© Catherine Couture, 2020
Modélisation de l'utilisation du phosphore et du calcium
alimentaires chez le poulet de chair
Mémoire
Catherine Couture
Maîtrise en sciences animales - avec mémoire
Maître ès sciences (M. Sc.)
Modélisation de l’utilisation du phosphore et du calcium
alimentaires chez le poulet de chair
Mémoire
Catherine Couture
Sous la direction de :
iii
Résumé
Une gestion précise des apports de phosphore (P) aux volailles est un enjeu important dans le domaine de la production avicole où il demeure un nutriment essentiel pour les performances des oiseaux, mais se doit d’être utilisé de façon précise pour nourrir les plantes afin d’éviter les problèmes environnementaux qui y sont associés. Le but de ce travail de maîtrise était de simuler le devenir du P alimentaire chez le poulet de chair à l’aide de la modélisation mécaniste en 1) évaluant la digestibilité iléale du P chez le poulet de chair à l’aide de l’outil de méta-analyse, 2) simulant la croissance des tissus protéique et lipidique et 3) en estimant la répartition de ces minéraux dans les différents tissus pour moduler l’excrétion du P selon les objectifs de production (ex. : optimisation des performances, diminution de l’excrétion de P). Dans un premier temps, la méta-analyse a permis d’étudier les interactions entre le calcium (Ca), les différentes formes de P et la phytase microbienne (PhytM). De façon générale, les rations riches en P phytique (PP) et en Ca ont montré une réponse plus élevée en termes de P digestible à la PhytM. Dans un deuxième temps, le modèle a permis d’estimer la répartition du P et du Ca dans les différents tissus, et ce, selon différents apports phosphocalciques. Ces simulations ont accentué l’importance d’un ratio Ca:P équilibré pour une utilisation optimale des deux minéraux par les poulets de chair et ce ratio doit être mis en lien avec l’apport de P donné aux oiseaux. En effet, considérant que les deux se déposent ensemble au niveau de l’os sous la forme d’hydroxyapatite, si le premier est apporté en excès par rapport à l’autre, il sera excrété et ainsi perdu engendrant une perte économique pour le producteur. Ce travail de maîtrise a donc permis d’avoir une meilleure compréhension des mécanismes contrôlant l’utilisation digestive et métabolique de P en lien avec les apports de Ca et de phytase microbienne et de développer un modèle mécaniste permettant de simuler le devenir métabolique du P et du Ca chez le poulet de chair. Les connaissances générées contribuent au développement d’outil d’aide à la formulation répondant aux enjeux de la durabilité de la production avicole.
iv
Table des matières
Résumé ... iii
Table des matières ... iv
Liste des tableaux ... vii
Liste des figures ... viii
Liste des annexes ... ix
Liste des abréviations ... x
Remerciements ... xi
Avant-propos ... xii
Introduction ... 1
Chapitre 1 Synthèse bibliographique ... 2
1.1. Le phosphore dans l’alimentation des poulets de chair ... 3
1.1.1. Formes de phosphore ... 3
1.1.2. Phytase ... 3
1.1.3. Utilisation du phosphore par l’animal ... 5
1.1.3.1. Flux de phosphore ... 6
a. Intestin ... 6
b. Reins ... 8
c. Os ... 9
1.1.3.2. Régulation de l’homéostasie phosphocalcique ... 11
a. Parathormone ... 11
b. Calcitriol ... 13
c. Calcitonine ... 14
1.1.4. Apports alimentaires de phosphore chez le poulet de chair ... 14
v
1.1.4.2 Estimation des besoins en phosphore chez le poulet de chair ... 15
1.1.4.3 Modélisation ... 16
1.1.4.4 Méta-analyse ... 19
1.2. Phosphore et développement durable ... 22
1.2.1. Apports vs besoins ... 23
1.2.2. Calcium ... 23
1.2.3. Déplétion-Réplétion ... 24
1.2.4. Alimentation de précision ... 25
1.3. Liste ... 28
Chapitre 2 Méta-Analyse de la digestibilité iléale du phosphore chez les poulets de chair : Effet du phosphore alimentaire et du calcium et de la supplémentation en phytase ... 37
Résumé ... 39
Abstract ... 40
2.1. Introduction ... 41
2.2. Matériels et méthodes ... 41
2.2.1. Collecte de données et codage ... 41
2.2.2. Calculs et détermination des variables dépendantes et indépendantes ... 41
2.2.3. Analyses statistiques et du méta-dispositif ... 42
2.3. Résultats et discussion ... 44
2.3.1. Effets des niveaux de phosphore phytique et non-phytique et du calcium sur le phosphore digestible ... 44
2.3.2. Effet de la phytase microbienne et de ses interactions sur le phosphore digestible... 45
2.3.3. Digestibilité iléale des différentes formes de P ... 46
2.4. Conclusion ... 47
vi
Chapitre 3 Modélisation de l’utilisation du phosphore et du calcium alimentaire chez le poulet de chair
... 56
Résumé ... 58
Abstract ... 59
3.1. Introduction ... 60
3.2. Matériels et méthodes ... 60
3.2.1. Prédiction de la consommation journalière ... 60
3.2.2. Prédiction du devenir du phosphore et du calcium ingéré et du dépôt de cendres corporelles ... 61
3.2.2.1. Devenir digestif du phosphore et du calcium ... 62
3.2.2.2. Devenir métabolique du phosphore et du calcium ... 63
3.2.3. Prédiction des dépôts protéiques et lipidiques et calcul du gain de poids journalier .. 64
3.3. Résultats et discussion ... 65
3.3.1. Comportement du modèle ... 65
3.3.1.2. Effets de déséquilibredes apports phosphocalciques sur leur rétention ... 66
3.3.1.3. Consommation théorique ... 67
3.3.1.4. Dépôt protéique ... 67
3.4. Conclusion ... 68
3.5. Liste des ouvrages cités ... 69
Conclusions et perspectives ... 72
Bibliographie ... 74
vii
Liste des tableaux
Tableau 1.1. Niveau d’organisation (adapté de (France et Kebreab, 2008))... 17 Tableau 2.1. Statistiques descriptives de la base de donnéesa ... 43
Tableau 2.2. Prédiction du P digestible en fonction de PNP (g/kg), PP (g/kg), PhytM (FTU/kg) et Ca (g/kg) ... 44 Tableau 2.3. Prédiction du P digestible en fonction des formes d’apports de phosphore ... 46
viii
Liste des figures
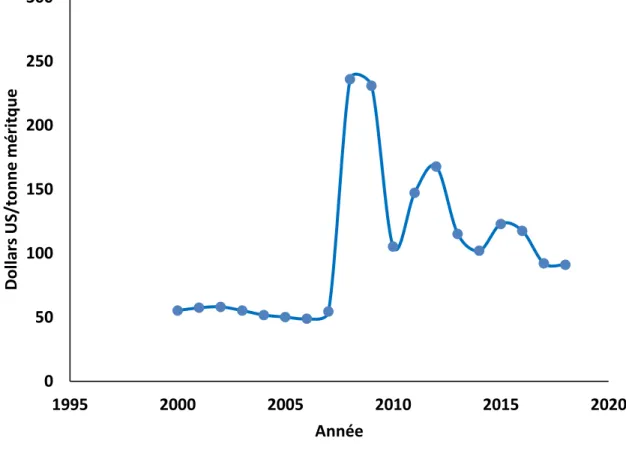
Figure 1.1. Prix annuels de la roche de phosphate entre 2000 et 2018 (World Bank, 2019) ... 4 Figure 1.2. Transport transcellulaire actif du P au niveau de l'intestin ... 7 Figure 1.3. Cycle du remodelage osseux. CB : cellule bordante, CMN : cellule mononucléaire, OB : ostéoblaste, OC : ostéoclaste, OCy : ostéocyte (adaptée de Bain et Watkins (1993)) ... 10 Figure 1.4. Voies de régulation du Ca et du P plasmatiques (adaptée de Li et al. (2017) et Proszkowiec-Weglarz et Angel (2013)). FGF23 : facteur de croissance des fibroblastes. ... 12 Figure 1.5. Métabolisme de la vitamine D (adaptée de Müller et Volmer (2015)) ... 13 Figure 1.6. Variation de la quantité de nutriment dans le temps entre deux compartiments (Éq. 1.3) 19 Figure 1.7. Les principales étapes d'une méta-analyse (adaptée de Sauvant et al. (2005)) ... 20 Figure 1.8. Concentration en nutriment distribué à des animaux selon un système d’alimentation quotidien, 3 phases ou 10 phases (adaptée de Pomar et al. (2009)) ... 25 Figure 1.9. Exemple du niveau d'incorporation de l'aliment initial (A) et de l'aliment final (B) dans des systèmes d'alimentation par mélange (adaptée de Pomar et al. (2009)) ... 26 Figure 2.1. Phosphore digestible (g/kg) en fonction du P non-phytique (PNP, g/kg), de la phytase microbienne (PhytM, FTU/kg) et du calcium pour un apport de P phytique de 2 g/kg... 45 Figure 3.1. Schéma général du modèle du devenir de calcium et de phosphore et de la croissance des différents tissus. ... 61 Figure 3.2. Dépôt corporel de calcium (a) et de phosphore (b) en fonction des apports phosphocalciques. ... 66 Figure 3.3. Bilan de calcium (a) et de phosphore (b) en fonction des apports phosphocalciques. ... 67
ix
Liste des annexes
Annexe 1. Affiche présentée lors de la 13èmes Journées de la Recherche Avicole et Palmipèdes à Foie Gras 2019 ... 91 Annexe 2. Premier article publié pour la 13èmes Journées de la Recherche Avicole et Palmipèdes à Foie Gras 2019 ... 92 Annexe 3. Deuxième article publié pour la 13èmes Journées de la Recherche Avicole et Palmipèdes à Foie Gras 2019 ... 98 Annexe 4. Paramètres du modèle ... 103 Annexe 5. Équations du modèle ... 107
x
Liste des abréviations
ATP Adénosine triphosphateCa Calcium CT Calcitonine CB Cellule bordante CMN Cellule mononucléée Fe Fer Mg Magnésium Mn Manganèse OB Ostéoblaste OC Ostéoclaste PTH Parathormone P Phosphore Pi Phosphore inorganique PNP Phosphore non-phytique PP Phosphore phytique PhytM Phytase microbienne Na Sodium
xi
Remerciements
Avant tout, j’aimerais remercier certaines personnes sans qui je n’aurais pu réaliser ce travail.
