HAL Id: hal-02822267
https://hal.inrae.fr/hal-02822267
Submitted on 6 Jun 2020HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Phéromones royales et régulations sociales chez l’abeille
Apis mellifera
Geoffrey Montes
To cite this version:
Geoffrey Montes. Phéromones royales et régulations sociales chez l’abeille Apis mellifera. [Stage] Université Jean Monnet (Saint-Etienne) (UJM), Saint-Etienne, FRA. 2009, 50 p. �hal-02822267�
Phéromones royales et régulations
sociales chez l’Abeille
Apis mellifera
(Source: http://www.thehoneygatherers.com/)
présenté par Geoffrey MONTES
MASTER 1
Sciences, Technologies, Santé ; MENTION Ecologie et Ethologie 2008/2009
Tuteur de stage : Jean-Louis BERTHON
Enseignant - Chercheur Université Jean-Monnet
Faculté des Sciences et Techniques
23, rue Dr Paul Michelon 42023 Saint-Etienne Cedex 2 Tél. : +33 (0)4 77 48 15 00
Maître de stage : Yves LE CONTE
Chargé de Recherche
INRA – UMR 406 Abeilles et Environnement
Laboratoire Biologie et Protection de l’abeille Site Agroparc - Domaine Saint-Paul
Ce qui distingue d’emblée le pire architecte de l’abeille la plus experte, c’est qu’il a construit la cellule dans sa tête avant de la construire dans la ruche.
Avant-propos
Dans un premier temps, je remercie Luc BELZUNCES, directeur de l’unité Abeilles et Environnement, pour m’avoir accueilli dans son unité.
Je tiens à remercier profondément mon maître de stage, Yves LE CONTE, pour la confiance qu’il m’a accordé en me faisant intégrer son équipe. Il m’a fait découvrir ce monde merveilleux qu’est celui de l’abeille. Je le remercie pour tous ses conseils et son entrain, qui ont permis à ce stage de se dérouler dans des conditions optimales.
Je remercie plus particulièrement Alban MAISONNASSE qui m’a encadré durant toute la durée de mon stage. Je tiens à le remercier pour sa disponibilité et son écoute, pour m’avoir guidé dans mon travail. Mon séjour dans ce laboratoire à été une expérience enrichissante et c’est en grande partie grâce à lui.
Je remercie chaleureusement toute l’équipe du laboratoire, Jean-Marc BECARD pour son aide en informatique et ses blagues à n’en plus finir, Didier CRAUSER pour sa passion pour l’apiculture et notre périple lors de la visite des ruchers.
C’est avec une grande sincérité que je remercie une autre personne du laboratoire, Dominique BESLAY, pour l’aide qu’elle aura apporté à mes manips ainsi que pour sa gentillesse et nos bons moments passés ensemble.
Je n’oublie évidemment pas de remercier les autres stagiaires qui étaient à mes cotés au cours de mon stage Horiya AMAACH, Claudia DUSSAUBAT, Cédric ALAUX, François DUCLOZ, Vincent MARTEAU, Claire et Martin LE CONTE pour m’avoir apportés leur aide dans l’une de mes manips, nos deux américains adorés Hunter BRATTON et Daniel HERLIHY, et plus particulièrement Aurélie BALDY pour m’avoir encouragé et permis de passer un merveilleux séjour à Avignon !
Sommaire
Avant-propos ... 3
Sommaire ... 4
Table des illustrations et tableaux ... 6
Introduction ... 7
Partie I : Une société très structurée... 7
I/- Colonie, castes et reproduction chez l’abeille domestique (Apis mellifera)... 7
1) Structure eusociale ... 7
2) Les différentes castes... 8
3) Une reproduction particulière ... 9
II/- Communication ... 9
Partie II : Phéromones royales et régulation de la colonie ... 11
I/ La communication chimique ... 11
1) Généralités sur les phéromones ... 11
2) La phéromone royale des glandes mandibulaires ou QMP ... 13
3) Les phéromones royales ... 13
II/- Phéromones royales incitatrices ... 14
1) Effet incitateur stimulateur des phéromones royales : la construction des rayons de cire ... 14
2) Effet incitateur stimulateur des phéromones royales : le phénomène de cour .... 16
III/- Phéromones royales modificatrices ... 17
1) Effet modificateur inhibiteur des phéromones royales : le développement ovarien ... 17
Matériel & Méthodes ... 19
I/- Reines démandibulées ... 19
1) L’élevage des reines ... 19
2) La démandibulation... 22
3) Mise en cagette... 23
II/ Comportement de cour ... 25
III/ Le développement ovarien ... 27
1) Dissection et évaluation des ovaires ... 27
2) Analyses statistiques... 28
IV/ Extraction du 9-ODA ... 28
Résultats ... 29
1) La construction des rayons ... 29
2) Le phénomène de cour ... 30
3) Le développement ovarien des ouvrières ... 31
4) Extraction du 9-ODA ... 32
Discussion ... 33
1. Le choix du format des cellules lors de la construction ... 33
2. Le cortège royal ... 35 3. L’inhibition ovarienne... 36 Bibliographie ... 38 Annexe 1 ... 42 Annexe 2 ... 43 Annexe 3 ... 44 Annexe 4 ... 45 Annexe 5 ... 46 Annexe 6 ... 47 Annexe 7 ... 48 Annexe 8 ... 49
Table des illustrations et tableaux
Figure 1: Biologie de l'abeille et distinction en trois castes ... 8
Figure 2: Rayon d'une ruche... 15
Figure 3: Schéma représentant l'organisation d'une ruche éleveuse de reines ... 19
Figure 4: Cadre de couvain ouvert de moins de trois jours... 21
Figure 5: Larves de moins de trois jours plongées dans de la gelée royale diluée avec de l'eau ... 21
Figure 6: Cellules royales sur une barrette... 21
Figure 7: Organisation d'une boite où est achevé l'élevage de la reine ... 22
Figure 8: Démandibulation d'une reine ... 23
Figure 9: Cagette pain pendant la construction du rayon ... 24
Figure 10: La ruche vitrée ... 26
Figure 11: Variation du diamètre moyen des cellules entre les différents traitements, toutes séries confondues ...29
Figure 12: Nombre de cellules moyen des cellules entre les différents traitements, toutes séries confondues ...29
Figure 13: Score moyen du développement ovarien pour la série I ... 31
Figure 14:Score moyen du développement ovarien pour la série II ... 31
Figure 15: Score moyen du développement ovarien pour la série III ... 31
Tableau 1: Tableau récapitulatif des différents types de sociétés et de leurs caractéristiques ... 7
Tableau 2: Tableau récapitulatif des expériences de Ledoux et al. (2001)... 16
Introduction
Partie I : Une société très structurée
I/- Colonie, castes et reproduction chez l’abeille domestique (Apis mellifera)
Les abeilles à miel appartiennent toutes à un seul genre, le genre Apis (Linné, 1758), constitué seulement de 7 espèces, celui-ci étant extrêmement répandu dans le monde. Apis
mellifera, qui fait l’objet de notre étude, correspond à l’abeille mellifère (ou abeille dite
domestique) utilisée pour ses productions de miel, de cire ou de gelée royale en apiculture ainsi que pour ses qualités de pollinisateur. Il existe plusieurs races géographiques de cette espèce comme A. m. mellifera L. (abeille noire endémique de France) ou A. m. lingustica S. (abeille italienne) (Le Conte, 2006). Aucune distinction entre les différentes races de l’espèce
Apis mellifera ne sera faite par la suite, et le terme abeille désignera l’abeille mellifère.
