentes (Garzoni & Kelley, 2011) (Figure 1.1).
5 Figure 1.1. Fonctions des protéines de la paroi cellulaire de S. aureus. Figure tirée de Foster et al. (2014).
Toutes ces stratégies de protection, de colonisation et de survie de S. aureus sont rendues possibles grâce, principalement, aux protéines de sa paroi cellulaire ou MSCRAMMs (microbial surface components recognizing adhesive matrix molecules) (Tableau 1.1).
Un troisième type de maladie causée par S. aureus concerne les toxinoses, ou les symptômes déclenchés par la sécrétion de toxines dans le milieu. C’est le cas des exfoliatines (ETs), de la toxine Panton-Valentine leucocidine (PVL), de la toxine du syndrome de choc toxique (TSST-1) et des entérotoxines (ESs) (Tableau 1.1).
6
Tableau 1.1. Protéines extracellulaires de S. aureus. Tableau adapté de Gillaspy & Iandolo (2009).
Protéine Localisation du gène
Hémolysines
• Toxine Alpha Chromosome
• Toxine Béta Chromosome
• Toxine Gamma Chromosome
• Toxine Delta Chromosome
• Panton-Valentine Leucocidines Chromosome/îlots de pathogénicité Entérotoxines • ES A Bactériophage/chromosome • ES B Chromosome/îlots de pathogénicité • ES C Plasmide • ES D Plasmide • ES E Chromosome • ES G Chromosome • ES I Chromosome
Enzymes et autres toxines
Lipase Chromosome
Nucléase Chromosome
Protéase V8 Chromosome
Estérase Chromosome
Coagulase Chromosome
Hydrolase de la paroi cellulaire Chromosome
Hyaluronidase Chromosome
Staphylokinase Bactériophage/chromosome
Protéine A Chromosome
Protéases sériques Chromosome/îlots génomiques
Phospholipase C Chromosome
Leucotoxines Chromosome/îlots génomiques Leucocidines Chromosome/îlots de pathogénicité Exfoliatine A (ETA) Chromosome
Exfoliatine B (ETB) Plasmide
Toxine du syndrome du choc toxique-1 (ES F) Chromosome/îlots de pathogénicité MSCRAAMs
Facteurs d’agglutination Chromosome Protéines A/B liées à la fibronectine Chromosome Protéine liée au fibrinogène Chromosome Protéine liée au collagène Chromosome Protéine liée à l’élastine Chromosome Protéines liée à la matrice extracellulaire Chromosome Protéines d’adhésion intercellulaire Chromosome
Les ETs dont ETA et ETB sont des protéases sécrétées par S. aureus qui entraînent la disparition des vésicules intercellulaires. Et ce, en ciblant la glycoprotéine desmogléine-1 responsable du maintien de l’adhésion cellulaire. Ceci donne lieu à la formation d'une
7 couche liquide entre les couches granuleuse et superficielle de l’épiderme et se manifeste macroscopiquement par le décollement intradermique. Lorsque ce phénomène est localisé grâce à la présence d’anticorps, il prend le nom de syndrome bulleux épidermique alors qu’il se nomme syndrome de la peau ébouillantée ou syndrome de Ritter lorsqu’il est généralisé (Bukowski et al., 2010; Larkin et al., 2009; Podbielska et al., 2011).
La toxine PVL, codée par un prophage, est responsable de la formation de pores dans les neutrophiles et les macrophages et entraîne une apoptose cellulaire et des infections nécrotiques cutanées et pulmonaires. Cette toxine est composée de deux sous-unités, LukS-PV et LukF-LukS-PV qui s’assemblent et agissent de façon synergique pour former des pores (Kaneko & Kamio, 2004). Les toxinoses causées par la PVL sont plus fréquentes prévalentes chez les nourrissons et les enfants, les personnes immunosupprimées, les travailleurs du domaine de la santé, les patients hospitalisés et chez les personnes ayant des infections staphylococciques suppuratives (Vandenesch et al., 2003). Les SARMs sont aussi responsables de la transmission et de la propagation de cette toxine dans la communauté, à travers le monde (Barnes & Sampson, 2011; Blanco et al., 2011; Boakes et
al., 2011; Chen et al., 2011; Nagao et al., 2010; Zhang et al., 2008).
Le syndrome du choc toxique est causé par la toxine TSST-1. Celle-ci porte aussi le nom de ES F ou entérotoxine staphylococcique F. Tout comme les exfoliatines et les autres types d’entérotoxines alimentaires, TSST-1 est superantigénique. Elle cause la stimulation accrue d’un vaste nombre de lymphocytes T et le déclenchement inapproprié d’une libération massive de cytokines affaiblissant le système immunitaire de l’individu (Kum et
al., 2001; Raulin et al., 2010). La TSST-1 se caractérise par une apparition soudaine de
fièvre, une hypotension artérielle, des rougeurs, une défaillance d’organes multiples et une desquamation notamment des paumes des mains et des plantes des pieds (Podbielska et al., 2011). Ce syndrome est causé aussi bien par S. aureus que par Streptococcus pyogenes du groupe A (Lappin & Ferguson, 2009).