Je tiens tout d’abord à remercier Marie-Pierre Létourneau-Montminy, ma directrice et la personne avec laquelle j’ai débuté mon parcours en recherche lors de baccalauréat. Je la remercie pour toutes les opportunités qu’elle m’a permises de saisir dont mon stage en France et ma présentation au Poultry
Science Association Meeting au Texas. Je la remercie pour son soutien, sa disponibilité, ses conseils
et ses réponses à mes 1001 questions.
Je tiens également à remercier Agnès Narcy, Bertrand Méda et les autres collègues de l’Institut National de Recherche Agronomique de Nouzilly avec qui j’ai eu la chance de travailler pour m’avoir accueillie, moi et mon accent québécois, dans leur équipe. Mon séjour là-bas n’aurait pas été aussi agréable sans les autres étudiants/stagiaires présents lors de mon séjour : Anne Vallière, Apolline Gervais, Audrey Gloux, Denise de Senna Cardoso et Marine Dewez. Je les remercie de m’avoir intégrée aussi rapidement dans leur cercle d’amis et de m’avoir fait passer de bons moments en leur compagnie à l’extérieur du travail.
Je voudrais également remercier mes collègues de maîtrise/doctorat et amis du 4206 avec qui j’ai partagé mes heures de repas et quelques pauses afin de décrocher un petit peu : Élisabeth, Marie-Pier, Mélina et Phanie.
Finalement, je remercie mes parents de m’avoir soutenue dans mes études et de m’avoir encouragée à continuer malgré quelques difficultés au tout début de mes études post-secondaires, sans eux je ne me serais peut-être pas rendue aussi loin! Merci d’avoir cru en moi. Je tiens également à remercier Steven pour son soutien, sa patience et sa capacité à me convaincre de prendre de petites pauses. Je remercie également ma marraine, Clémence, qui m’a soutenue depuis mes débuts dans le monde de la recherche en nutrition avicole. Je souhaite également remercier Ghislaine Roch de m’avoir fait découvrir la merveilleuse profession qu’est d’être agronome et pour tous les conseils qu’elle m’a prodigués lors de mon parcours universitaire.
xii
Avant-propos
Ce mémoire comporte une introduction générale, suivie de trois chapitres et d’une conclusion générale. L'introduction générale est organisée comme suit, mise en contexte et description de la problématique et des objectifs de l’étude. Le premier chapitre est une synthèse bibliographique présentant le métabolisme du phosphore (P) et du calcium (Ca), les méthodes d’estimations des besoins en P des poulets de chair ainsi que l’aspect durable du phosphore dans la production avicole. Le second chapitre, rédigé sous forme d'article scientifique, a permis d’étudier les interactions entre les formes de P, le Ca et la phytase microbienne sur la réponse du poulet en termes de digestibilité iléale de P. L’article du chapitre 2 n’a subi que de légères modifications (présentation) sans changement majeur du contenu de l’article par rapport à la version publiée le 20 mars 2019 et le premier auteur est moi-même et les co-auteurs sont Agnès Narcy, Rachel Chiasson et Marie-Pierre Létourneau-Montminy. Le troisième chapitre, rédigé également sous forme d'article scientifique, a permis d’évaluer l’utilisation du P et du Ca alimentaires chez le poulet de chair et d’étudier la répartition de ces deux minéraux dans les différents tissus du poulet de chair. L’article du chapitre 3 a subi quelques modifications (présentation et ajout de matériel) par rapport à la version publiée le 20 mars 2019 et le premier auteur est moi-même et les co-auteurs sont Bertrand Méda, Agnès Narcy et Marie-Pierre Létourneau-Montminy. Les deux articles publiés ont été rédigés par moi-même et j’ai reçu l’appui de Marie-Pierre Létourneau-Montminy, Bertrand Méda et Agnès Narcy via les suggestions de correction. Finalement, une conclusion générale présente les conclusions et perspectives de ce travail de maîtrise.
1
Introduction
Depuis un peu plus d’une décennie au Canada, la consommation de poulet par habitant ne cesse d’augmenter et surpasse même celle du bœuf. Afin de subvenir à cette demande croissante, les producteurs de poulets de chair se doivent d’augmenter leur production et, par conséquent, les déjections animales. Dans un contexte de durabilité des productions animales, il est primordial de trouver des voies alimentaires permettant l’optimisation de l’utilisation des nutriments nuisibles pour l’environnement si apportés en excès des besoins des plantes tel le phosphore (P). Il s’agit d’un nutriment essentiel au métabolisme des poulets de chair, mais qui est faiblement utilisé puisque le P des céréales présent dans les aliments destinés à la production avicole est sous la forme phytique (PP) (Sauvant et al., 2004). Le P non utilisé par les animaux peut se retrouver dans l’environnement et risque de causer des problèmes d’eutrophisation si apporté en excès des besoins des milieux récepteurs. De plus, une diminution des apports de P pourrait permettre de réduire les coûts d’alimentation dépendamment du prix des phosphates qui sont volatils. En effet, considérant la faible utilisation de PP chez les monogastriques, les aliments doivent être supplémentés en P non-phytique (PNP) via une source minérale, les phosphates ou les farines animales dans le contexte nord-américain. Les phosphates sont une ressource non-renouvelable et il est estimé que les réserves mondiales pourraient être épuisées d’ici 50 à 100 ans ce qui pourrait conduire à des augmentations de coûts si l’offre était réduite (Cordell et al., 2009). Les phosphates présentent également un problème environnemental puisqu’ils ne diminuent pas le PP présent dans la ration contrairement à l’enzyme phytase. En effet, l’ajout de phytase microbienne dans l’aliment est une des voies d’amélioration de l’utilisation du PP chez les monogastriques puisqu’en hydrolysant le PP, elle libère du P et le rend disponible pour l’animal et est ainsi non-excrété. Pour améliorer l’utilisation du P par les poulets de chair, le calcium (Ca) ne doit également pas être négligé puisqu’il interagit avec le P tant de façon négative que positive; le Ca est nécessaire pour le dépôt osseux de P mais, lors d’excès, le Ca peut nuire à son absorption.
Afin de répondre à ces différents enjeux, la présente étude avait deux objectifs soit 1) la prédiction de la digestibilité iléale du P chez le poulet de chair à l’aide de l’outil de méta-analyse ainsi que 2) la construction d’un modèle du devenir digestif et métabolique du P et du Ca alimentaire ainsi que de la dynamique de dépôts des cendres corporelles chez le poulet en croissance tel que précédemment réalisé chez le porc (Létourneau-Montminy et al., 2015).
2
3
1.1.
Le phosphore dans l’alimentation des poulets de chair
1.1.1. Formes de phosphore
Les sources de phosphore (P) utilisées dans les aliments pour monogastriques sont principalement d’origine végétale, minérale et animale (ex. : farines de viande, d’os, de poisson). Le P dans les sources végétales se présente sous deux formes : phosphore phytique (PP, forme organique) et phosphore non-phytique (PNP, forme inorganique). Le P est majoritairement sous forme de PP dans les produits végétaux (50 à 85% du P total) et, par conséquent, dans les aliments destinés aux monogastriques qui sont composés en grande partie de produits végétaux (65 à 75% du P total consommé par l’animal provient des végétaux). L’acide phytique (phytate) ou acide myo-inositol hexaphosphorique (IP6) est composé d’un radical inositol estérifié par six radicaux phosphates et
contient ainsi 28,2% de P (Pointillart, 1994). Le phytate est la réserve principale de P dans les grains (Kemme et al., 1999). La déphosphorylation de l’IP6 engendre différents produits (inositol-X-phosphate,
IPX) soit l’IP5, l’IP4, l’IP3, l’IP2 et l’IP1 et ce sont les trois derniers qui sont les plus susceptibles de passer la barrière intestinale (Pointillart, 1994). Quant au P provenant des sources minérale et animale, il est sous forme non-phytique. Seules les formes inorganiques solubles sont absorbables dans l’intestin, ainsi les molécules de PNP et de PP ne seront pas absorbées en quantités similaires. Les groupements phosphates liés au PP ne peuvent être absorbés qu’à la suite de leur hydrolyse par une enzyme, la phytase. En raison de la faible hydrolyse de PP par les monogastriques par manque de phytase endogène (Humer et al., 2015), il est nécessaire de supplémenter les aliments en PNP hautement disponible pour combler leurs besoins. Les phosphates de source minérale principalement sous forme monocalciques, monobicalciques et bicalciques sont utilisés pour satisfaire les besoins. Chez le poulet de chair, la biodisponibilité relative de ces différents phosphates par rapport au phosphate monosodique est la suivante: 91% pour le monocalcique, 85% pour le bicalcique hydraté, 80% pour le monobicalcique et 76 % pour bicalcique anhydre (Sauvant et al., 2004).