1) Structure eusociale
Les sociétés complexes d’abeilles constituent un défi formidable pour les écologistes évolutifs, principalement en raison de la présence de castes stériles (Brockman 1984). Une des classifications proposée par Crespi et Yanega en 1995 a permis de reconnaitre cinq systèmes sociaux distincts, qui correspond à autant de niveaux de complexification de l’évolution des comportement sociaux (Tableau 1). Les individus des systèmes eusociaux, comme chez les abeilles domestiques du genre Apis, présentent des spécialisations qui sont irréversibles (Crespi et Yanega, 1995) :
• La présence d’individus sexués et asexués morphologiquement différents
• L’élevage en commun de la progéniture
• La division du travail (Michener, 1974)
• La coexistence d’au moins deux générations successives (Wilson, 1971)
Tableau 1: Tableau récapitulatif des différents types de sociétés et de leurs caractéristiques (d’après Crespi et Choe, 1997)
Type de société Soin parental
Site de reproduction partagé Coopération à l'élevage Soin alloparental Castes Subsocial + Colonial - + Communautaire + + + Elevage coopératif + + + +
2) Les différentes castes
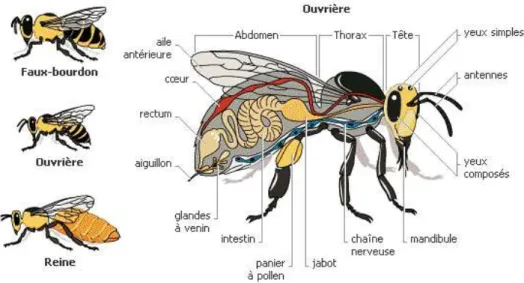
Les sociétés d’abeilles, composées de 30.000 à 80.000 individus, sont organisées en trois castes à la morphologie (Fig. 1) et aux rôles bien distincts qui accomplissent chacun un travail spécifique au sein de la colonie.
Figure 1: Biologie de l'abeille et distinction en trois castes (Source: http://apiculture.virginradioblog.fr/)
Les mâles (ou faux-bourdons) ont un rôle strictement limité à la fécondation des jeunes reines pendant le vol nuptial. Ceux qui parviennent à féconder la reine provoquent la déchirure de leur bulbe et de leurs plaques chitineuses. Ils meurent ainsi peu de temps après, suite à la perte de leurs organes dévaginés. Les autres mâles, quant à eux, seront tués par la colonie et disparaîtront dès la fin de l’été, abandonnés par le reste de la colonie.
Les femelles sont divisées en une caste d’ouvrière non reproductrice et une caste reproductrice représentée par une unique reine. La reine est le seul individu fertile et assure exclusivement la ponte et le maintient de la cohésion de la société. C’est l’élément central de la ruche. La progéniture non reproductrice de la reine, les ouvrières, apporte pour sa part nourriture et soins divers au couvain et à la reine, ceci par un partage des taches qui diffère en fonction de leur âge. Ce phénomène est plus communément appelé polyéthisme lié à l’âge. Deux stades peuvent être distingués : le stade nourrices où elles exécutent des taches d’intérieurs dites sans risques (nettoyage des cellules, operculation, stockage du nectar, construction des rayons), et plus tard dans leur vie, des taches extérieures dites risquées (gardiennage, butinage). En effet, en étant à l’intérieur de la ruche, l’abeille est protégée par la
structure du nid (artificiellement, il s’agit de la ruche) ainsi que par la colonie tout entière, alors qu’à l’extérieur l’abeille qui accompli sa tâche est soumise à son environnement (prédations, etc.). Il existe par ailleurs une grande plasticité de développement comportemental. Ainsi, la succession de ces taches n’est pas figée mais peut, au contraire, être réversible ou accéléré suivant les besoins de la colonie et des changements de l’environnement social (Huang et Robinson, 1996). Chez les abeilles, la division des tâches dans la colonie est bien connue, mais des recherches sont en cours sur son mode de fonctionnement.
3) Une reproduction particulière
Les abeilles peuvent se reproduire par parthénogénèse, celle-ci se définie comme le développement d’un ovule qui n’a pas été fécondé. En effet, ce sont les mâles qui naissent d’œufs non fécondés et qui sont par conséquent haploïdes ce qui mène à l’appellation de parthénogénèse arrhénotoque.
Au contraire, les œufs fécondés par la reine donnent naissance à des femelles, reine ou ouvrière, et sont diploïdes. La différence entre reine et ouvrière provient uniquement de la nourriture apportée par les nourrices aux larves à partir du 3ème jour de développement. En effet, dès le 3ème jour de la gelée royale excrétée à partir des glandes hypopharyngiennes (ou glandes nourricières) et des glandes mandibulaires des ouvrières est donnée exclusivement aux larves contenues dans les cellules royales (Mc Cleskey et Melampy, 1939), alors que les mâles et les ouvrières reçoivent une bouillie larvaire1. C’est cette unique différence qui opère la distinction entre femelle sexuée et non sexuée, indifférenciées au départ.
II/- Communication
La régulation de la division temporelle du travail ainsi que le haut niveau d’organisation de la colonie est basée sur une communication efficace entre tous les individus de la ruche. Cette communication est basée sur des messages sonores (bruissement de la colonie), tactiles, et chimiques. Il est connu une communication comportementale comme la
1
Bouillie larvaire : il s’agit d’un mélange de pollen et de miel qui apporte de l’eau aux larves à hauteur de 66% en moyenne (Haydack, 1968). Cette nourriture est reçue par bouche à bouche avec leurs nourrices et celles-ci rejetteront leurs excès d’eau par leurs glandes salivaire. Les nourrices ont besoin de cette salive, à base d’enzymes appelées diastases, qu’elles lèchent pour digérer efficacement le pollen, car leur appareil digestif
communication par la danse, langage d’une extrême précision. Décrite par Karl von Frisch en 1967, la danse en huit ou danse frétillante apporte des informations sur la direction, la distance, et la qualité des ressources disponibles. Il existe cependant d’autres types de communication comme la communication par contact antennaires. Chez tous les insectes sociaux, comme chez l’abeille Apis mellifera, la communication chimique tient également une part très importante dans le fonctionnement et la régulation des relations entre les individus que cette société compose.
Partie II : Phéromones royales et régulation de la colonie
I/ La communication chimique
1) Généralités sur les phéromones
Communiquer est vital pour survivre et se reproduire pour les animaux qui utilisent leurs sens pour percevoir leur environnement. L’abeille communique tout aussi bien par des signaux sonores et visuels que chimiques. Elles utilisent des odeurs perçues par olfaction et des molécules non volatiles perçues par gustation ou par contact comme information. L’odorat des abeilles est très spécialisé. Cette communication chimique est basée sur la production et la réception de phéromones (mot inventé par Karlston & Lüber en 1959) qui sont des molécules volatiles et ont une action intraspécifique. On peut les définir comme substance chimique émise par un ou plusieurs individus, ce qui déclenche une modification comportementale spécifique ou physiologique chez un individu de la même espèce. Une phéromone peut être formée d’un mélange de quelques composés chimiques, appelé « bouquet phéromonal » en proportion stable pour une espèce ou une population. Le sens du message phéromonal et les comportements induits reposent sur la composition particulière du mélange. Ce message est perçu par les récepteurs antennaires et agit directement sur le système nerveux. Cette détection est possible grâce aux sensilles, des neurones récepteurs olfactifs contenus dans des soies présentes en très grand nombre sur l’antenne.
Plusieurs types de phéromones peuvent être distingués :
• Les phéromones qui provoquent une réaction comportementale rapide et réversible dès réception de la substance sont dites incitatrices (Releaser pheromones) (Wilson et Bosser, 1963). Il s’agit principalement de phéromones sexuelles qui orientent le processus d’accouplement, d’autres sont quant à elles impliquées dans les processus d’alarme et de défense, d’orientation, d’oviposition, d’agrégation, d’intégration des activités de la colonie...
• Il existe également des phéromones modificatrices (Primer pheromones) qui entraînent un changement physiologique des individus qui les reçoivent, avec un effet différé (Wilson et Bosser, 1963). Elles ont pour objectif de coordonner le développement physiologique et comportemental d’un ensemble d’individus. Du fait de leur action à plus
long terme, elles sont relativement difficiles à identifier. Il s’agit principalement de phéromones de castes, de grégarisation, etc.