Une nouvelle classe de toxines a été identifiée depuis l’an 2000, celles des modulines solubles au phénol ou PSMs. Ces peptides contribuent à la virulence de S. aureus et surtout des CA-SARMs. Elles peuvent tout d’abord favoriser la structure, le développement et le détachement des biofilms ainsi que faciliter la dissémination des infections staphylococciques associées aux biofilms aux autres organes du corps. De plus, les PSMs,
8
n’étant pas dépendants d’un récepteur, ciblent presque toutes les membranes cytoplasmiques eucaryotes avec un tropisme pour les leucocytes et les érythrocytes. Ces peptides restent peu étudiés, mais font l’objet de cibles prometteuses pour le développement de vaccins anti-staphylococciques (Peschel & Otto, 2013). Plus particulièrement, le système de sécrétion de ces protéines responsables de la lyse des neutrophiles, et essentiel à la croissance bactérienne, a été identifié récemment. Ce système de sécrétion de toxine constitue une cible potentielle pouvant inhiber la croissance et la virulence de S. aureus (Chatterjee et al., 2013).
D’autres types d’ESs superantigéniques se retrouvent dans la liste des facteurs de virulence de S. aureus et seront traitées dans la section suivante. Celles-ci sont reliées aux maladies d’origine alimentaire.
Danger alimentaire
S. aureus est aussi un pathogène alimentaire contaminant, entre autres, le lait et les
dérivés laitiers. En effet, le lait est un milieu riche et propice pour la croissance d’organismes de par le fait qu’il contient une teneur en eau élevée, une abondance de nutriments et un pH près de la neutralité (entre 6,4 et 6,8) (Touch & Deeth, 2009). Des microorganismes d’altérations et pathogènes peuvent se retrouver dans tout type de lait, que ce soit du lait cru, pasteurisé, à durée de conservation prolongée (ESL) ou traité à une ultra-haute température (UHT). La majorité des intoxications liées à la consommation de lait cru sont causées par les bactéries pathogènes Listeria monocytogenes, Escherichia coli,
Campylobacter jejuni, Salmonella spp., Yersinia enterocolitica et S. aureus. Leur
prévalence dans le lait cru dépend de la région géographique, de la saison, du nombre d’animaux dans la ferme et de l’hygiène du lieu et du personnel (Touch & Deeth, 2009).
Pour ce qui est de S. aureus, sa présence dans le lait cru et les produits laitiers peut être due à plusieurs facteurs. Premièrement, une mammite ou inflammation des glandes mammaires de ruminants peut conduire au transfert de S. aureus dans le lait cru. Une étude a montré que presque 45 % des échantillons de lait cru bovin testés contenaient des souches staphylococciques à coagulase positive (Rola Jolanta et al., 2013). Ces souches staphylococciques retrouvées dans le lait sont généralement géographiquement différents
9 ou endémiques à la ferme et à la région dans laquelle elles ont été isolées (Piccinini et al., 2010; Proietti et al., 2010).
Deuxièmement, un manque d’hygiène du personnel durant la traite ou durant la manipulation du lait et ses dérivés et l’utilisation impropre d’appareils industriels peuvent affecter l’innocuité du produit laitier. D’autre part, une acidification insuffisante et lente du caillé peut mener à des épidémies staphylococciques associées à ces produits (Le Loir et
al., 2003). Dans le cas des produits laitiers pasteurisés, les souches de S. aureus sont
détruites après le traitement à la chaleur. Ainsi, une présence de S. aureus dans ces aliments est considérée comme un indicateur d’une contamination après le traitement par la chaleur et donc, un manque d’hygiène (Asao, 2003; Bennett et al., 2013). Ce critère non respecté pourrait être à l’origine de la présence de SARMs dans ces aliments et de souches hypermutables résistantes à plusieurs types d’antibiotiques et porteuses de facteurs de virulence (Graham et al., 2006; Normanno et al., 2007; Soares et al., 2011; Wang et al., 2013).
Le dernier facteur favorisant la présence de S. aureus dans l’aliment est le non-respect des conditions de transformation de l’aliment ou l’utilisation de conditions inadéquates durant les différentes étapes de fabrication, de stockage et de transport du produit fini. Plusieurs facteurs comme la température, le pH, l’activité de l’eau (Aw), la concentration de
sel et la compétition nutritionnelle de la microflore du milieu affectent la croissance de
S. aureus et sa production de toxines. De plus, les conditions environnementales pendant la
phase initiale de production de fromage est presque optimale pour la multiplication de
S. aureus (Bennett et al., 2013; Rola Jolanta et al., 2013). Donc, S. aureus peut être présent
dans les produits laitiers issus de lait pasteurisé et cru (Coveney et al., 1994).