1.1.2. Phytase
L’ajout d’une source minérale de P permet une augmentation du P disponible pour l’absorption provenant de l’aliment. Une autre façon d’augmenter l’apport de P disponible est d’ajouter de la phytase dans l’aliment. En hydrolysant PP, la phytase libère du P. Il existe trois types de phytase : végétale, microbienne et endogène (présente dans l’intestin de l’animal). Certains végétaux (seigle > blé > orge)
4
ont une activité phytasique intrinsèque comparativement à d’autres qui en possèdent peu ou pas (maïs > soya) (Viveros et al., 2000). Cependant, les traitements (ex. : effet de la température, entres autres, lors de la granulation) appliqués lors de la fabrication de l’aliment peuvent impactés négativement l’activité de la phytase provenant des végétaux (Tran et Skiba, 2005). Les aliments destinés aux poulets de chair sont généralement granulés et subissent donc un traitement de chaleur. De ce fait, l’effet de la phytase végétale ne peut pas être pris en compte comme source de P dans la majorité des aliments destinés à la production avicole à la suite de l’inhibition de son activité par la chaleur. Il est de plus à noter qu’en Amérique du Nord le maïs est la céréale majoritaire. Suite à la forte hausse du prix des phosphates en 2008, lequel est redescendu par la suite (Figure 1.1), et les réglementations de plus en plus sévères sur l’épandage du P des fumiers/lisiers, le marché de la phytase microbienne a subi une grande expansion (Lei et al., 2013).
Figure 1.1. Prix annuels de la roche de phosphate entre 2000 et 2018 (World Bank, 2019)
La phytase microbienne ou exogène est celle qui est ajoutée à l’aliment. Une unité phytasique (FTU) est la quantité d’enzyme nécessaire pour libérer 1 µmol de phosphate inorganique par minute de 5,1 mol/l de phytate de sodium à un pH de 5,5 et à une température de 37°C (Engelen et al., 1994).
0 50 100 150 200 250 300 1995 2000 2005 2010 2015 2020 Dol la rs US /t on n e m é ri tq u e Année
5
Plusieurs phytases commerciales sont disponibles sur le marché et peuvent être ajoutées aux aliments sous formes solide avant la granulation ou liquide après la granulation de l’aliment pour éviter les problèmes de thermostabilité à des températures de granulation de plus de 80°C. Il est à noter que la plupart des nouvelles générations de phytase ne sont pas influencées par des températures élevées et peuvent donc être intégrées à l’aliment avant la granulation à la vapeur et toujours être efficaces après le traitement. Il existe deux types de phytase microbienne : 3-phytase et 6-phytase. Ces types de phytases microbiennes sont définies selon la position à laquelle la phytase débute l’hydrolyse de PP : position C3 ou C6 de l'anneau d'hexaphosphate de myo-inositol (Selle et Ravindran, 2007). Le poulet de chair, selon certaines conditions, peut hydrolyser une partie du PP alimentaire via la production de phytase endogène au niveau de la muqueuse intestinale. En effet, McCuaig et Motzok (1972) et McCuaig et al. (1972) ont montré qu’en présence d’apport faible en P, les poulets de chair ont la capacité d’augmenter la sécrétion de phytase endogène ainsi que l’activité des phosphatases alcalines au niveau de l’intestin. Le Ca aurait également un effet sur la phytase endogène puisque Tamim et al. (2004) ont montré que les poulets de chair n’ayant pas de Ca alimentaire présentaient une disparition iléale de PP plus élevée (62,9% vs 25,4%) que ceux recevant le régime contenant 5 g/kg de Ca alimentaire. McCuaig et al. (1972) et Applegate et al. (2003) ont également montré que le Ca avait un effet négatif sur l’activité de la phytase endogène. Le mode d’action exact n’est cependant pas bien compris.
Le pH optimum pour les phytases microbiennes se situe entre 3 et 5 (Brejnholt et al., 2011) et suggère que les segments supérieurs du tractus digestif (jabot, proventricule, gésier) seraient les sites actifs majeurs propices à une plus grande efficacité de la phytase microbienne (Selle and Ravindran, 2007; Li et al., 2016). La phytase endogène de l’animal serait quant à elle sécrétée dans la partie proximale de l’intestin et, plus précisément, au niveau du duodénum (Maenz et Classen, 1998).
1.1.3. Utilisation du phosphore par l’animal
Le P joue un rôle important dans la minéralisation osseuse. En effet, il forme avec le Ca l'hydroxyapatite (Ca10(PO4)6(OH)2) qui constitue 60 à 70% de la masse de l’os (Rath et al., 2000) et environ 80% du P
et plus de 95% du Ca corporel se retrouve dans le squelette sous la forme d’hydroxyapatite. Le 20% restant joue un rôle essentiel dans les fonctions métaboliques des organelles et des parois des cellules (phospholipides et phosphoprotéines) (Veum, 2010). Le P est également un composé des acides nucléiques (ADN et ARN) ainsi que des nucléotides comme l’ATP qui est un élément important du
6
métabolisme énergétique (Soares, 1995). Une carence en P se traduit par une perte d’appétit (et une faible croissance) qui se déclare bien avant que le squelette ne se déminéralise (Suttle, 2010). En cas de forte carence en P, le rachitisme (hypophosphatémique) peut se manifester et est caractérisé par une hypertrophie des chondrocytes et d’une accumulation de vaisseaux métaphysaires allongés dans la plaque de croissance (Lacey et Huffer, 1982). En présence de carence en Ca, le rachitisme (hypocalcémique) se manifeste différemment au niveau de l’os et se caractérise par l’épaississement de la plaque épiphysaire engendré par une prolifération des chondrocytes (Thorp et Waddington, 1997).
1.1.3.1. Flux de phosphore
Le P circule dans l’organisme de l’animal entre trois organes principaux : l’intestin grêle, les reins et les os.
a. Intestin
Chez les volailles, le P est majoritairement absorbé au niveau de l’intestin et, plus précisément, au niveau de la partie supérieure du jéjunum (Hurwitz et Bar, 1970). Le P peut être absorbé selon un processus de transport paracellulaire passif ou transcellulaire actif selon la concentration intraluminale en P (Barlet et Coxam, 1995). Le transport paracellulaire est généralement orienté de la lumière intestinale vers le plasma, mais ce type de transport est faible puisque la paroi épithéliale intestinale présente une faible perméabilité aux phosphates (Cross et al., 1990). Ces jonctions intercellulaires sont deux fois plus perméables aux cations monovalents qu’aux anions monovalents (Frizzell et Schultz, 1972). La diffusion passive du P inorganique (Pi) aurait probablement lieu au niveau du duodénum proximal où le pH acide (légèrement supérieur à 4) favorise la présence d’ions H2PO4-. Au
niveau du jéjunum et de l’iléon où le pH est d’environ 7,5, les ions HPO42- dominent et représentent
80% du Pi ionisé. Ainsi, l’absorption des phosphates alimentaires ne peut pas s’effectuer passivement puisque la concentration intraluminale des ions HPO42- devrait être d’environ 15 fois supérieure à celle
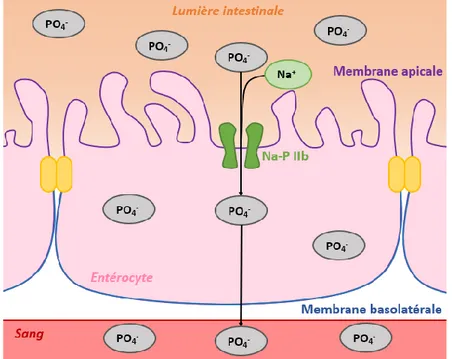
mesurée dans le cytoplasme des entérocytes. Ainsi, le transport de P est majoritairement transcellulaire (actif) chez le poulet de chair (Cross et al., 1990). Le transport transcellulaire actif du Pi (PO4-) est Na-dépendant (cotransport Na-Pi, Hildmann et al., 1982) et le transporteur Na-Pi le plus
présent au niveau de l’intestin est le Na-P IIb (Proszkowiec-Weglarz and Angel, 2013). Ce transport comporte 3 étapes (Figure 1.2) : 1) entrée dans les cellules épithéliales de l’intestin (entérocytes) via
7
la membrane de bordure en brosse (membrane apicale), 2) déplacement à travers la cellule et 3) sortie via la membrane basolatérale (Yan et al., 2007).