A ces deux types de phéromones se sont ajouté deux autres catégories :
• Les phéromones coloniales (Colony pheromones) sont issues de multiples castes et de différents stades de développement de l’abeille. Elles peuvent avoir une fonction incitatrice ou modificatrice (Slessor et al., 2005)
• Les phéromones opportunistes (Passenger pheromones) ont, elles, un rôle de modification comportementale ou physiologique sur l’individu qui reçoit cette substance après transfert entre les insectes. Lors de l’échange, le complexe phéromonal reste inactif mais devient actif sur l’individu cible (Slessor et al., 2005).
La découverte de la phéromone royale (Barbier et Lederer, 1960) a permis d’introduire l’idée que la communication chimique est prépondérante dans le langage de l’abeille, lançant un véritable engouement scientifique. Ce concept, avéré exact, a permis la découverte de nouvelles phéromones royales (Slessor et al., 1988 ; Keeling et al., 2003), de couvain (Le Conte et al., 1990), de butineuses (Leoncini et al., 2004),..., qui permettent une meilleure compréhension des sociétés d’abeilles. Les comportements et les mécanismes de la communication chimique sont aussi variés que les molécules elles-mêmes. Un insecte comme l’abeille est une véritable usine chimique émettant à la fois des esters, des acides, des alcools, des hydrocarbures...
En utilisant le même langage, une colonie d’Apis mellifera peut coordonner la vie de plusieurs dizaines de milliers d’individus. Cependant, la communication chimique reste extrêmement complexe. Certaines molécules ont été découvertes, d’autres sont en cours d’identification et leurs effets sur le comportement de l’animal n’est pas toujours connu. Nos études se sont portées principalement sur les phéromones royales de l’abeille et l’effet qu’elles peuvent avoir sur le plan comportemental et morphologique. La reine, véritable sécrétrice de phéromones, réalise ses principales interactions avec les ouvrières par le biais de messages chimiques, et ceci afin de maintenir la cohésion et l’efficacité de la colonie...
2) La phéromone royale des glandes mandibulaires ou QMP
La principale phéromone produite par la reine est un paradoxe puisqu’elle agit à la fois et ceci de façon immédiate sur le comportement puis sur la physiologie de façon retardée (Butler, 1959). La nature chimique de cette phéromone, connue depuis 1969 (Barbier et Lederer, 1960) en fait l’une des toutes premières découvertes chez les insectes. Uniquement une molécule avait été identifiée à cette époque, le 9-ODA (acide 9-oxo-(E)-2-décènoïque). Aujourd’hui, on sait qu’il s’agit d’un bouquet phéromonal, appelé QMP (Queen Mandibular
Pheromone), comprenant 5 molécules (trois composés acides, deux composés aromatiques)
(Slessor, 1988). Le mélange est constitué de trois composés acides : acide 9-oxo-(E)-2-décènoïque (ou 9-ODA), acide (R)-9-hydroxy-2(E)-9-oxo-(E)-2-décènoïque (ou R-9-HDA) et acide (S)-9-hydroxy-2(E)-décènoïque (ou S-9-HDA) ; et des deux aromatiques : parahydroxybenzoate de méthyle (ou HOB) et 4-hydroxy-3-méthoxyprophényléthanol (ou HVA) (Annexe 1A). Ces molécules agissent en synergie, chacun de ces composés pris isolément n’ayant aucune activité, mis à part le 9-ODA qui seul a une activité très faible. Du point de vue qualitatif, la fraction acide de cette QMP est plus importante.
Les glandes mandibulaires d’une reine s’étant accouplée, et ayant pondue des œufs comprend en moyenne 200µg de 9-ODA, 56µg de R-9-HDA, 24µ g de S-9-HDA, 20µ g de HOB et 20µ g de HVA (Slessor et al., 1988). Ces quantités décrites correspondent à un équivalent reine (Qéq), et aux quantités de molécules produites par les glandes mandibulaires royales en 24h (Naumann et al., 1991) (Annexe 1A).
3) Les phéromones royales
Keeling et al. ont identifié en 2003 quatre autres composés produits par la reine et qui fonctionneraient en synergie avec les QMP dans le but d’attirer les ouvrières à elle pour former la cour autour de la reine. Ces molécules, ensembles, correspondent à une autre phéromone royale appelée QRP (Queen Retinue Pheromone). La QRP est donc un mélange des cinq composés de la QMP et des quatre composés suivant : le (9Z)-octadec-9-ènoate de méthyle (Oléate de méthyle ou MO), le (E)-3-(4-hydroxy-3-méthoxyphényle)-prop-2-en-1-ol (alcool conyférilique), l’hexadecan-1-ol (alcool de cétyle), l’acide octadeca-9,12,15-triènoïque (acide linolénique) (Annexe 1B).
La transmission de la phéromone royale résulte essentiellement d’une contamination des ouvrières de la cour royale qui lèche et antenne la reine. La phéromone circule ensuite d’une ouvrière à une autre par échange de nourriture (trophallaxie), contact entre les individus, et volatilité (Naumann, 1991). Les phéromones royales, particulièrement les QMP, jouent sur de nombreux aspect du comportement et de la physiologie, comme l’essaimage, l’attirance des mâles (Gary, 1962), le format de la construction (Ledoux, 2001), le comportement de cour (Slessor, 1988), le développement des ovaires (Hoover, 2003), et d‘autres fonctions de la colonie. En revanche, s’il a été reconnu que les QMP ont une influence sur ces fonctions, on ignore encore si d’autres molécules royales peuvent jouer un rôle sur ces comportements ou sur cette inhibition des ovaires. Des études préliminaires ont prouvées qu’une reine démandibulée (sans mandibules et glandes mandibulaires) permettait de supprimer toutes traces de phéromones royales dans le corps de celle-ci. L’objectif de notre étude est d’étudier le format de la construction du nid, le phénomène de cour et le développement ovarien en absence des QMP afin de savoir si d’autres paramètres, peut être phéromonaux, sont impliqués dans ces mécanismes ou non.
II/- Phéromones royales incitatrices
1) Effet incitateur stimulateur des phéromones royales : la construction des rayons de cire
L’homme a toujours été fasciné par la régularité et la précision du travail des abeilles. La construction des rayons est un processus essentiel afin de mettre à disposition les cellules dans lesquelles la reine pourra pondre et assurer la pérennité de la colonie, les ouvrières accomplissant, quant à elles, toutes les autres fonctions nécessaires au bon développement et à la survie de la société. La cire utilisée pour construire les rayons est produite par les ouvrières à leur 15ème jour en moyenne et ceci à partir de leurs glandes cirières situées entre le 4ème et le 7ème sternite (4 paires). La cire est émise sous forme de petites écailles, auxquelles l’abeille ajoute de la salive et qu’elle malaxe avec ses mandibules afin de construire des cellules hexagonales très régulières. Le rayon peut comprendre 2 types de cellules, le premier au format « ouvrière », et l’autre au format « mâle ». Les cellules de type ouvrières sont localisées au centre du rayon (artificiellement, le cadre), alors que les cellules de type mâle se localisent en périphérie (Fig. 2). Le diamètre des cellules au format mâle est plus grand que
Cellule de mâle Cellules d’ouvrières Miel
celui des cellules au format ouvrière (x = 6,64 pour les mâles et x = 5,56 pour les femelles d’après Ledoux, 2001). En condition normale, les cellules au format ouvrière sont plus nombreuses que les cellules au format mâle (Hepburn, 1986 et Winston, 1987). Cependant, les mâles peuvent aussi être élevés dans les cellules au format ouvrière, mais ils sont, une fois adulte, plus petits et moins fertiles (Ribbands, 1953).