Plus particulièrement, certaines souches de S. aureus, à une concentration spécifique dans l’aliment, peuvent sécréter des ESs dans un milieu à Aw élevée, à un pH entre 4,8 et
9,0 et à une température optimale entre 34 et 40 ºC. Environ 50 % des souches de S. aureus sont productrices de ESs (Su & Lee Wong, 1997; Sutra, 1998). Il a été démontré qu’une concentration de 105 unités formatrices de colonies staphylococciques (UFC) par millilitre ou par gramme d’aliment sont nécessaires pour qu’il y ait production et sécrétion de ES dans le milieu. Cependant, certaines souches virulentes de S. aureus sont capables de libérer leurs ESs à une concentration de 103 UFC/ml (Meyrand et al., 1998).
10
Une fois sécrétées dans l’aliment, une concentration en entérotoxines de l’ordre du nanogramme peut déclencher les symptômes d’une intoxication alimentaire (diarrhées, fièvre, nausées, crampes abdominales, maux de tête) à la personne ingérant l’aliment contaminé (Hennekinne et al., 2012; Krakauer & Stiles, 2013; Ostyn et al., 2010). Ces symptômes peuvent être bénins comme mentionnés plus haut, mais, dans des cas rares, ils se déclenchent par des variations de la pression artérielle et même par la mort de la personne par entérotoxicose et déshydratation (Balaban & Rasooly, 2000; Do Carmo et al., 2004). Ces derniers symptômes surviennent plus fréquemment chez les enfants et les personnes âgées (Do Carmo et al., 2004).
Les ESs sont des protéines solubles dans l’eau et dans les solutions salines. Elles tolèrent des conditions environnementales extrêmes (séchage, congélation) et maintiennent leur activité biologique dans le tractus digestif après ingestion, en résistant à l’activité protéolytique des enzymes digestives, comme la pepsine et la trypsine (Hennekinne et al., 2012; Le Loir et al., 2003), contrairement aux cellules de S. aureus. De plus, elles sont thermorésistantes et peuvent tolérer une température de 121 ºC pendant 28 minutes (Balaban & Rasooly, 2000).
Il existe jusqu’à présent, 23 différentes ESs, dont les cinq principales ES A à E, de nouveaux ESs (ES G à I) et des « ESs-like » (ESI J à X) (Alibayov et al., 2014; Hennekinne
et al., 2012). Les ESs et les « ESs-like » présentent tous deux un effet superantigénique
puisqu’ils activent les lymphocytes T et stimulent la prolifération cellulaire et le relargage massif des cytokines. Ceci va déclencher une réponse pro-inflammatoire et affaiblir le système immunitaire contre les infections bactériennes (Gillaspy & Iandolo, 2009; Larkin
et al., 2009; Podbielska et al., 2011). Ce qui diffère ces deux types est la capacité des ESs à
déclencher une réponse émétique ou vomitive suite à une intoxication, alors que les «
ESs-like » ne le sont pas ou n’ont pas encore été testés efficacement sur des primates (Lina et al., 2004).
Les toxines communément rencontrées sont celles de type A, qui sont attribuables à des souches virulentes chez l'humain et celles de types B et C, qui sont attribuables à des souches isolées des bovins et caprins (Villeneuve et al., 2007). En effet, les souches de
S. aureus isolées d’échantillons de lait de chèvre et de vache sont, le plus souvent,
11 La première description d’intoxication alimentaire staphylococcique date de 1884. Elle a été investiguée par Vaughan et Sternberg. Ces derniers ont montré que cette intoxication est survenue à la suite de la consommation de fromages contaminés (Hennekinne et al., 2012). Depuis, il y eut plusieurs épidémies d’intoxications alimentaires. Mentionnons celle qui s’est déroulée en Osaka, au Japon, en l’an 2000 et lors de laquelle treize mille personnes ont été intoxiquées à cause d’une quantité minime de ES A présente dans du lait en poudre et liquide (Ikeda et al., 2005). Cette entérotoxine est la plus abondante des ESs. Ceci s’explique par une différence entre les ESs au niveau de leur production et la sécrétion. Une étude récente a étudié l’expression et la production des principales ESs lors de la croissance bactérienne de souches staphylococciques isolées de milieux alimentaires (Derzelle et al., 2009). Les auteurs ont observé une production de ES A durant toute la phase exponentielle de croissance de S. aureus (Derzelle et al., 2009; Wallin-Carlquist et
al., 2010a). À l’opposé, les ES B, C et D sont produits seulement durant la transition entre
les phases exponentielle et stationnaire (Derzelle et al., 2009).
De plus, ces entérotoxines sont situées sur différents éléments génétiques mobiles comme des prophages (le gène de l’entérotoxine staphylococciques A ou sea), des plasmides (seb, sec, sed), des îlots de pathogénicité et aussi sur des transposons, facilitant ainsi des transferts horizontaux de gènes. En effet, le gène seb est trouvé sur un îlot de pathogénicité dans certaines souches et sur un plasmide dans d’autres (Hennekinne et al., 2012). Cependant, la présence du gène de toxine dans le matériel génétique de S. aureus n’implique pas nécessairement la production de la toxine (Loncarevic et al., 2005).