Le P présent dans le tractus digestif provient majoritairement de l’aliment, mais également des pertes endogènes. Le P attribuable aux pertes endogènes provient des sucs digestifs (biliaires et pancréatiques) ainsi que du renouvellement constant des entérocytes. Une excrétion plus élevée de P endogène est généralement observée dans le duodénum (Hurwitz et Bar, 1965) et ce sont les sécrétions biliaires et pancréatiques qui sont responsables de ces pertes endogènes de P. La bile qui est constituée principalement de sels biliaires, de phospholipides (lécithine ou phosphatidylcholine), de cholestérol et de protéines (Borgström et Patton, 1991) est sécrétée par deux conduits dans la partie médiane à distale de la boucle du duodénum. Les sécrétions pancréatiques qui sont principalement des enzymes digestives (phospholipase, amylase, lipase et protéases) et des électrolytes rejoignent le duodénum dans sa partie distale (Dibner et Richards, 2004). Les sucs digestifs (biliaires et pancréatiques) contiennent également des ions bicarbonates (HCO3-) qui ont pour
but de neutraliser l’acidité du bol alimentaire provenant de l’estomac. Une partie de ce P endogène est réabsorbée et l’autre partie est excrétée via les fientes. Les pertes endogènes de P chez le poulet de chair n’ont pas été largement étudiées et sont sujettes à certains facteurs de variations : stratégie d'alimentation en calcium (ratio Ca:P de 1,4 vs 3,5 g de Ca/kg d’aliment) et la durée de la période d'adaptation à l’aliment (0, 24 et 48h; Perryman et al., 2017) ainsi que par le type de céréale (80, 609,
8
418 et -87 mg/kg de matière sèche ingérée respectivement pour les aliments à base de blé, de tourteau de soya, de drèches de distillerie de maïs avec solubles et de sorgho). Mutucumarana et al., (2014) ont estimé que les pertes endogènes de P chez le poulet de chair pouvaient varier de 227 à 446 mg/kg de matière sèche ingérée tandis que Dilger et Adeola (2006) ont estimé qu’elles étaient de 235 mg/kg de matière sèche ingérée. Khaksarzareha et al. (2017) ont rassemblé les données de la littérature à ce sujet et ont déterminé les pertes endogènes fécales selon le poids vif de l’animal (où PV = poids vif : – 0,0021PV² + 0,0185 PV + 0,0034).
b. Reins
Chez l’homme, une réabsorption de 85% du P a lieu dans le tubule contourné proximal et moins de 10% est réabsorbé dans les segments tubulaires plus distaux (Courbebaisse et Souberbielle, 2011). La quantité de Pi excrétée est déterminée par les taux cumulés de trois processus: la filtration glomérulaire, la réabsorption tubulaire et la sécrétion tubulaire (excrétion urinaire). La quantité de P provenant de la filtration globulaire (plasma vers urine primitive) peut être calculée en multipliant le débit de filtration glomérulaire par la concentration de Pi ultrafiltrable dans le plasma. Le Pi dans le plasma est généralement considéré comme complètement ultrafiltrable malgré qu’une infime fraction puisse être liée aux protéines (Wideman, 1987). L’urine primitive provenant de la filtration glomérulaire subit par la suite la réabsorption de P par les tubules rénaux. Cette réabsorption tubulaire est possible grâce à des mécanismes de diffusion transcellulaire et paracellulaire comme dans l’intestin (Courbebaisse et Souberbielle, 2011). Comparativement aux mammifères, l’urine chez les volailles est excrétée avec les fèces via le cloaque rendant l’étude de l’excrétion urinaire minérale plus complexe. Cependant, dans une certaine mesure, la différence entre la digestibilité iléale et la rétention (collecte des fientes) peut représenter l’excrétion urinaire (Shastak et al., 2012). Peu d’études mesurant les pertes urinaires chez les poulets de chair ont été faites. Manangi et al. (2018) ont mesuré l’excrétion urinaire de P en effectuant des colostomies sur des poulets de chair âgés de 40 et 50 jours. Cette étude a montré qu’une augmentation de l’apport de PNP augmenterait la concentration plasmatique de Pi, ainsi qu’une augmentation de la rétention du P total et du PNP jusqu’à l’atteinte d’un état d’équilibre physiologique (7,51 mg Pi / dL - 8,13 mg Pi / dL). L'excès de P au-delà de ce seuil physiologique est éliminé dans l'urine via les reins et est associé à une diminution de la rétention du P.
9
c. Os
Le système osseux a tout d’abord un rôle mécanique de support et de locomotion. Il constitue également la réserve de P et de Ca de l’organisme puisque 99% du Ca et 80% du P corporels sont présents dans l’os sous forme d’hydroxyapatite (Proszkowiec-Weglarz et Angel, 2013). Le ratio Ca:P de l’os est relativement constant et se rapproche de celui de l’hydroxyapatite qui est de 2,2 (De Vries et al., 2010). De ce fait, l’os joue un rôle important dans la régulation de l’homéostasie phosphocalcique (Bain et Watkins, 1993). À l’éclosion, le squelette du poussin n’est que partiellement minéralisé puisque la minéralisation osseuse ne se complète qu’entre 4 à 11 jours d’âge (Williams et al., 2000).
L’os est composé d’une matrice minérale (inorganique, 70%) et organique (20%) et d’eau (10%). La matrice minérale est principalement composée de Ca et P présents sous la forme d'hydroxyapatite qui procure une rigidité et une résistance à la compression à l'os. Le collagène, une protéine, est le principal constituant de la matrice organique (80 à 90%) et confère à l’os une résistance à la traction ainsi qu’un support à la matrice minérale. Les protéoglycanes, les lipides et les protéines non collagènes telles que l'ostéocalcine, l'ostéonectine et les ostéopontines constituent l’autre partie de la matrice organique. Les protéines non collagènes contribuent à diverses fonctions de l’os, telles que la stabilisation de la matrice, la calcification ainsi que d'autres activités de régulation métabolique (Rath et al., 2000).
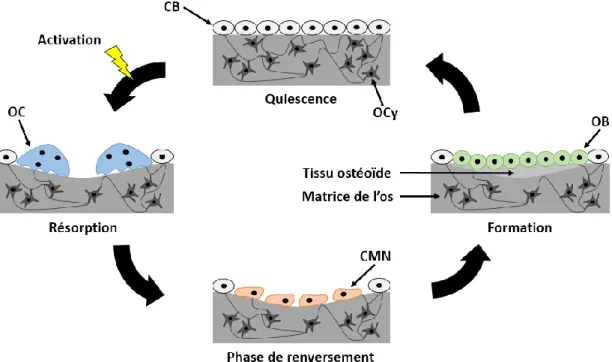
La formation et le renouvellement de l’os est assuré par plusieurs cellules osseuses (ostéoblaste, ostéoclaste, ostéocyte) ayant chacune un rôle spécifique. Les ostéoclastes sont caractérisés par leurs multiples noyaux et proviennent de précurseurs ayant pour origine des cellules souches hématopoïétiques et sont présents dans les espaces lacunaires (ou lacunes de résorption) qui sont créés lors de la résorption et comblés par le tissu ostéoïde lors de la formation de l’os (Figure 1.3). Ils sont responsables de la résorption osseuse. Ils déminéralisent et dégradent la matrice protéique osseuse en excrétant des protons et des protéases et libèrent ainsi des ions phosphates et calciques (Desoutter et al., 2012). Les cellules osseuses permettant la formation de l’os (fraction organique) sont les ostéoblastes. Ils proviennent de cellules souches d’origine mésenchymateuse et sont retrouvés dans la plaque de croissance, le périoste et l’endoste et sont alignés le long du tissu ostéoïde (matrice osseuse non-minéralisée). À la fin de la formation osseuse, les ostéoblastes deviennent des ostéocytes, des cellules bordantes ou meurent par apoptose. Les ostéocytes sont les cellules osseuses majoritairement (95%) retrouvés dans la matrice de l’os. Avec leur réseau de canalicules, ils sont en contact permanent avec les ostéoclastes et les ostéoblastes. Les ostéocytes jouent un rôle
10
important dans le remodelage osseux via l’activation des autres cellules osseuses en présence de contraintes mécaniques ou de dommages causés dans la matrice (Desoutter et al., 2012). En effet, au cours de sa croissance, l’os subit des changements de forme et de taille via le processus de remodelage osseux. Le remodelage osseux consiste en l’alternance d’une phase de résorption et d’une phase de formation osseuse nécessaire au maintien de l’intégrité du squelette et de l’homéostasie phosphocalcique. Le cycle de remodelage osseux (Figure 1.3) est initié lorsque les cellules bordantes (CB) présentes sur la surface osseuse quiescente se rétractent et exposent le minéral sous-jacent. Une fois le site de remodelage activé, les ostéoclastes (OC) débute la résorption osseuse. Au cours de la phase de renversement, les OC meurent par apoptose et laissent places aux cellules mononucléées (CMN) qui, elles, lissent la surface des lacunes de résorption créées par les OC et les ostéoblastes (OB) sont ensuite mobilisés pour remplir l'espace de remodelage lors une période de formation. Après la formation du tissu ostéoïde et le remplissage de la lacune de résorption, la surface osseuse revient à quiescence (Bain et Watkins, 1993).