Figure 2: Rayon d'une ruche (Source: M. LE CONTE)
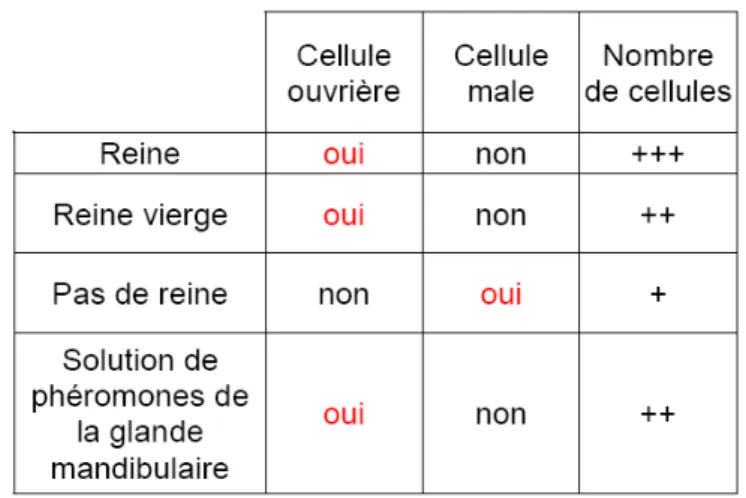
Le choix du type de cellules construit est une décision capitale et aura des conséquences dans l’histoire de vie de la colonie. La construction des cellules en présence royale a beaucoup été étudiée, et il a été constaté qu’un grand nombre de facteurs démographiques peut déterminer quand et comment les rayons mâles des colonies sont construis (Lee et Winston, 1985, 1987 ; Pratt, 1998). Cependant, d’autres études montrent par ailleurs que la reine elle-même a un effet stimulateur sur la quantité de cadres construits et le type de cellule, particulièrement grâce a ses phéromones (Darchen, 1957). L’absence de phéromone royale dans des ruches orphelines peut avoir des effets profonds sur l’étendue et la nature de la construction des rayons, construisant des cellules format mâles et intermédiaires (Taber et Owens, 1970). Les expériences de Ledoux en 2001 ont montrées que les glandes mandibulaires de la reine jouent un rôle dans la forme de construction des cellules. En effet, si il y a présence des molécules constituant la QMP ou d’une reine, il y a construction en format ouvrière. En revanche en l’absence de la reine ou de la phéromone mandibulaire, il y a construction en format de mâle (Tableau 2).
Tableau 2: Tableau récapitulatif des expériences de Ledoux et al. (2001) (Source : Geoffrey MONTES)
L’objectif de cette étude est, dans un premier temps, d’examiner l’influence de la reine et de son signal phéromonal sur la construction des rayons en élevant des mini colonies d’abeilles avec et sans reines. Plus exactement, nous avons comparé le type de cellule construit ainsi que la quantité de cellules au format ouvrière et mâle dans des mini colonies d’abeilles en présence soit d’une reine normale, soit avec une reine ayant subie une démandibulation (retrait des mandibules et de la glande mandibulaire), soit sans reine donc aucune phéromone royale.
2) Effet incitateur stimulateur des phéromones royales : le phénomène de cour La reine, assurant ponte et bonne organisation de la ruche, sort rarement de la ruche (sauf pour deux occasions : le vol nuptial et l’essaimage). En effet, ses ailes courtes, ses pattes longues dépourvues « d’outils » (contrairement aux ouvrières qui possèdent des outils spécifiques au pollen) ainsi que sa langue trop courte ne lui permettent pas d’aller butiner. Ainsi, la présence de la reine et son attractivité se manifeste par une attention particulière donnée par les ouvrières à celle-ci à l’intérieur du nid. Quand la reine est immobile sur un rayon, elle est rapidement encerclée par des ouvrières (gardiennes et jeunes ouvrières) qui se positionnent face à elle, lui offrant de la nourriture par trophallaxie, la palpant avec leurs antennes, la léchant.
Les expériences de Slessor et al. (1988) ont permis d’étudier le comportement de cour des ouvrières en fonction de différents traitements. La substance produite par les glandes mandibulaires est importante pour le phénomène de cour chez les abeilles à miel, Apis
L’observation in vivo de reine démandibulées à l’intérieur même de la ruche permettra de voir si une escorte est présente ou non, et d’en observer le nombre. L’objectif de notre étude est ici aussi de découvrir si les phéromones mandibulaires sont indispensables à ce phénomène et si d’autres molécules y sont impliquées.
III/- Phéromones royales modificatrices
1) Effet modificateur inhibiteur des phéromones royales : le développement ovarien
La répartition des tâches de reproduction est l’une des caractéristiques primaires chez les sociétés eusociales d’insectes. La différenciation des castes ouvrières / reines s’opère à l’état larvaire, par le nourrissage distinct des larves qui entraîne l’atrophie des ovaires des ouvrières (Oertel, 1930), avec la mort progressive des cellules entre le 4ème et 5ème stade larvaire. L’hormone juvénile2 (JH) et les ecdystéroïdes3 sont les principaux agents de cette différenciation.
Chez les abeilles, l’appareil reproducteur des femelles comporte une paire d’ovaires de type méroïstique polytrophe, ce qui indique que les cellules vitellines sont intercalées par groupes entre les ovules (Source : encyclopedie-universelle.com). Cependant, chez les abeilles sociales Apis mellifera, le développement de la gonade ou ovaire des ouvrières, n’est pas du tout le même que celui de la reine, qui est la seule féconde. La morphologie de la reine est adaptée à sa reproduction : elle présente un abdomen très développé notamment en période de reproduction ; deux ovaires hypertrophiées occupant une grande partie du volume de l’abdomen, reliés à la chambre vaginale par un oviducte ; une spermathèque, caractéristique fondamentale de la reine, qui va recueillir le sperme des différents mâles qui vont s’accoupler avec elle lors du vol nuptial. Le développement de la gonade femelle des ouvrières n’est pas du tout identique à celui de la reine : il est atrophié.
2 Hormone juvénile : appartenant à la classe de sesquiterpènes, le taux de JH permet de contrôler la mue dans la
période immature de l’animal et la reproduction, au stade adulte de nombreux insectes. L’hormone juvénile est 2 fois plus élevée chez les prénymphes qui vont évoluer en reines que chez celles qui deviendront des ouvrières.
post-Chez les abeilles domestiques Apis mellifera, ce sont la reine et le couvain qui régulent le développement ovarien (Kropacova, 1969). Les productions phéromonales de la reine sont particulièrement importantes en ce qui concerne l’inhibition du développement ovarien des ouvrières. L’acide 9-oxo-(E)-2-décènoïque (9-ODA), composant majeur des glandes mandibulaires royales, a été la première molécule suggérée comme étant la principale phéromone modificatrice régulant le développement ovarien.
Les expériences de Hoover (2003) ont démontrés que la QMP (mélange de 5 molécules dont le 9-ODA) est responsable de l’inhibition du développement des ovaires des ouvrières. La dissection d’ouvrières ayant été en présence d’une reine démandibulée, c’est-à-dire incapable de produire la QMP, permettra de savoir si cette phéromone mandibulaire royale est la seule substance permettant le contrôle du développement des gonades femelles chez les ouvrières, ou si d’autres molécules sont impliquées dans cette fonction.
Matériel & Méthodes
Différents traitements ont été testés : 1) avec une reine vierge « T+ », 2) avec une reine démandibulée « Dém » (retrait des deux mandibules4 et des deux glandes mandibulaires), 3) sans présence de reine, sans phéromones « T- ». Il y a eu plusieurs essais par traitement et par répétitions, ainsi que trois répétitions. Il a été démontré dans des études préliminaires que la démandibulation permet la disparition de la QMP dans tout le corps de la reine.
I/- Reines démandibulées
1) L’élevage des reines
Des ruches éleveuses sont mises en place préalablement à chaque essai. Elles sont constituées de deux parties bien distinctes. La première correspond à une ruche classique avec la présence d’une reine qui régule la plupart des phénomènes de la société. La seconde partie est séparée de la précédente par une grille qui limite le transfert de la reine et de la phéromone royale (Fig. 3). Elle est dite orpheline et ne comprend donc pas de reine. En effet, c’est ici que les cellules royales seront constituées. L’absence de la reine permet l’élevage de nouvelles reines, la reine détruisant les cellules royales nouvellement constituées dans une ruche classique.