Bien que les intoxications alimentaires staphylococciques ne représentent pas une maladie très sévère, elles peuvent mener à une panique sociale, touchent à la santé publique et font l’objet d’une charge sociale considérable (Su & Lee Wong, 1997). De ce fait, des limites de comptes bactériens staphylococciques ont été établies par les agences gouvernementales de chaque région. Au Québec, la loi sur les produits alimentaires P-29 stipule que la concentration maximale acceptée de S. aureus ne doit pas excéder 2 000 UFC/ml dans le lait cru et 1000 UFC/g dans le fromage issu du lait cru. Pour ce qui est du fromage issu de lait pasteurisé, celui-ci doit contenir une concentration maximale de 100 UFC/g pour passer le contrôle de qualité (Commission Canadienne du Lait, 2011). Un
12
non-respect de ces normes mène au rejet des lots de laits et de fromages contaminés et une perte économique.
Identification du problème
S. aureus est à l’origine d’infections humaines et d’intoxications alimentaires. Ces deux
différents dangers staphylococciques peuvent être détectés suite à l’identification, au génotypage et au phagotypage (ou lysotypie, typage par les phages) des souches de
S. aureus.
Effectivement, il existe des méthodes de typage qui regroupent les souches bactériennes selon leur parenté génétique. Ceci permet de reconstruire la chaîne d’infection et les possibles évènements de l’évolution d’une espèce bactérienne donnée se produisant sur de courts délais, en considérant les taux élevés de recombinaisons génétiques (Feil et al., 2004; Wolf et al., 2011).
L’une des méthodes permettant le génotypage des souches bactériennes est l’obtention de séquence type par MLST (multilocus sequence type). Celle-ci est hautement discriminatoire et caractérise les isolats bactériens sur la base de séquences d’une longueur d’environ 450 paires de bases pour chacun des sept gènes domestiques ou « housekeeping genes » sélectionnés. Chez S. aureus, ces sept gènes sont : arcC (Carbamate kinase), aroE (Shikimate déhydrogenase), glpF (Glycérol kinase), gmk (Guanylate kinase), pta (Phosphate acetyltransférase), tpi (Triosephosphate isomérase) et yqi (Acétyl coenzyme A acétyltransférase). Les fragments des gènes glpF et gmk sont les plus uniformes parmi les sept gènes utilisés dans la méthode de MLST alors que les gènes arcC et aroE sont les plus variables. Chaque gène constitue un allèle ou un locus et la combinaison des 7 loci donne lieu à un profil allélique ou séquence type (ST). Les variations nucléotidiques à l’intérieur de chaque gène s’accumulent relativement lentement au cours de l'évolution de l'espèce, permettant d’identifier l’ancêtre commun d’un ST et ses descendants. L’avantage de cette méthode est la comparaison immédiate des données obtenues avec celles d’autres laboratoires de recherche, et ce, sur la base de données accessible en ligne (Enright et al., 2000).
Par la suite, un algorithme appelé eBURST a été développé permettant le regroupement des STs en des complexes clonaux (CCs) (Feil et al., 2004). Un CC est
13 composé d’un ST prédominant et relié à des STs relativement proches, différant d’un (SLV ou variant d’un seul locus) ou de deux loci (DLV ou variant de double loci). On présume que plus la fréquence du ST prédominant augmente dans la population, plus il évolue graduellement, se diversifie (suite à des évènements de recombinaison et des mutations ponctuelles) et donne lieu à des STs proches. Cet algorithme peut être utilisé pour expliquer l’émergence et la diversité des clones bactériens.
Plusieurs études ont adapté ces méthodes de génotypage et de regroupement pour caractériser les souches staphylococciques isolées localement, qu’elles soient d’origine humaine ou animale. Dans le cas de souches de SARMs ou de souches de S. aureus sensibles à la méthicilline (SASMs), Enright et al. (2000) ont comparé 155 isolats de S.
aureus isolés d’infections nosocomiales et communautaires de la région d’Oxford
(Angleterre). Ceux-ci ont observé des clones majeurs de SARMs appartenant majoritairement à ST36. Les membres de ce ST semblent provenir du groupe ST30 (comportant des SASMs). Ces clones ont supposément évolués en acquérant le gène mecA par transfert horizontal leur conférant une résistance aux bêta-lactames et ont bifurqué dans le groupe ST36. D’autres SASMs de la région sont rassemblés dans les groupes ST25, ST8, ST39, ST1 et ST5. Une deuxième étude a comparé les CCs de souches staphylococciques humaines et bovines et a identifié un transfert de souches humaines aux bovins. C’est le cas de CC8 adapté aux bovins qui commence à se propager de la Suisse à travers l’Europe (Sakwinska et al., 2011). D’autres CCs sont exclusivement d’origine bovine comme le CC97 et le CC151 et sont distribués dans tout le globe (Wolf et al., 2011) alors que la prédominance d’autres groupes est relative à la région de laquelle les souches ont été isolées. Par exemple, une prédominance de ST126 est observée au Brésil et alors qu’en Norvège, c’est le groupe ST133 qui est prédominant (Sakwinska et al., 2011). Ce dernier ST est aussi observé dans des régions françaises et allemandes (Wolf et al., 2011).