Figure 1.3. Cycle du remodelage osseux. CB : cellule bordante, CMN : cellule mononucléaire, OB : ostéoblaste, OC : ostéoclaste, OCy : ostéocyte (adaptée de Bain et Watkins (1993))
11
1.1.3.2. Régulation de l’homéostasie phosphocalcique
Trois principales hormones sont impliquées dans la régulation de l’homéostasie phosphocalcique, soit la parathormone, le calcitriol et la calcitonine. Ces hormones agissent sur trois principaux organes : l’intestin, les reins et les os.
a. Parathormone
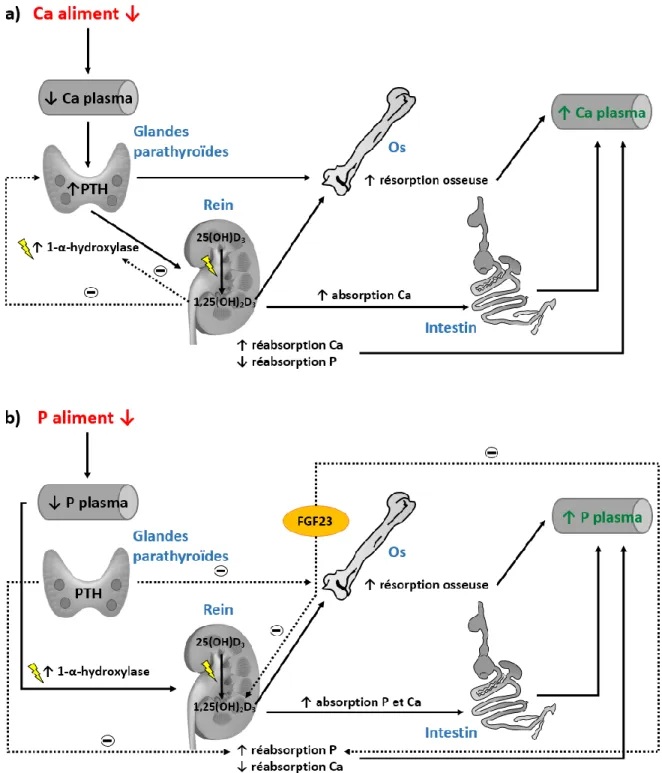
La parathormone (PTH) est synthétisée par les cellules principales des glandes parathyroïdes sous la forme d’un prépro-peptide comportant une préséquence de 25 acides aminés (séquence signal) et une proséquence de 6 acides aminés. La PTH mature (84 acides aminés) est obtenue après le clivage de ces deux séquences (Bergwitz et Jüppner, 2010). La PTH est sécrétée en présence de faible concentration de Ca dans le sang (hypocalcémie). La PTH répond aussi bien aux variations subites et à court terme qu’à une faible concentration soutenue de Ca plasmatique (Proszkowiec-Weglarz et Angel, 2013). Dans le cas d’hypocalcémie, la PTH stimule l’activité de la 1-α-hydroxylase qui permet la synthèse de la forme active de la vitamine D3 à partir de 25-hydroxyvitamine D3 (25(OH)D3) au niveau
des reins (Wideman, 1987) et engendre une réabsorption rénale du Ca. Une concentration élevée de la PTH diminue la réabsorption de P (augmente la sécrétion de P) (Proszkowiec-Weglarz et Angel, 2013) via l’inhibition de l’activité des cotransporteurs Na-Pi de type II au niveau de l’épithélium des tubules proximaux du rein (Dudas et al., 2002). La PTH engendre la mobilisation du Ca provenant de l’os pour maintenir une concentration constante du Ca plasmatique (Dittmer et Thompson, 2011). Les ostéoclastes (OC) permettent la mobilisation (résorption) du Ca en provenance de l’os, mais ces derniers subissent indirectement l’effet de la PTH via les ostéoblastes (OB). En effet, la PTH stimule l’expression de RANKL (récepteur activateur pour NF-кB ligand) et un ligand de surface sur les OB se lie aux OC via RANK (récepteur activateur pour NF-кB) ou à un récepteur leurre nommé ostéoprotégérine tous deux engendrant l’ostéoclastogenèse (Kousteni et Bilezikian, 2008). La Figure 1.4 montre, entre autres, l’effet de la PTH sur les différents organes impliqués dans l’homéostasie phosphocalcique en présence d’une diminution de l’apport en Ca ou en P.
12
Figure 1.4. Voies de régulation du Ca et du P plasmatiques (adaptée de Li et al. (2017) et Proszkowiec-Weglarz et Angel (2013)). FGF23 : facteur de croissance des fibroblastes.
13
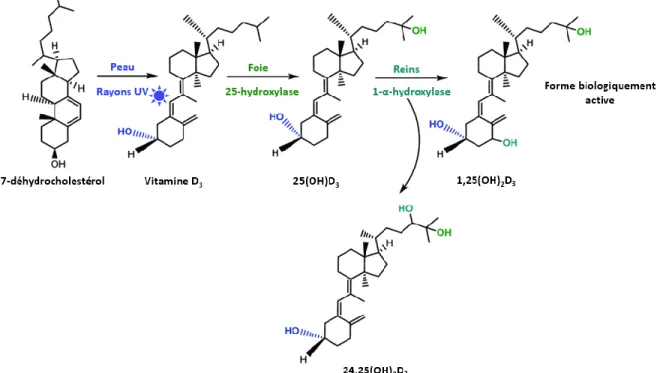
b. Calcitriol
La vitamine D3 (cholécalciférol) peut être synthétisée via l’isomérisation de 7-déhydrocholestérol en
vitamine D3 après l’exposition de la peau aux rayons ultraviolets ou via l’aliment ou la supplémentation
de vitamine D2 ou D3 (Figure 1.5).
Figure 1.5. Métabolisme de la vitamine D (adaptée de Müller et Volmer (2015))
Cependant, la vitamine D3 est davantage disponible puisque les protéines liant la vitamine D (vitamin
D binding protein) et ses métabolites ne se lient pas efficacement à la vitamine D2, forme
majoritairement retrouvée dans les végétaux. Les protéines se liant à la vitamine D3 (et ses métabolites)
permettent son transport au niveau du sang (Soares et al., 1995) vers le foie où elle sera hydroxylée par l’enzyme 25-hydroxylase en 25-hydroxyvitamine D3 (25(OH)D3), le principal métabolite (également
lié à une protéine) circulant de la vitamine D3. Le calcitriol (1,25(OH)2D3; 1,25-dihydroxycholécalciférol),
forme active de la vitamine D3, est par la suite synthétisé dans les reins via l’action de 1-α-hydroxylase
(Dusso et al., 2005). L’enzyme 1-α-hydroxylase est régulée par la PTH, la calcitonine ainsi que par un rétrocontrôle négatif de 1,25(OH)2D3 qui est principalement due à l’inhibition de la PTH lors du
rétablissement d’un niveau normal de Ca plasmatique. Le P plasmatique interagit également avec l’enzyme 1-α-hydroxylase sans toutefois dépendre des concentrations de PTH ou de Ca (Dittmer et Thompson, 2011).
14
Le facteur de croissance des fibroblastes, le FGF23, joue également un rôle dans le contrôle du métabolisme de la vitamine D et des phosphates. En effet, les deux rôles de FGF23, qui est produit au niveau de l’os par les ostéoblastes, sont de réduire la réabsorption rénale de P et de diminuer la synthèse de calcitriol au niveau du rein. Le FGF23 peut réduire la synthèse de vitamine D, mais cette dernière régule également ce facteur (Barthel et al., 2007). Un taux de FGF23 inadéquat en circulation dans le sang peut donc provoquer divers débalancements en P, en vitamine D et avoir des répercussions sur le métabolisme osseux (Yamazaki et al., 2008). Bobeck et al. (2012) ont montré que les poules ayant reçu une vaccination contre le FGF23 pouvaient transférer passivement les anticorps anti-FGF23 à leur progéniture. Les poulets présentant des anticorps anti-FGF23 induits par la poule et nourris avec un aliment déficient en P avaient une concentration en P plasmatique et des cendres osseuses comparables aux poulets présentant l’anticorps anti-FGF23 recevant un aliment adéquat en P. Une réduction du FGF23 plasmatique permettrait donc de réduire les besoins en P considérant que les rôles de FGF23 de limiter l’assimilation du P alimentaire et d’augmenter l’excrétion urinaire des phosphates (Yamazaki et al., 2008) se trouve ainsi réduits.
c. Calcitonine
La calcitonine (CT) est un polypeptide de 32 acides aminés sécrété par les cellules parafolliculaires C de la glande thyroïde en réponse à une hypercalcémie et une hyperphosphatémie (Jones et al., 1998). La CT inhibe la résorption osseuse en agissant directement sur les ostéoclastes diminuant ainsi la concentration de Ca dans le plasma (Chambers et Magnus, 1982). La CT agit également au niveau du métabolisme rénal via une plus grande excrétion de Ca. Cependant, la CT ne semble pas avoir d’impact majeur sur l’excrétion urinaire du Ca et du P chez la volaille (Wideman, 1987) ni chez le porc (Langlois et al., 2016). Chez l’humain, la CT joue un rôle dans la régulation du métabolisme de la vitamine D en maintenant la concentration plasmatique de 1,25(OH)2D3 en conditions
normocalcémiques. La CT induirait la transcription de 1-α-hydroxylase dans les cellules des reins (Zhong et al., 2009).