4
Grille limitant le transfert de la phéromone royale
Cadre de miel et de pollen
Partie avec reine Partie orpheline
Cadre de couvain ouvert
Cadre d’élevage à deux barrettes
Figure 3: Schéma représentant l'organisation d'une ruche éléveuse de reines (Source : Geoffrey MONTES)
Les alvéoles (cellules) d’élevage sont différentes. Pour les ouvrières, il s’agit d’une quelconque cellule dans un rayon, qui aurait pu aussi bien contenir du miel ou du pollen, alors que la reine sera élevée dans une cellule spécialement bâtie à son attention. La cellule royale est plus grande et un peu plus large, et ressemble à un « cocon de cire » accroché dans un coin de rayon. Par ailleurs, la nourriture que vont consommer les larves est différente, les ouvrières étant nourries à la bouillie larvaire et les reines à la gelée royale.
Deux barrettes, comprenant chacune 50 cupules, sont déposées dans la partie orpheline de la ruche éleveuse (H0). Les cupules sont des alvéoles construites artisanalement à partir de cire dans lesquelles les reines seront élevées. Le fait d’introduire au préalable ces barrettes permet aux cupules d’être mordillées par les ouvrières et de les imprégner de leurs odeurs (Annexe 2A). Cette manipulation permettra une meilleure prise en charge des futures larves. Du sirop, constitué d’eau chaude et de sucre, est fournie à la ruche pour stimuler les abeilles et les attirer du côté orphelin.
En fin de journée, un cadre de couvain ouvert de moins de trois jours est ensuite prélevé (H3). Le cadre correspond à des alvéoles non operculées comprenant des larves de stade larvaire inférieur à L3 (3ème jour du stade larvaire) (Fig. 4). Cette information permettra de prédire le jour d’émergence des reines (Annexe 2B)
Les barrettes et leurs cupules sont retirées juste avant le greffage les larves ainsi que le cadre de couvain. De la gelée royale est déposée préalablement et diluée avec de l’eau (Fig. 5) à l’intérieur de chaque cupule afin de permettre aux reines de se développer. En effet, dans la nature, les abeilles choisissent au hasard quelques cellules (de moins de trois jours) à qui elles donnent une nourriture spéciale appelée gelée royale. Il s’agit d’un produit blanchâtre, gélatineux et à saveur un peu acide que les nourrices fabriquent grâce à des glandes qu’elles possèdent dans la tête. Cette nourriture tonifiante offre la possibilité de transformer n’importe quelle larve d’abeille en reine potentielle. Le greffage se réalise à l’aide d’un « picking » : les larves sont prélevées délicatement dans leurs cellules et sont déposées dans les cupules artificielles (Annexe 2C). Une fois les larves greffées, les cadres de couvain et les barrettes sont reposés dans la ruche éleveuse pendant 7 jours. Les ouvrières vont ensuite élever les larves contenues dans les cellules royales.
Larve de moins de 2 jours environs, idéale pour le greffage Œuf Gelée royale diluée avec de l’eau
Figure 4: Cadre de couvain ouvert de moins de trois jours (Source : zebulon1er.free.fr) Figure 5: Larves de moins de trois jours plongées dans de la gelée royale diluée avec de l'eau
(Source: apiculture-populaire.com)
Une semaine plus tard, le cadre porteur des barrettes et de leurs cupules est retiré délicatement (Fig. 6). Puis, en laboratoire, les cellules royales sont séparées les unes des autres et sont placées chacune dans des boites contenant une dizaine d’abeilles appartenant à la ruche (Fig. 7). Ces dernières finiront l’élevage des reines jusqu’à leur émergence.
Figure 6: Cellules royales sur une barrette (Source: Alban MAISONNASSE)
Ces boites contiennent du candy5 et de l’eau pour nourrir les abeilles qui alimentent ensuite la reine par trophallaxie. Les boites sont placées à l’étuve à 34°C en présence d’un récipient rempli d’eau permettant un bon taux d’hygrométrie. Les boites sont entretenues régulièrement pendant une période de 10 jours (changement du papier buvard, eau et ajout de candy si nécessaire) jusqu’à l’émergence des reines (Fig. 7).
Abreuvoir Ancienne cellule royale
Figure 7: Organisation d'une boite où est achevé l'élevage de la reine (Source : Geoffrey MONTES)
2) La démandibulation
Après 17 jours, sont sélectionnées uniquement les boites contenant une reine viable. Parmi elles, environs les 2/3 seront démandibulées, car la démandibulation est une opération délicate qui peut engendrer une mortalité.
Les boites comprenant les reines qui seront démandibulées sont alors soumises au CO2,
permettant une anesthésie préalable. La manipulation consiste dans un premier temps à prélever délicatement la reine avec des gants. Elle est maintenue sur le dos entre le pouce et l’index de façon à dégager la tête (Fig. 8a). Ensuite, sous loupe binoculaire, une mandibule est saisie sous l’articulation par l’intermédiaire d’une pince fine. Par un geste délicat, en tournant la pince, la mandibule doit s’arracher laissant apparaitre un trou (Fig. 8b). Il faut ensuite saisir dans le trou créé la glande mandibulaire se trouvant à l’intérieur de la tête, celle-ci ressemblant à un gros sac de couleur blanchâtre (Fig. 8c). La glande est placée dans de l’isohexane afin de permettre sa conservation (Fig. 8d).
A la fin de la manipulation, du miel est apposé sur le dos de la reine dite désormais démandibulée puis celle-ci est replacée dans sa boite. Le miel permet une meilleure acceptation par ses congénères puisque cette première à été manipulée et peut perdre son identification. Papier filtre La reine Candy Ouvrières finissant l’élevage de la reine
Index
Mandibule
Figure 8: Démandibulation d'une reine (Source : Alban MAISONNASSE) a) Contention et arrachage de la mandibule d’une reine b) Trous après arrachage des mandibules
c) Glandes mandibulaires extraites à l’aide de pinces d) Résultat de la démandibulation
3) Mise en cagette
Deux jours après la démandibulation (19ème jours), trois groupes sont constitués, leur nombre variant suivant le nombre de reines nées, et le nombre de reines démandibulées ayant survécu : « reines démandibulées » ; « reines normales » (témoins +) ; « absence de reine » (témoin -).
Un ou deux jours avant chaque mise en cagette, une visite des ruchers est réalisée afin de récupérer des abeilles naissantes. Les abeilles naissantes correspondent à des abeilles de moins de trois jours. Elles sont récupérées dans différentes ruches et à différents ruchers la veille de la mise en cagette. Le but est de repérer les cadres et de faire une estimation de nombre d’abeilles prête à émerger. Pour cela, il faut se référer au nombre de cellules operculées, en gratter l’opercule de quelques unes pour regarder si un nombre suffisant ouvrières sont prêtes à en émerger.
a
b
a
c
a
d
c
Glande mandibulaire extraite Mandibule extraiteCandy Barrette de cire
Abreuvoirs
Les reines sont placées dans des cagettes d’élevage, ou cagettes pain (mises en place par Pain en 1966) contenant 100 abeilles naissantes, du pollen broyé dans un mortier et mélangé avec du miel en guise de protéines, du candy pour l’énergie (sucre), et des barrettes de cire afin de faciliter la construction (Fig.9). Ces cagettes sont ensuite placées à l’étuve pendant 16 jours à 34°C afin d’observer la construction. Elles sont entretenues tout les 2-3 jours : ajout de candy (nourriture), changement de l’eau et du papier buvard, comptage des ouvrières mortes.
Figure 9: Cagette pain pendant la construction du rayon (Source : Geoffrey MONTES)
Seize jours après l’émergence, deux cas de figure se présentent après 16 jours de mise en cagettes pain : construction ou non. Les reines sont alors retirées des cagettes pain afin de les séparées des ouvrières. Chaque cagette est ensuite placée au congélateur afin de tuer les toutes les ouvrières de chacun des traitements. Toutes ces ouvrières sont alors classées en les identifiant suivant le traitement, le numéro de cagette et la répétition. Puis elles sont conservées dans des sacs et au congélateur pour les manipulations futures (dissection des ovaires des ouvrières). Pour leur part, les reines ayant été récupérées sont placées dans des cagettes pain en présence d’une cinquantaine d’abeilles naissantes. Les reines seules ne peuvent survivre. Les nouvelles ouvrières lui permettront d’assurer les échanges nécessaires (trophallaxie par exemple) en attendant qu’elles soient utilisées pour l’observation du comportement de cour. Ouvrière morte Pollen Papier filtre Construction
Suite à cela débute l’analyse des constructions. Chaque construction est pesée, le nombre et la taille des cellules les constituants sont mesurés. La présence d’œufs ou la production de miel sont par ailleurs référencés.