L’identification du problème accentue l’importance de la recherche de procédés pouvant éradiquer ou réduire significativement la concentration de S. aureus dans les divers milieux tout en maintenant la biodiversité et l’écologie microbienne de ces environnements. Par ce fait, l’utilisation de bactériophages comme agents de biocontrôle est envisagée au cours de cette étude.
14
Les bactériophages, agents de biocontrôle potentiels
Comme leur nom l’indique, les bactériophages, ou phages, sont des virus qui « mangent » ou attaquent les bactéries. Ces entités lysent entre 4 à 50 % des bactéries produites chaque jour (Breitbart & Rohwer, 2005). Ce sont les entités biologiques les plus abondantes sur la Terre et sont présentes dans tout type d’environnement incluant les aliments (Brüssow, 2005). On estime qu’il existe plus de 1031 de phages sur la Terre (Clokie et al., 2011). Un millilitre d’eau de mer peut contenir un million de bactéries et environ 10 millions de phages (Breitbart & Rohwer, 2005). En effet, ceux-ci sont régulièrement et accidentellement consommés à travers l’ingestion d’eau et d’aliments (Mahony et al., 2011). Leur capacité à lyser spécifiquement une espèce bactérienne donnée, à se répliquer exponentiellement, à être sécuritaires, à s’autoréguler et l’absence de perturbation de la microflore normale font d’eux des outils potentiels pour le contrôle des bactéries pathogènes (Garcia et al., 2010).
Tout au long de cette section, des exemples de phages staphylococciques seront traités préférentiellement.
Historique des phages
Les phages ont été co-découverts par Frederick Twort, un bactériologiste anglais et Félix d’Hérelle, un microbiologiste Franco-canadien entre les années 1915 et 1917 (Twort, 1915). La première application rapportée de phages contre les maladies infectieuses humaines a pris place en 1921. Les phages ont été utilisés pour traiter les infections cutanées staphylococciques. Après 24 à 48 heures d’injection de la préparation phagique, les infections ont régressées et des patients ont été guéris (Sulakvelidze et al., 2001). Des produits phagiques dont le Bacté-staphy-phage ont été commercialisés en France (par la compagnie prenant aujourd’hui le nom de L’Oréal), aux États-Unis (par la compagnie Eli Lilly, Indianapolis) et surtout en Géorgie et en Pologne (Sulakvelidze et al., 2001). Cependant, avec l’introduction des antibiotiques dans les années 1940, les pays occidentaux ont adopté ces derniers comme agents antimicrobiens et ce n’est qu’après l’émergence de souches pathogènes résistantes aux antibiotiques, que l’intérêt et la recherche en phagothérapie ont repris leur cours aux États-Unis et dans plusieurs pays occidentaux, dont
15 l’Angleterre, les Pays-Bas, le Portugal, l’Israël, l’Australie, le Canada, etc. (Housby & Mann, 2009).
Classification des phages
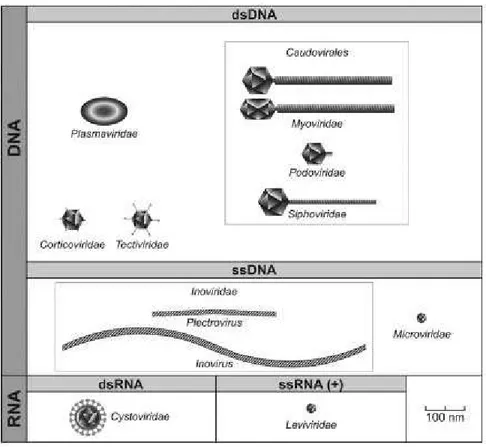
Les phages connus jusqu’à présent sont majoritairement classés sous l’ordre des
Caudovirales ou phages à queue et se divisent en trois familles : Myoviridae, Podoviridae
et Siphoviridae. Ces phages à ADN double brin diffèrent par leur morphologie et la taille de leur génome (Ackermann & Prangishvili, 2012). Les phages appartenant à la famille des
Myoviridae ont une longue queue contractile et la présence d’une forme de « cou » qui
sépare la capside de la queue. Avec une taille de génome supérieure à 120 000 paires de bases (pb), les myophages ont habituellement le plus large génome des trois familles. Les
Siphoviridae possèdent une longue queue non contractile et une capside icosaédrique ou
allongée, dépendamment du morphotype, ainsi qu’un génome d’environ 40 000 pb. Enfin, les Podoviridae ont une queue courte et une taille de génome plus petite que 20 000 pb (Ackermann & Prangishvili, 2012; King et al., 2011; Kwan et al., 2005) (Figure 1.2).