1.1.4. Apports alimentaires de phosphore chez le poulet de chair Le P présente un enjeu environnemental important puisqu’une mauvaise gestion de cet élément peut avoir des répercussions négatives sur les cours d’eau. Une connaissance des besoins des animaux, des facteurs affectant les apports, la digestibilité et l’utilisation des nutriments permettent une meilleure formulation des aliments. De plus, une formulation adéquate des aliments pour les poulets de chair
15
favoriserait l’atteinte des objectifs fixés en termes de performances, de coût d’alimentation et de contrôle des impacts environnementaux.
1.1.4.1 Méthodes d’estimation des besoins
Il existe deux méthodes d’estimation des besoins nutritionnels chez les animaux : la méthode empirique et la méthode factorielle. Dans la méthode empirique, les besoins nutritionnels sont définis comme la quantité minimale de nutriment nécessaire pour maximiser ou minimiser les réponses des animaux pour un ou plusieurs critères de performance (ex. : gain de poids, minéralisation osseuse), et ce, pendant une période donnée. Dans cette méthode, les besoins en un nutriment spécifique sont déterminés selon des essais dose-réponse comportant des aliments avec des doses graduellement croissantes en ce même nutriment. La méthode factorielle consiste à déterminer les besoins en combinant les besoins estimés d’entretien (pertes endogènes dans les fèces et l’urine), de croissance (déposition quotidienne de P nécessaire pour une minéralisation osseuse et une croissance des tissus mous optimales) et de production (applicable à la poule pondeuse et de reproduction) (Vieira et al., 2017). Cette dernière méthode nécessite donc la détermination d’un dépôt optimal obtenu via des compositions corporelles ou par bilan métabolique (ingéré-excrété).
1.1.4.2 Estimation des besoins en phosphore chez le poulet de chair
Les dernières recommandations en P pour les volailles sont celles du NRC (1994) et remontent donc à près de 25 ans. Une mise à jour des besoins en P des poulets de chair s’avère donc nécessaire bien que les compagnies donnent des recommandations pour leur génétique qui demeurent élevées et qui ont peu évoluées dans le temps. En plus de dater, il n’existe pas de consensus sur la façon d’exprimer la valeur P des ingrédients et les besoins des poulets. Le NRC (1984, 1994) exprime le besoin de P en P non-phytique qui est la différence entre le P total et le P phytique tous deux mesurables en laboratoire ce qui constitue un avantage. Le P disponible est utilisé au Canada et en Europe. La disponibilité de P d’une matière première est déterminée par des essais de dose-réponse permettant de comparer la source étudiée à une source de référence, souvent le phosphate monocalcique ou monosodique, à laquelle on attribue une valeur de 100 % de disponibilité. Le critère mesuré est généralement la minéralisation osseuse, et ce, chez des animaux en subcarence, qui reçoivent des doses croissantes de P provenant de la source étudiée ou de la source de référence. Après avoir vérifié la linéarité de la réponse du critère de minéralisation à l’apport de P pour chacune des deux sources, la disponibilité est calculée comme le rapport des deux pentes (Sauvant et al., 2004).
16
Afin d’uniformiser les données sur la valeur P des ingrédients, un comité de réflexion a été créé et ce dernier a recommandé d’utiliser la digestibilité pré-caecale (iléale) de P. Ce comité a également mis au point un protocole bien normé rendant la mesure facilement répétable (Rodehutscord, 2013). Cependant, malgré les efforts de standardisation de la méthode de mesure de digestibilité iléale de P, Rodehutscord et al. (2017) ont montré lors d’un essai impliquant 17 centres de recherche et les mêmes aliments que la digestibilité iléale de P du tourteau de soya variait de 19 à 51%. La relation entre la digestibilité iléale de P et la disparition d’IP6 était bien corrélée. La variation de digestibilité iléale de P
pourrait s’expliquer par le fait que les facteurs qui influencent l’hydrolyse d’IP6 (ex. : flore intestinale,
âge de l’abattage, niveaux d’apport de Ca et P) aient un impact important sur la variation de la digestibilité iléale de P. Les conditions pré-expérimentales d’alimentation et de logement (parquets vs cages, aliment démarrage), qui n’étaient pas contrôlées, pourraient avoir eu un impact sur ces facteurs. Par exemple, l’ingestion de la litière chez les oiseaux en parquets peut avoir affecté le développement général du tractus digestif et de la flore intestinale favorisant ou non la digestibilité de P (Rodehutscord et al., 2017).
1.1.4.3 Modélisation
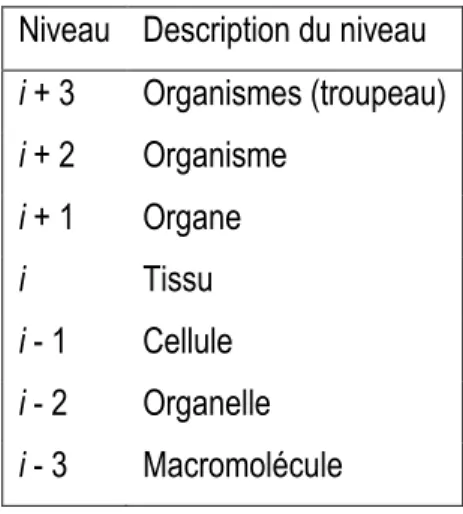
La définition des besoins nutritionnels des animaux a d’abord été possible via l’utilisation des systèmes d’unités d’alimentation. Les besoins nutritionnels étaient calculés à partir de la connaissance des besoins de production (protéiques, énergétiques, etc.) pour ainsi formuler un régime alimentaire permettant la couverture la plus précise possible de ces besoins. La mesure des flux d’entrée (ingestion) et de sortie (excrétions urinaires, fécales, etc.) a permis de concevoir les unités d’alimentation au sein de l’animal sans toutefois savoir ce qui se passe entre les deux. En d’autres termes, ce qui entre et sort de l’animal est connu, mais l’utilisation métabolique des nutriments au sein de l’animal est inconnue. Les objectifs principaux des productions animales étant de maximiser l’efficacité et la productivité, le système d’unités d’alimentation n’est donc pas suffisant et les réponses des animaux à différents régimes alimentaires doit être étudiée afin d’investiguer l’utilisation des apports nutritionnels (Sauvant, 1992). Une meilleure compréhension des systèmes sous-jacents de l’organisme (Tableau 1.1) est donc nécessaire. Tous les niveaux présentés au Tableau 1.1 peuvent être considérés comme étant un système composé de sous-systèmes situés à un niveau inférieur, ou en tant que sous-système de systèmes de niveau supérieur (France et Kebreab, 2008).
17
Tableau 1.1. Niveau d’organisation (adapté de (France et Kebreab, 2008))
Niveau Description du niveau
i + 3 Organismes (troupeau) i + 2 Organisme i + 1 Organe i Tissu i - 1 Cellule i - 2 Organelle i - 3 Macromolécule
L’étude de certains des systèmes sous-jacents peut s’avérer complexe et plus ou moins nécessaire dépendamment de l’objectif. La modélisation serait cependant envisageable pour, par exemple, vouloir étudier l’effet de la nutrition sur les performances de croissance. La modélisation est une partie centrale et intégrale de la démarche scientifique. Rosenblueth et Wiener (1945) définissent la modélisation comme suit : « l’intention et le résultat d’une enquête scientifique sont d’obtenir une compréhension et un contrôle d’une partie de l’univers. Aucune partie substantielle de l'univers n'est si simple qu'elle puisse être saisie et contrôlée sans abstraction. L'abstraction consiste à remplacer la partie de l'univers considérée par un modèle de structure similaire, mais simplifié. Les modèles, formels ou intellectuels d'une part, ou matériels de l'autre, sont donc une nécessité centrale de la démarche scientifique ». En d’autres termes, la modélisation permet d’étudier un système complexe en le simplifiant. Par conséquent, les modèles fournissent des moyens d’appliquer les connaissances, d’exprimer la théorie et de faire avancer la compréhension de systèmes complexes.
Les modèles en nutrition animale et dans plusieurs domaines se caractérisent par trois principaux points (Sauvant, 1992):
− Statique ou dynamique. Les modèles statiques sont indépendants ou ne varient pas en
fonction du temps contrairement aux modèles dynamiques qui montrent l’évolution d’un système dans le temps;
− Empirique ou mécaniste. Les modèles empiriques décrivent les propriétés externes des
systèmes contrairement aux modèles mécanistes (explicatifs) qui permettent la traduction des propriétés d’un système via l’application de la connaissance de certaines de ses caractéristiques internes;
18
− Déterministe ou stochastique. Les modèles déterministes sont basés sur des paramètres
connus contrairement aux modèles stochastiques (probabilistes) qui s’appuient en partie sur des paramètres représentés par des variables aléatoires.