II/ Comportement de cour
Pour étudier le phénomène de cour, deux expériences successives ont étés conduites.
1) Mise en cagettes
La première consiste à l’aménagement de cagettes. Elles ont étés constituées de pollen malaxé avec du miel en guise de protéines, de candy pour l’énergie, d’abreuvoirs, et de la cire étiré vide (morceau de rayon vide) placé au fond de celles-ci. Puis, par cagette, ont étés disposés une reine (reine témoin normale ou reine démandibulée), ainsi que 60 abeilles naissances récupérées au rucher. Deux jours après, l’étude du phénomène de cour est réalisée à la lumière solaire, ainsi que sous lampe rouge (la lampe rouge permet aux abeilles de se sentir comme dans la ruche).
2) Mise en ruches vitrées
Dans la deuxième expérience, chacune des reines démandibulées de chaque modalité et de chaque réplica est placée dans une ruche vitrée placée à l’intérieur du « rucher couvert » (pièce régulée du point de vue de la température). La ruche communique avec l’extérieur par l’intermédiaire d’un tuyau (Fig. 10). Elle est constituée de deux vitres qui permettent une meilleure observation des abeilles : distinction des castes, des « métiers » de la ruche, du phénomène de trophallaxie, etc. Dans notre cas, nous nous intéresserons au comportement de cour.
Figure 10: La ruche vitrée (Source : Geoffrey MONTES)
Dans un premier temps, un unique cadre est inséré, puis, 350g d’abeilles naissantes sont introduites. Ce qui équivaut à environ 3500 abeilles (une abeille pesant environ 0,1g). La reine, marquée au vernis sur le thorax pour une meilleure visibilité, est ensuite introduite. Dans un premier temps, celle-ci est déposée dans une petite boite percée de trous à l’intérieur de la ruche. Ceci permet à la reine de s’acclimater, aux phéromones de circuler, et ainsi aux autres abeilles de s’imprégner de son odeur. Dans cette boite est déposé du candy qui empêche la reine de sortir temporairement. Au bout d’un certain temps, le candy aura été consommé et la reine pourra sortir de la boite et rejoindre le reste de la ruche. Une fois la reine libérée, le phénomène de cour peut être observé facilement. Concernant les observations, la reine est caractérisée par une démarche plus lente, et se déplace entourée d’une cour qui l’escorte, la lèche et prend grand soin d’elle. Le nombre d’ouvrières participant au phénomène de cour est relevé, ceci par l’intermédiaire de photos qui seront visualisées plus tard.
Deux ruches vitrées ont étés mises en place. La manipulation est répétée autant de fois qu’il y a de reines (démandibulées et non) par modalité et par réplica. A la fin de la manipulation, chaque reine est conservée au congélateur pour de futures manipulations. En effet, pour être certain de l’absence de phéromones mandibulaires chez les reines démandibulées, chacune d’elles va être analysée afin de vérifier si du 9-ODA est présent ou
Tuyau permettant la communication avec l’extérieur
Cadre constitué de réserves (miel)
Vitre où à lieu l’étude de différents comportements de l’abeille (phénomène de cour)
III/ Le développement ovarien
1) Dissection et évaluation des ovaires
Les ouvrières conservées au congélateur ayant participées à la construction des rayons sont récupérées. Pour chacune des cagettes (de chaque traitement et de chaque répétition), 20 abeilles sont choisies au hasard pour la dissection. La manipulation, étant excessivement longue, est réalisée par deux techniciens différents. Chacun d’entre eux prenant en charge une ou deux répétitions. Par ailleurs, cette manipulation est réalisée en aveugle afin de ne pas fausser les résultats.
La dissection des ovaires est réalisée sous loupe binoculaire. Sur chaque abeille, la tête, les ailes et les pattes sont retirés. L’abeille est déposée sur le ventre et deux incisions latérales délicates sont réalisées. Intestin et jabot sont retirés afin de laisser apparaître les ovaires. L’objectif ensuite est d’assigner un chiffre à chaque ovaire. Ce chiffre correspond à un stade du développement de l’ovaire sur une échelle d’ordre 5 (Velthuis, 1970 ; Pernal & Currie, 2000) (Annexe 3):
• Stade 1 : correspond à un ovaire très peu développé, d’aspect filiforme et translucide
• Stade 2 : il s’agit d’un ovaire peu développé, légèrement plus épais mais toujours translucide
• Stade 3 : représente un ovaire développé, d’aspect blanchâtre, avec la présence de quelques ovarioles6 peu distinguables
• Stade 4 : c’est un ovaire très développé plus épais que l’ovaire de type 3
• Stade 5 : correspond à un ovaire extrêmement développé, très épais, avec beaucoup d’ovarioles bien distinctes
2) Analyses statistiques
Les transformations usuelles ne permettant pas d’obtenir des variables qui satisfassent aux conditions d’utilisation des ANOVAs, des méthodes non paramétriques ont été utilisées. Il s’agit ici d’une analyse inter-sujet à une variable indépendante (le stade ovarien) et à trois modalités (« Sans reine », « Reine démandibulée » et « Reine normale »). Pour chaque essai, des tests de Kruskal-Wallis ont été utilisés pour étudier si il y a des variations dans chaque même groupe pour chaque essai. Puis des tests de Mann-Whitney sont utilisés afin de voir si une différence significative se manifeste entre les trois groupes « Sans reine », « Reine démandibulée », « Reine normale » pour chaque essai.
IV/ Extraction du 9-ODA
Pour s’assurer de la disparition de toute trace des QMP chez les reines démandibulées, un protocole d’extraction du 9-ODA a été mis en place. L’expérience est effectuée sur toutes les reines démandibulées ou non. Pour les témoins (T+), une extraction préalable des glandes mandibulaires est réalisée. Puis, chaque reine est sectionnée en trois parties : tête/thorax/abdomen. Chacune de ces parties est ensuite broyées avec 200µl de méthanol et 11µl du standard interne : l’acide décanoïque à 250ng/µl dans un tube à 4ml. Le tout est dans un bac contenant de l’eau à 4°C pendant 2 minutes.
L’échantillon est ensuite centrifugé à 4000 tours/min à 4°C pendant 20 minutes. Afin de récupérer uniquement le surnageant dans un tube de 2ml, l’échantillon est évaporé sous flux d’azote auquel 25µ L de BSTFA (catalyseur) sont rajoutés. La solution est passée au vortex puis laissée à température ambiante pendant 40 minutes. Puis, le tout est dilué dans 100µl d’hexane. Enfin, on concentre un peu la solution pour l’injecter dans la colonne. Il s’agit d’une CPG à phase stationnaire apolaire et une phase mobile correspondant à du dihydrogène.
T- (10) T+ (19) Traitée (17) 0 1 2 3 4 5 6 7 Traitements D ia m è tr e m o y e n d e s c e ll u le s d e t o u te s l e s c o n s tr u c ti o n s (nb cagette total ayant construit)
Résultats
1)
La construction des rayons
Le diamètre moyen des cellules construites par des ouvrières ayant été en contact avec une reine démandibulée (ẍ = 5,21 mm) ne diffère pas du diamètre moyen des cellules construites par les ouvrières ayant été en contact avec une reine non opérée (ẍ = 5,13) (Fig. 11). Cependant, ces deux moyennes diffèrent du diamètre moyen des cellules réalisées par les ouvrières orphelines c’est-à-dire du traitement sans reine (ẍ = 6,15). Les mesures ont été prises dès que les colonies avaient construites des cellules bien discernables au bout de 16 jours de délai de construction.