16
Figure 1.2. Les morphotypes des bactériophages. Figure adaptée de King et al. (2011). Les phages de S. aureus isolés jusqu’à présent sont nombreux et leurs génomes constituent une large section dans les bases de données publiques du NCBI. Une classification de ces phages a récemment été proposée et supplémente les travaux précédents d’autres équipes (Daniel et al., 2007; Deghorain & Van Melderen, 2012; Kwan
et al., 2005). Quatre-vingt-cinq génomes de phages et de prophages dont 15 spécifiques aux
staphylocoques à coagulase négative, ont été regroupés en se basant sur leurs similarités protéomiques respectives. Cette approche a donné lieu à 9 groupes distincts dont 7 groupes sont composés de phages appartenant à la famille des Siphoviridae (classe II), un groupe formé de Myoviridae (Classe III) et un dernier rassemblant les Podoviridae (classe I). Parmi les phages appartenant à la classe II, 2 groupes sont composés de phages de
S. aureus et de phages d’autres espèces de Staphylococcus, montrant une certaine proximité
17 Figure 1.3. Représentation du réseau entre les phages de staphylocoques basée sur le contenu protéomique. Les cercles représentent les 9 groupes différents. Les couleurs indiquent l’espèce de l’hôte (magenta: S. aureus; mauve: S. aureus et staphylocoques à coagulase négative; bleu: staphylocoques à coagulase négative). Le nombre de génomes est indiqué entre parenthèses. Le groupe 8 correspond à la classe I (Podoviridae), le groupe 7 correspond à la classe III (Myoviridae) et le reste, à la classe II (Siphoviridae). Les lignes grises entre chaque groupe sont indicatrices d’une identité d’au moins 30 % en protéines partagée entre deux génomes de phages et plus. Figure tirée de Deghorain & Van Melderen (2012).
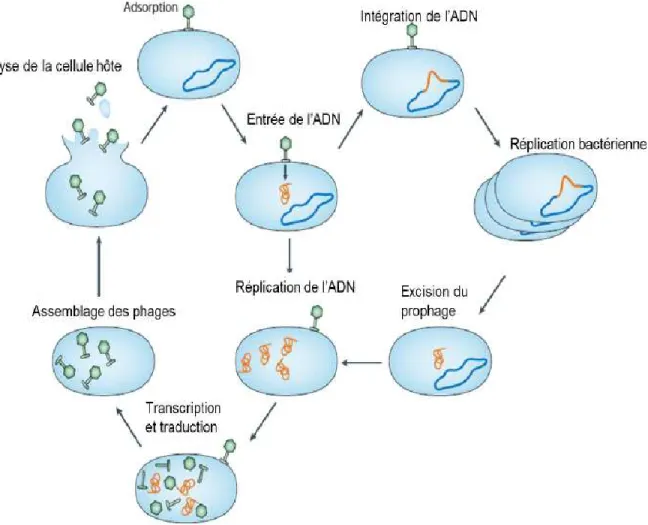
Cycles lytique et lysogénique
Les phages se divisent en deux groupes suivant le cycle adopté. Il existe des phages dits virulents et des phages tempérés. Les phages virulents suivent un cycle lytique qui s’achève par la lyse bactérienne et la libération de nouveaux virions dans le milieu allant infecter d’autres cellules. De l’autre côté, les phages tempérés suivent un cycle lysogénique en intégrant leur ADN dans le chromosome bactérien. Prenant le nom de prophages, ils se répliquent passivement avec leur hôte jusqu’à ce qu’une induction quelconque déclenche l’excision de leur matériel génétique et permette la continuation d’un cycle lytique résultant en la lyse bactérienne et la libération de nouveaux virions (Housby & Mann, 2009) (Figure 1.4).
18
Figure 1.4. Cycles lytiques (gauche) et lysogéniques (droite) des phages virulents et tempérés respectivement. La figure est adaptée et traduite de Labrie et al. (2010).
Les deux cycles lytique et lysogénique commencent par la reconnaissance et l’adsorption du phage à un récepteur bactérien spécifique. Quelques études ont tenté d’identifier les récepteurs bactériens et phagiques chez S. aureus. Toutefois, ceux-ci sont loin d’être confirmés. Une étude a pu démontrer que les acides téichoïques de S. aureus sont des polymères indispensables à l’interaction et l’adhésion de la bactérie aux cellules épithéliales nasales humaines (Weidenmaier & Peschel, 2008). Par ce fait, ces éléments pourraient jouer un certain rôle dans l’adhésion des phages à la surface bactérienne. Les acides téichoïques sont des éléments majeurs de la paroi bactérienne présents à la surface des bactéries à Gram positif. Ils se lient de façon covalente à la couche de peptidoglycanes constituant 50 % du poids de la paroi bactérienne (Lowy, 1998). Du côté phagique, une étude faite par Kaneko et al. (2009) a pu identifier un constituant de la structure du bout de la queue du phage staphylococcique phiSLT de classe II, groupe 4 (ORF636). Celui-ci
19 pourrait être à l’origine de la reconnaissance et de l’adhésion du phage à la chaîne de l’acide lipotéichoïque présent à la surface de S. aureus. Ce composant semble se lier spécifiquement à S. aureus et ne peut se lier à aucune autre espèce de staphylocoque ou bactérie à Gram positif (Kaneko et al., 2009). L’adsorption est l’une des étapes influençant le spectre d’action phagique et expliquant, en partie, la spécificité des phages à une espèce ou à une souche bactérienne donnée.