Les modèles dynamiques sont les plus utilisés en agriculture pour décrire divers phénomènes s’échelonnant de quelques jours à plusieurs semaines (courbes de croissance, vitesse d’élimination, etc.) (Thornley et France, 2007). Les modèles statiques peuvent être utilisés, par exemple, pour estimer la variation de la consommation alimentaire en fonction des caractéristiques de poids et/ou de production (Sauvant, 1992). Les modèles empiriques utilisent directement les données expérimentales pour quantifier les relations et sont basés principalement à un seul niveau (par exemple l’animal entier ou l’organisme) dans l’organisation des différents niveaux présentée plus haut (Tableau 1.1; France et Kebreab, 2008). La modélisation empirique décrit la réponse d’un système à l’aide d’équations mathématique simple de type entrée-sortie ou boîte noire. Contrairement aux modèles empiriques, les modèles mécanistes fournissent un degré de compréhension ou d’explication des phénomènes modélisés et sont construits en prenant en considération au moins deux niveaux (par exemple l’organisme et les organes; Thornley et France, 2007). La modélisation mécaniste définit à l’aide d’équations relatives aux propriétés des systèmes sous-jacents le comportement d’un système de niveau supérieur. Ce type de modèle utilise la notion de compartiments tissulaires et métaboliques ainsi que des voies métaboliques (flux) reliant les différents compartiments permettant de bien illustrer les phénomènes nutritionnels (Sauvant, 1992).
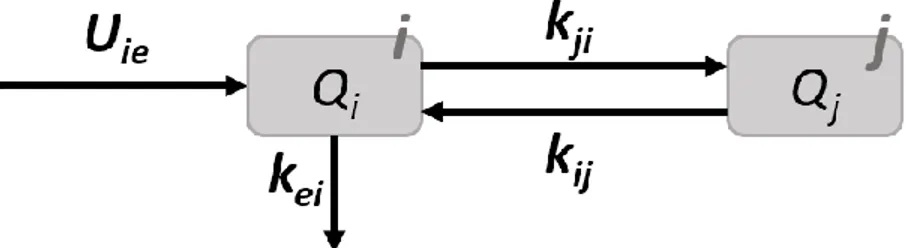
Le principe de base de cette approche est de considérer que la variation de la quantité d’un nutriment (𝑑𝑄𝑖) présent en quantité 𝑄𝑖 dans un compartiment i, et ce, lors d’une courte durée (𝑑𝑡) est la
différence entre la somme des flux entrants (∑ 𝐹𝑒) et sortants (∑ 𝐹𝑠) (Éq. 1.1; Sauvant, 1992). 𝑑𝑄𝑖
𝑑𝑡
⁄ = ∑ 𝐹𝑒− ∑ 𝐹𝑠 (Éq. 1.1)
L’équation précédente détermine la quantité d’un nutriment. Il est également important de déterminer le flux d’écoulement de sortie de ce même nutriment. Ce flux d’écoulement (𝑘 ∗ 𝑄𝑖, masse/temps) est directement proportionnel à la quantité 𝑄𝑖 déjà présente dans le compartiment au même instant et sort
19 𝑑𝑄𝑖
𝑑𝑡
⁄ = −𝑘 ∗ 𝑄𝑖 (Éq. 1.2)
Cette approche appliquée à un système à plus d’un compartiment et ouvert sur l’extérieur se présente comme suit (Éq. 1.3.; Sauvant, 1992;Figure 1.6):
𝑑𝑄𝑖 𝑑𝑡 ⁄ = ∑ 𝑈𝑖 𝑖𝑒 + ∑ 𝑘𝑗 𝑖𝑗 ∗ 𝑄𝑗 − ∑ 𝑘𝑖 𝑗𝑖 ∗ 𝑄𝑖 − ∑ 𝑘𝑖 𝑒𝑖∗ 𝑄𝑖 (Éq. 1.3) Où, 𝑑𝑄𝑖 𝑑𝑡
⁄ : variation de la quantité de nutriment dans le temps dans le compartiment i
𝑈𝑖𝑒 : flux d’entrée au compartiment i en provenance de l’extérieur
𝑘𝑖𝑗 : taux de sortie du compartiment j vers le compartiment i 𝑘𝑗𝑖 : taux de sortie du compartiment i vers le compartiment j 𝑘𝑒𝑖 : taux de sortie du compartiment i vers l’extérieur
𝑄𝑖, 𝑄𝑗 : quantité de nutriment présente respectivement dans les compartiments i et j
1.1.4.4 Méta-analyse
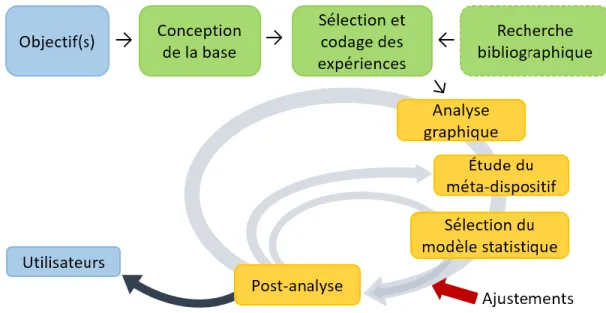
Les principes de méta-analyse présentés dans cette section ont été tirés de Sauvant et al. (2005, 2008). La méta-analyse est pertinente lorsque le nombre de données est suffisant et que la nécessité de comptabiliser et d’établir des lois de réponse générales est présente. Elle peut également être utilisée, entre autres, pour la modélisation mécaniste pour obtenir une équation avec autant de variables dépendantes souhaitées pour le calcul d’un flux. Dans le même ordre d’idées, cet outil statistique peut aider à la détermination de facteurs influents permettant d’estimer une variable. En d’autres termes, la méta-analyse, par sa capacité de synthèse, peut déceler des relations entre diverses variables. Les principales étapes d’une méta-analyse sont présentées à la Figure 1.7.
Figure 1.6. Variation de la quantité de nutriment dans le temps entre deux compartiments (Éq. 1.3)
20
Figure 1.7. Les principales étapes d'une méta-analyse (adaptée de Sauvant et al. (2005))
La première étape consiste à la définition du ou des objectifs de la méta-analyse qui permettra de mieux définir les étapes suivantes. Par la suite, la base de données peut être conçue en fonction des objectifs préalablement établis et tout ce qui est pertinent pour la méta-analyse doit être inclus dans cette base. Une recherche bibliographique est faite et il est important de bien définir les mots clés utilisés afin d’aller chercher le plus d’articles pertinents possible. Selon les objectifs établis et les critères de sélection choisis (ex. : type de ration, sans additifs alimentaires, etc.), un tri des expériences est fait et les expériences sélectionnées sont codées. Il faut d’abord coder les publications et, plus précisément, les expériences à l’intérieur d’une même publication. Le codage est un élément important avant même de débuter la méta-analyse puisque ce dernier permet d’éviter l’interprétation des ensembles de données au sein desquelles plusieurs objectifs expérimentaux ont été confondus. Après la sélection des expériences et leur codage, une analyse graphique de la ou les variables dépendantes (Y) en fonction des variables indépendantes (X) ainsi que les X en fonction des X (détection de corrélation entre les variables explicatives) est effectuée. Cette analyse graphique permet d’avoir une première idée de la dispersion des données, des relations intra- et inter-expériences ainsi que des hypothèses pour le choix d’un modèle statistique. Cette démarche graphique permet également de détecter les valeurs aberrantes ou les tendances présentes dans la base de données. Une étude du méta-dispositif expérimental est faite. Le méta-dispositif est déterminé par la structure des expériences
21
pour chaque variable explicative. Plusieurs étapes doivent être appliquées avant et après l’analyse statistique et ces étapes dépendent du nombre de variables explicatives et du type de méta-design. Dans le cas d’une seule variable explicative, un histogramme présentant la répartition de la variable explicative permet d’identifier les zones de valeurs sur lesquelles les chercheurs ont axé leur recherche et dont les expériences sont plus nombreuses. Il est également important de s’assurer de la présence ou non d’un effet expérience significatif et d’utiliser le bon traitement statistique. Dans le cas de deux variables explicatives ou plus, en plus des étapes mentionnées dans le cas d’une seule variable explicative, il est important de s’assurer de la non-colinéarité entre deux variables explicatives (X) via l’analyse graphique des X deux par deux. À la suite de l’analyse de la base de données, vient le choix du modèle statistique. Ensuite, il faut définir si l’effet expérience est aléatoire ou fixe, sachant que ce sujet fait débat tant dans les sciences médicales qu’en science animale. En générale, on peut considérer que l’effet expérience est aléatoire si chaque expérience est un échantillon pris au hasard au sein d’une population plus large et unique. Cet effet expérience est utilisé pour contrôler l’effet de la variabilité d’échantillonnage aléatoire présent entre les différentes expériences. L’effet expérience fixe est applicable lorsque chaque expérience est considérée comme issue d’une population différente et les expériences peuvent être supposées comme un choix arbitraire par les chercheurs. Le type de modèle statistique dépend de plusieurs facteurs. Ici, le modèle présenté (Modèle avec variables continues, Éq. 1.4; Sauvant et al., 2005) est celui qui a été utilisé pour la méta-analyse de la digestibilité iléale du P (Chapitre 2). Le modèle est celui avec des variables continues (quantitatives) et est décrit par la formule suivante :
𝑌𝑖𝑗 = 𝐵0+ 𝑆𝑖+ 𝐵1𝑋𝑖𝑗+ 𝑏𝑖𝑋𝑖𝑗 + 𝑒𝑖𝑗 (Éq. 1.4)
Où,
𝑌𝑖𝑗 : variable expliquée, traitement j de l’expérience i 𝐵0 : ordonnée à l’origine générale considérée en effet fixe
𝑆𝑖 : ordonnée à l’origine aléatoire des expériences i 𝐵1 : coefficient de régression général Y sur X (effet fixe) 𝑋𝑖𝑗 : variable explicative continue (quantitative)
𝑏𝑖 : effet aléatoire de l’expérience i sur le coefficient de régression de Y sur X
22
À la suite du choix du type de modèle à utiliser, des post-analyses sont nécessaire pour s’assurer que le modèle respecte tous les postulats. Tout d’abord, les résidus doivent suivre une loi normale. Tomassone et al. (1983) proposent de considérer les points ayant des écarts « studentisés » de plus de 3 ou plus (dépendamment du nombre d’observations) comme aberrants. Il est à noter qu’avant de retirer un point de la méta-analyse, une évaluation des conséquences engendrées par le retrait du ou des points doit être faite. En effet, le retrait de points ou de traitements jugés comme aberrants peut mener au retrait complet de l’expérimentation. Par la suite, une étude à posteriori du dispositif peut être faite afin de mesurer le poids des différents traitements dans la détermination des valeurs des paramètres des modèles ajustés. Les différentes étapes qui mènent à la sélection d’un modèle statistique final peuvent être répétées plusieurs fois et le modèle peut subir plusieurs ajustements. Lorsque le modèle est satisfaisant et évolue de façon logique, il peut être intégré dans un modèle mécaniste comme ce sera le cas au chapitre 3. Cependant, quand on veut faire un modèle de prédiction qui sera fourni à des utilisateurs, des étapes de validation et d’analyse de sensibilité sont importantes à réaliser.