Le nombre de cellules par construction dans le traitement « reine démandibulée » (ẍ = 93,47 cellules) n’est en moyenne pas différent du nombre de cellules construites par les ouvrières en présence d’une reine normale (ẍ = 63,37 cellules) (Fig. 12). En revanche, ces deux moyennes diffèrent l’une de l’autre par le nombre moyen de cellules par construction concernant le traitement « sans reine ». Les constructions des ouvrières n’ayant jamais été au contact avec une reine présentent très peu de cellules, alors que les constructions appartement aux ouvrières ayant été en présence d’une reine, démandibulée ou non, comportent de nombreuses cellules.
Figure 11: Variation du diamètre moyen des cellules entre les différents traitements, toutes séries confondues (Source : Geoffrey MONTES). T- = sans reine. T+ = en présence d’une reine vierge. Traitée = en présence d’une reine démandibulée.
T- (10) T+ (19) Traitée (17) 0 20 40 60 80 100 120 140 Traitements N o m b re m o y e n d e c e ll u le s p a r c o n s tr u c ti o n (nb cagette total ayant construit)
Des informations supplémentaires ont été relevées après 16 jours de construction : le poids de chaque construction et la présence de miel où d’œufs dans les cellules. Pour chacun des essais, les moyennes des poids des constructions réalisées par les ouvrières en présence d’une reine (démandibulée ou non) sont supérieures à la moyenne des poids des constructions établies par les ouvrières orphelines (sans présence royale). Concernant les œufs ou la production de miel, ceci est variable suivant le numéro de cagette et l’essai (Annexe 4, 5 et 6).
2)
Le phénomène de cour
La première expérience n’a donné aucun résultat. Les morceaux de cire étirée vides placés au fond des cagettes étaient mal agencés et ne comblaient pas entièrement le fond de celles-ci. Aucun phénomène de cour n’a pu être observé.
La deuxième expérience, conduite chez les reines démandibulées uniquement, toutes séries confondues, fut une réussite. La facilité d’observation des comportements des abeilles dans une ruches vitrée a permis de montrer que la reine manifestait toujours la présence d’une escorte autour d’elle, même en absence de glandes mandibulaires. La cour royale se constitue en moyenne, toutes séries confondues, de 10,34 ouvrières (tableau 3). Par ailleurs, des cellules au format mâle ont été construites à la périphérie du cadre des deux ruches vitrées.
Tableau 3: Moyennes du nombre d'ouvrières constituant la cour royale (Source : Geoffrey MONTES)
Numéro de série Moyenne du nombre d'ouvrières constituant la cour 1 10,19 2 9,02 3 11,82
Figure 12: Nombre de cellules moyen par construction, suivant différents traitements, toutes séries confondues (Source : Geoffrey MONTES). T- = sans reine. T+ = en présence d’une reine vierge. Traitée = en présence d’une reine démandibulée.
1 1,2 1,4 1,6 1,8 2 2,2 2,4 2,6 2,8 3
Pas de reine Reine Normale Reine demandibulée
s c o re m o y e n ( + S E ) ** * NS
3)
Le développement ovarien des ouvrières
La dissection des ovaires des ouvrières montrent pour chacun des essais les mêmes tendances. Cependant, les résultats sont légèrement différents puisque deux manipulateurs différents se sont succédé pour faire cette expérience. L’essai I, réalisé par un premier manipulateur, ne montre au travers des résultats obtenu aucune différence significative dans le stade de développement entre le groupe «reine normale» et le groupe «reine démandibulée» (Fig. 13), le score étant en moyenne de 1,94 pour le traitement T+ et de 1,9 concernant les reines sans QMP. On constate que ces deux groupes ont en revanche une différence significative dans leur développement ovarien avec le groupe « sans reine » (p < 0,001 ; test non paramétrique de Mann-Whitney), le score des T- s’élevant en moyenne à 3,26.
1 1,5 2 2,5 3 3,5 4
Reine Normale Pas de reine Reine demandibulée
s c o re m o y e n ( + S E ) *** NS
Figure 13: Score moyen du développement ovarien pour la série I
(NS : non significatif ; ***p < 0,001 ; test de Mann-Whitney) (Source : Geoffrey MONTES)
Sans reine Reine normale Reine démandibulée
Figure 14: Score moyen du développement ovarien pour la série II
1 1,2 1,4 1,6 1,8 2 2,2 2,4 2,6 2,8 3
Sans Reine Reine Normale Reine Démandibulée
s c o re m o y e n ( + S E ) *** NS
Les essais II et III ont été réalisés par un autre manipulateur. Le second ne révèle aucune différence significative entre les deux groupes où la reine est présente, qu’elle soit démandibulée ou non (Fig. 14). Cependant, une différence a pu être observée entre le groupe « reine normale » et « sans reine », avec un score de développement ovarien moyen respectivement de 1,94 et de 1,85 (0,01 < p < 0,05, Mann-Whitney), et entre le groupe « reine démandibulée » avec un score moyen de 2,17 et « sans reine » (0,001 < p < 0,01 ; Mann-Whitney). Le dernier essai ne montre, comme les deux premiers, aucune différence significative dans le stade de développement entre « reine normale et reine démandibulée en se qui concerne le stade d’évolution de la gonade femelle (Fig. 15). Seule une nette différence est observée entre ces deux groupes (score moyen : T+ = 1,93 ; Dém. = 1,93) et le groupe « sans reine » (score moyen T- = 1, 2,57) (p < 0,001 ; Mann-Whitney).
4)
Extraction du 9-ODA
Seules des expériences préliminaires ont pu être réalisées sur 3 reines normales ainsi que 3 reines démandibulées de 2006. Pour confirmer l’absence de 9-ODA dans le corps des reines démandibulées, la CPG et le SM ont permis d’obtenir les résultats suivants : les reines normales présentent du 9-ODA uniquement dans la tête, thorax et abdomen n’en ont révélé aucune trace ; les reines démandibulées ne présentent aucune trace de 9-ODA que ce soit au niveau de la tête, du thorax ou bien de l’abdomen.
Les extractions potentielles de 9-ODA dans les reines démandibulées ou non, ayant permises ces expérimentations, n’ont pas encore été réalisées.
Figure 15: Score moyen du développement ovarien pour la série III
Discussion
La perfection et le développement ordonné d’une communauté d’abeilles est un sujet d’étude fascinant. Au cœur de cette activité se trouve la reine, productrice de phéromones, régulant la vie dans la colonie.
Tout au long des manipulations, un grand nombre de reines étaient mortes. A partir des 100 larves greffées pour chaque répétition, seulement une vingtaine de reines normales émergeaient. En effet, le greffage des larves dans les cupules est une manipulation délicate. Il faut prendre soin de ne pas blesser les larves, qui, dans ce cas, ne se développeraient pas. Tandis qu’un tiers des reines émergées constituait la modalité « reines normales », les deux autres tiers de l’effectif subissaient une manipulation de démandibulation. Ici aussi, les pertes furent énormes puisqu’en moyenne la moitié des reines opérées n’ont pas survécu. La démandibulation est une opération extrêmement complexe pour nous expérimentateurs, et douloureuse pour les reines qui la subisse. Le matériel biologique est un matériel fragile, d’où la nécessité de travailler avec le plus grand soin possible.
1.
Le choix du format des cellules lors de la construction
Notre étude permet de constater que sans reine les abeilles peuvent adopter une stratégie de construction des cellules au format mâle (Fig. 11). Dans cet exemple, nous avons placé des ouvrières dans une situation analogue à une colonie orpheline, et celle-ci construisait tout d’abord des cellules au format mâle quand il y avait construction de rayon. La présence de la reine et du couvain dans une colonie normale inhibe les œufs des ouvrières se développant. Quand la reine est perdue, cette inhibition est levée, les ouvrières commencent à développer des œufs non fertiles ce qui aboutira à des faux-bourdons (Winston, 1987). La présence de cellules au format mâle dans les ruches vitrées au cours de l’identification de phénomène de cour, marque par ailleurs une absence royale temporaire. Cette production de mâle est le seul mode de reproduction en absence de reine. Le cas échéant, la colonie sera orpheline et mourra d’ici quelques mois sans la production de nouvelles ouvrières qui maintiennent la colonie fonctionnelle.