La seconde étape consiste en l’éjection de l’ADN dans la cellule hôte. Rashel et al. (2008) ont démontré qu’une hydrolase à peptidoglycanes située à l’extrémité de la queue du phage staphylococcique phiMR11, appartenant à la famille des Siphoviridae (classe II, groupe 2, selon Deghorain & Van Melderen, 2012), favorise l’entrée de l’ADN phagique dans le cytoplasme bactérien en formant des trous dans la couche de peptidoglycanes. Dans le cas des phages de la famille des Myoviridae, le tube central de la queue du phage est abaissé dans le pore formé jusqu’à ce qu’il ait un contact avec la membrane cytoplasmique et l’ADN est injecté (Lobocka et al., 2012). La capside protéique, dépourvue d’ADN, reste à l’extérieur de la cellule.
Une fois l’ADN phagique inséré, deux voies sont possibles selon les gènes disponibles : le mode lytique et le mode lysogénique.
Le mode lytique peut se poursuivre par la transcription des gènes de phages, précoces, intermédiaires et tardifs. Les gènes précoces détourne le métabolisme bactérien et initient la réplication d’ADN phagique. Les gènes intermédiaires permettent la synthèse et la réplication de l’ADN grâce à l’ADN polymérase cellulaire. Durant cette étape, de multiples copies du génome phagique sont formées, et ce, 3 à 8 minutes après l’infection. Enfin, les gènes tardifs, commençant environ 8 minutes après l’infection, codent pour les protéines structurales, l’encapsidation de l’ADN dans la capside phagique (procapside) ainsi que pour les protéines d’endolysine et d’holine (Ackermann, 1999).
L’encapsidation constitue la dernière étape dans la maturation de l’ADN des phages caudés et est opérée à l’aide d’une terminase. Cette dernière coupe l’ADN et le transporte dans la procapside. Il existe notamment deux modes principaux d’encapsidation selon le type de phage. Dans le cas d’un phage de type « pac », l’encapsidation achève lorsque la procapside est entièrement pleine. Le clivage de l’ADN par la terminase est alors produit à une position quelconque donnant lieu à des concatémères de tailles différentes et des
20
extrémités redondantes. Pour ce qui est des phages de type « cos » ou à extrémités cohésives, le site de clivage dépend d’un site « cos » reconnu par la terminase. Celle-ci coupe à la même position donnant des copies complètes du génome de phage (Ackermann, 1999).
Le cycle lytique prend fin lorsque la holine et l'endolysine perforent la paroi bactérienne et lyse la cellule hôte respectivement. La holine est une petite protéine qui cause des lésions ou des trous non spécifiques en se polymérisant dans la membrane bactérienne. Ceci permet à l'endolysine d’atteindre la couche de peptidoglycanes et de l’hydrolyser. Chacune de ces protéines a une cible bien définie dans la couche de peptidoglycanes. Une fois la lyse cellulaire réalisée, les nouveaux virions sont libérés dans l’environnement extracellulaire, et s’apprêtent à infecter d’autres cellules bactériennes. Le nombre de particules virales relarguées dépend de la souche de phage, de l’étape de croissance de la bactérie hôte lors de l'infection, des composants nutritifs entourant la cellule et de plusieurs facteurs environnementaux (Ackermann, 1999; Weinbauer, 2004; Wittebole et al., 2014).
Dans le cas d'un cycle lysogénique, et après l’étape d'entrée de l’ADN phagique dans le cytoplasme bactérien, l'ADN est intégré dans le chromosome bactérien. Ce processus se déroule grâce à un gène d’intégrase présent dans le génome phagique, à des sites d’attachement marquant l’emplacement favorable à l’insertion du génome phagique et à un gène répresseur empêchant la poursuite d’un cycle lytique. L'entité virale intégrée prend alors le nom de « prophage » et se réplique passivement avec le chromosome bactérien. Il est donc transféré verticalement avec le génome complet de son hôte à la progéniture de chaque cellule fille bactérienne (Weinbauer, 2004). Un stimulus comme les rayons UV, l’induction chimique ou thermale, et autres, peut causer l’excision du prophage et déclencher un cycle lytique conduisant à la libération de nouveaux virions dans l’environnement (Ackermann, 1999).
Critères de sélection des phages destinés à des fins de biocontrôle
Destiné aux utilisations à des fins médicales et alimentaires, un phage doit être bien caractérisé et passer par plusieurs étapes de validation définissant son statut d’agent de biocontrôle. La définition d’un produit phagique antimicrobien acceptable est un phage
21 strictement virulent, possédant un génome dépourvu de gènes de virulence, polyvalent (infectant plusieurs souches de l'espèce ciblée), efficace dans tout type de matrice, in vitro et in situ et maintenant une activité lytique stable à long terme. Il est à noter que ces critères ne sont pas énumérés par ordre d’importance.