Les résultats de Ledoux et al. (2001) montrent que la présence royale permet le contrôle du format de construction et la phéromone mandibulaire royale est le signal majeur
pour les abeilles en essaimage. Les colonies nouvellement fondées commencent à construire des cellules au format ouvrière (Free, 1987 ; Lee et Winston, 1987) et pendant que ces ouvrières se développent, des cellules au format mâle sont construites en périphérie des nids. Naumann et al. (1993) ont travaillé sur le mouvement de la QMP dans les colonies populeuses et non populeuses en utilisant du 9-ODA marqué, et ils ont constaté que les ouvrières appartenant aux colonies populeuses ont reçu des quantités plus faibles de 9-ODA radioactif que les colonies non populeuses. Il semblerait que dans les colonies populeuses la quantité de QMP émise par la reine pour chaque abeille est moindre, particulièrement à la périphérie, ce qui peut provoquer la construction en format mâle.
Concernant l’analyse des constructions avec les reines démandibulées, les résultats sont très explicites. Sur 100% des mini colonies « reines démandibulées » ainsi que pour 100% des témoins « reines normales » qui ont construit, la construction présente de nombreuses cellules mais celles-ci sont plus petites (Fig. 11 et 12). La construction est donc rapide et en format ouvrière. Les phéromones royales des glandes mandibulaires ne sont, par conséquent, pas les seules à jouer un rôle dans le choix du format de construction. En outre, puisque sans reine, la construction est bien plus lente et s’effectue au format mâle (cellules plus grandes mais moins nombreuses) (Fig. 11 et 12), ceci montre bien que c’est la reine qui indique le choix de la construction, grâce aux phéromones dérivant d’une part de ses glandes mandibulaires mais aussi grâce à un autre paramètre qui correspond à des phéromones autres que les QMP.
La taille des cellules de type ouvrière et mâle, ici respectivement de 5,13 et 6,15 mm, était légèrement plus grande que les valeurs moyennes citées dans les études antérieures (Winston 1987 ; Pratt, 1998). Ceci pourrait être du à plusieurs facteurs, particulièrement à leurs bases génétiques ou à leur cloisonnement dans des mini-colonies.
Par delà ces affirmations, le poids des constructions varie considérablement d’une mini-colonie à une autre. Cependant, le poids des constructions, étant relativement lié aux nombre de cellules dans chacune d’elles, suit les mêmes tendances (Annexes 5, 6 et 7). De plus, certaines colonies commençaient à faire des réserves dans leurs cellules alors que d’autres commençaient à peine à construire ce qui fait varier le poids des constructions.
identiques, et chaque traitement devant impérativement être effectué dans des étuves différentes, nous avons du placer les reines démandibulées dans une étuve différente, d’où la possibilité d’une imprécision dans les résultats. Par ailleurs, du fait de la présence d’une quantité importante de cagettes pain dans chaque étuve, la ventilation avait peut être du mal à se réaliser, et des cagettes avec un taux d’humidité inhabituel étaient constatées.
2.
Le cortège royal
L’expérience menée avec les cagettes aménagées n’a permis d’obtenir aucun résultat. La reine pouvait seulement être aperçue suivie d’une escorte mais il était impossible de prendre de quelconques mesures. L’utilisation d’une lampe rouge, sensée apporter aux abeilles la sensation d’être comme dans une vraie ruche et de se déplacer librement dans celle-ci, n’a pas apporté davantage d’informations, c’est pourquoi un deuxième protocole a été mis en place en utilisant des ruches vitrées.
Nos expériences avec les reines démandibulées dans les ruches vitrées nous à permis d’observer divers comportement à l’intérieur même de la ruche. En temps normal huit ouvrières ou plus sont présentes quand la reine est immobile, mais le contact est généralement vite perdu quand la reine se met à marcher rapidement le long du rayon (Free et al., 1987). Par exemple, en moyenne 6,2 ouvrières ont étés dénombrés quand la reine se déplace, augmentant à 8,7 quand elle pond un œuf et à 10,8 quand elle est stationnaire (Allen, 1956). La quantité d’ouvrières léchant et palpant les reines augmente quand celles-ci s’accouplent et pondent des œufs. Mais quand elles deviennent trop vieille et pondent seulement des œufs haploïdes elles reçoivent moins d’attention (Free et al., 1987). Dans notre étude, une moyenne de 10,3 ouvrières a été dénombrée autour de la reine démandibulée quand celle-ci était stationnaire (Tableau 3). Ceci révèle donc qu’un autre paramètre, hormis les QMP, peut être responsable de ce comportement.
Keeling et al. avaient identifié en 2003 quatre composés produits par la reine et qui fonctionneraient en synergie avec les QMP, formant les QRP, dans le but de constituer son escorte. Ici, une reine dépourvue de glandes mandibulaires donc de QMP (à vérifier avec l’analyse par CPG des extraits de reine) présente un phénomène de cour, donc la phéromone royale ne paraît pas être une molécule essentielle pour la manifestation d’une cour escortant la reine, et il est possible que d’autres paramètres participent à ce phénomène.
3.
L’inhibition ovarienne
Les résultats de notre étude démontrent clairement que les QMP empêchent le développement des ovaires chez les ouvrières. Nous n’avons trouvé aucune différence significative entre les reines démandibulées et les reines normales sur l’impact du développement ovarien (Fig. 13, 14 et 15). En effet, la présence de reines dépourvues de glandes mandibulaires inhibe quand même le développement des ovaires. Ceci suggère la présence d’autres molécules de reine qui pourraient empêcher le développement ovarien chez les abeilles européennes à miel.
Cependant, dans le cas d’une colonie orpheline ou dans le cas d’une surpopulation de la colonie, les ouvrières peuvent développer leurs ovaires (Groot & Voogd, 1954). Cette stérilité facultative peut permettre à certaines ouvrières de développer des follicules ovariens et ainsi de produire des œufs mâles haploïdes provenant d’œufs non fertilisés. Moins de 1% d’œufs mâles dans une colonie d’abeilles, où une reine féconde est présente, provient des ouvrières (Visscher, 1989). En revanche, si une colonie perd sa reine et devient orpheline, les ouvrières ont la capacité de développer rapidement leurs ovaires ainsi que leur comportement de ponte. Hess (1942) a montré que les ovaires de 10% des ouvrières s’étaient développés suite à l’enlèvement de la reine pendant une période d’une semaine ; après 13 semaines d’absence, 87% des ouvrières ont augmentées la taille de leurs ovaires.
Cependant, même si l’expérience a été réalisé en aveugle (les manipulateurs ne connaissaient pas l’appartenance des ouvrières pour tel ou tel traitement), elle reste relativement imprécise. En effet, le fait que deux techniciens de laboratoire aient manipulé sur un ou deux essais, la notation reste différente ce qui peut fausser légèrement les résultats. En revanche, la finalité devrait rester la même.
Par ailleurs, pour faciliter le transfert les ouvrières des cagettes où avait lieu la construction jusqu’au congélateur, une dose non réellement mesurée de CO2 leur a été donnée. Cette dose
administrée était légèrement différente d’une cagette à une autre, ce qui peut fausser légèrement les résultats, d’autant plus quand on sait qu’une dose conséquente de CO2 peut
Les expériences ont permises d’obtenir un grand nombre de reines démandibulées, donc exemptes de toutes traces de QMP. Cependant, même si des études préliminaires ont été réalisées concernant la démandibulation et ont prouvé que suite à une telle opération, la reine ne présentait plus de phéromones royales dans le corps entier, les données de la chromatographie en phase gazeuse et du spectromètre de masse, qui sont des techniques relativement longues et couteuses, restent essentielles pour valider ces résultats. Si les analyses ne révèlent pas la présence de QMP dans le corps de ces reines démandibulées, des comparaisons de profils chimiques entre reines démandibulées et ouvrières pourrait permettre la recherche des molécules impliquées dans ces différents phénomènes, en plus de la phéromone mandibulaire royale.