Un phage strictement virulent
Un phage strictement virulent est incapable d’intégrer ou d’insérer son ADN dans celui de l’hôte bactérien, et ce, à cause de l’absence des fonctions génétiques requises pour l’intégration (Klumpp et al., 2008) ou de la présence de mutations ponctuelles affectant le module de lysogénie du phage (Garcia et al., 2009a).
Contrairement à ceux-ci, les phages tempérés peuvent s’intégrer dans le chromosome bactérien suite à l’entrée de l’ADN dans la cellule de l’hôte. Par ce fait, ils contribuent et favorisent le transfert horizontal du phénotype de virulence en transportant des gènes codant pour la production de toxines, entre autres, entre les populations bactériennes (Ackermann, 1999; Scharn et al., 2013).
Ce phénomène prend le nom de transduction et fait partie des trois méthodes employées (conjugaison et transformation étant les autres) par les bactéries pour transférer ou recevoir des gènes d’une cellule à une autre. Idéalement, ce matériel génétique exprimera la même fonction dans la cellule réceptrice comme dans la cellule donneuse (Kelly et al., 2009). L’avancement dans la technologie de séquençage de génomes bactériens complets et les comparaisons phylogénétiques ont permis de confirmer l’occurrence de ce phénomène (Thomas & Nielsen, 2005).
Une fois induit par un stress environnemental, le prophage est excisé et libéré du génome bactérien, transportant à l’occasion des gènes bactériens, et les transférant à la prochaine cellule hôte (Thomas & Nielsen, 2005). Cet échange génétique provoque une évolution, une diversification naturelle et, parfois, une virulence plus accentuée à la cellule hôte. En effet, des analyses bio-informatiques récentes de génomes de plusieurs souches différentes de S. aureus confirment le développement de la pathogénicité de cette espèce grâce à l’acquisition de gènes via des transferts horizontaux (Suzuki et al., 2012). Des observations de transfert d’îlots de pathogénicité mobiles (ou SaPI) contenant plusieurs facteurs de virulence (Novick, 2003), dont les gènes d’entérotoxines staphylococciques sea et see présents dans des prophages (Balaban & Rasooly, 2000) et des gènes conférant une
22
résistance aux antibiotiques (Mašlaňová et al., 2013) ont été notées chez S. aureus. Chen & Novick (2009) ont aussi observé un transfert d’îlots de pathogénicité entre une souche de S.
aureus et une souche de Listeria monocytogenes dans le lait cru à l’aide des phages 80α,
ɸ11, ɸNM2 et ɸNM4. Il suffit qu’une souche non pathogène ait acquis un gène de virulence par l’intermédiaire d’un prophage pour qu’elle constitue un danger à la santé publique (Skurnik et al., 2007).
Ces transferts horizontaux se font le plus souvent suite à une induction ou un stimulus. Il a été montré que les antibiotiques induisent la réponse SOS chez S. aureus résultant en la mobilisation de prophages et le transfert horizontal d’îlots de pathogénicité (Cirz et al., 2007; Maiques et al., 2006). Il est intéressant de noter qu’en général, certains phages transportent un seul facteur de virulence. Toutefois, il existe quelques exceptions, dont les siphophages phiN315 (classe II, groupe 6) et phiSa3 (classe II, groupe non identifié) qui encoderaient jusqu’à cinq facteurs de virulence (Malachowa & DeLeo, 2010).
Dans le cas des siphophages phi11 (classe II, groupe 1), phi80a (classe II, groupe 1) et phi80 (classe II, groupe 2) de S. aureus, leur capacité à transporter des gènes du chromosome bactérien, ainsi que des gènes retrouvés sur des éléments mobiles, est bien documentée (Mašlaňová et al., 2013). Un autre exemple de phage qui ne répond pas à ce critère de sélection est SA12, un phage tempéré de S. aureus appartenant à la famille des
Siphoviridae (classe II, groupe non identifié), qui a été isolé d’une infection clinique
staphylococcique (Chang et al., 2013). Le séquençage de son génome a permis la détection du gène d’intégrase intact. Ce phage est considéré posséder un élément favorisant le transfert horizontal de gènes de virulence (Chen & Novick, 2009).
Un phage ayant un génome dépourvu de tout facteur de virulence
Un agent de biocontrôle doit avoir un génome dépourvu de facteur de virulence. L’utilisation d’un phage encodant des gènes de virulence lors des applications médicale et alimentaire, provoquera sans doute le transfert de propriétés pathogènes aux bactéries de la microflore normale humaine. Ainsi, il constituerait un risque à la santé humaine. De ce fait, le génome d’un phage destiné au contrôle microbien devrait être séquencé afin d’assurer son innocuité et l’absence de gènes de virulence potentiels.
Des exemples de phages non utilisables en thérapie ont été rapportés dans quelques études. Ces phages possèdent des gènes de virulence dans leur génome encodant