Un aspect important dans l’utilisation d’un modèle statistique est de considérer que l’intervalle de prédiction du modèle doit être le même que celui des données utilisées pour faire la méta-analyse. Par exemple, si le modèle pour prédire le P digestible a été établi avec des essais présentant des valeurs de Ca alimentaire entre 2 et 10 g/kg, le modèle ne peut prédire le P digestible qu’avec des valeurs similaires de Ca alimentaire. Une étape importante avant l’utilisation des modèles est l’évaluation de leur capacité prédictive à l’aide de base de données dont les résultats sont comparés avec les prédictions du modèle pour en évaluer la précision. On peut ainsi déterminer la capacité d’extrapoler ou non le modèle à différents contextes.
1.2. Phosphore et développement durable
Le P est un nutriment essentiel pour les sols, les plantes et les animaux. Cependant, il est nécessaire qu’un équilibre soit atteint en apportant les quantités nécessaires aux besoins des cultures réceptrices. Bien que le règlement sur les exploitations agricoles (REA) exige d’avoir un bilan P équilibré depuis juin 2003 (MELCC, 2017), les épisodes d’algues bleues-vertes sont toujours d’actualité. De plus, les phosphates utilisés pour supplémenter les aliments des monogastriques en P présente un impact environnemental et économique puisque ces derniers sont sujets à des variations de prix et sont une
23
ressource non-renouvelables avec un épuisement des réserves mondiales estimé entre 50 et 100 ans selon les scénarios (Cordell et al., 2009).
1.2.1. Apports vs besoins
Les dernières recommandations en P chez les poulets de chair remontent à près de 25 ans (NRC, 1994). La production avicole n’est plus ce qu’elle était. Les techniques d’élevage, les équipements ainsi que la génétique des volailles ont subis de grandes améliorations aux cours des dernières années. Cependant, la formulation des aliments en P n’a pas évolué. Considérant les problèmes environnementaux associés aux excès de P et les performances bien différentes, il est nécessaire de revoir les besoins en P des poulets de chair. Selon Li et al. (2017), les besoins réels en P des poulets de chair d’aujourd’hui sont inférieurs à ceux présentés dans le NRC (1994). Une diminution de l’apport en P doit cependant être effectuée de façon prudente. Létourneau-Montminy et al. (2008) ont montré qu’une réduction du PNP chez les jeunes poulets de chair est possible si une réduction du Ca alimentaire est conjointement réalisée afin de prévenir l'effet négatif du Ca sur la disponibilité du P. Au contraire, dans certains contextes où des niveaux élevés de P peuvent être fournis, par exemple lors de l’utilisation de drêches de distillerie de maïs, il serait important de maintenir des apports de Ca élevés pour éviter que ce P soit excrété par manque de Ca. Donc, l’optimisation du P dans les aliments ne peut être faite sans prendre en considération le Ca alimentaire.
1.2.2. Calcium
Le Ca alimentaire a un effet négatif sur la digestibilité iléale du P (Li et al., 2018). Une formulation adéquate en Ca des aliments destinés aux poulets de chair pourrait donc permettre une meilleure utilisation de P. Cependant, ce n’est pas que le niveau de Ca alimentaire qui affecte la digestibilité de P, mais également les aspects qualitatifs de la source de Ca. La source principale de Ca utilisée en nutrition animale est d’origine minérale, soit des phosphates calciques ou du calcaire qui sont majoritairement composé de carbonate de calcium. La taille des particules de calcaire varie grandement de moins de 75 µm à plus de 800 µm et la solubilité du Ca dépend de ce facteur. Un calcaire plus fin libère plus de Ca2+ plus rapidement qu’un calcaire plus grossier. Non seulement la
taille des particules varie, mais également la pureté des calcaires est sujette à une variation (Li, 2018). En effet, la source minérale de Ca n’est pas composée à 100% de Ca, elle peut contenir d’autres minéraux (ordre décroissant d’affinité pour les phytates à pH de 7 : Zn2+ >> Fe2+ > Mn2+ > Fe3+ > Ca2+
24
engendrer une perte d’efficacité de la phytase. La solubilité du Ca influence donc sa capacité à se lier aux phytates. Ainsi, le Ca d’un calcaire plus fin et donc soluble plus rapidement crée des complexes Ca-phytate plus rapidement ne laissant pas le temps à la phytase d’hydrolyser le PP et réduisant ainsi l’efficacité de la phytase à libérer du P digestible (Kim et al. (non-publié) cité de Li, 2018). L’augmentation du pH dans le tractus digestif favorise la formation des complexes Ca-phytate (Angel et al., 2002). Donc, les complexes Ca-phytate se formeraient au niveau de l’intestin et impacteraient négativement la phytase endogène. La phytase microbienne agissant principalement au niveau du tractus digestif supérieur ne devrait donc pas être affectée par les complexes Ca-phytate. De plus, des concentrations élevées en Ca sont nécessaires pour impacter négativement l’hydrolyse de PP à pH acide. Cependant, les autres minéraux ayant une affinité plus élevée aux phytates que le Ca sont nécessaires en moins grande quantité pour avoir un effet à bas pH (Maenz et al., 1999). Plusieurs contaminants peuvent être retrouvés à l’intérieur des carbonates dont les minéraux comportant les cations ayant le plus d’affinité pour les phytates : 200 à 6000 mg/kg de fer sous la forme de Fe2O3, 20
à 1000 mg/kg de manganèse sous la forme MnO2 et 3 à 500 mg/kg de zinc (Oates, 1998). Ainsi, un
effet négatif sur l’efficacité de la phytase à libérer du P serait à tort attribué au Ca provenant en réalité de l’impureté (présence d’autres minéraux) de la source de Ca.
1.2.3. Déplétion-Réplétion
Le métabolisme phosphocalcique fait l’objet de régulations hormonales permettant le maintien des concentrations plasmatiques (voir section 1.1.3.2). Ainsi, en cas de restriction alimentaire en Ca et en P assez importante pour diminuer les réserves osseuses (déplétion), l’utilisation du P alimentaire par les animaux pourrait être améliorée. L’effet d’une période de déplétion suivie d’une période de réplétion a été étudié chez les poulets de chair. Rousseau et al. (2016) et Yan et al. (2005) ont montré que les poulets de chair ont la capacité de s’adapter à des niveaux bas de Ca et P en bas âge en augmentant leur efficacité digestive. Ce processus d’adaptation engendre une augmentation de l’expression de certains gènes codant les transporteurs intestinaux de P et de Ca permettant ainsi aux oiseaux d’être plus efficaces à absorber le P et le Ca. Cette capacité d’adaptation et l’étendue de la compensation en termes de performances et de minéralisation osseuse dépend cependant des taux de Ca et de P dans le traitement dit de ‘’réplétion’’ qui suit celui de déplétion (Rousseau et al., 2016). La même stratégie peut être utilisée pour les oiseaux en phase de croissance sans affecter les performances de croissance finales à condition que l’aliment de finition apporte suffisamment de Ca et de P pour
25
rattraper le déficit de minéralisation osseuse induit par la déplétion (Valable et al., 2018). Compte tenu que la consommation de P est plus importante en phase de finition, l’idée serait de rendre les oiseaux plus efficaces par des carences en bas âge leur permettant ainsi de recevoir des aliments réduits en minéraux en phase de finition. Une stratégie de déplétion-réplétion en P et en Ca permettrait donc de réduire l’utilisation de P alimentaire en conservant des performances de croissance similaires à ceux recevant des aliments conventionnels.
1.2.4. Alimentation de précision
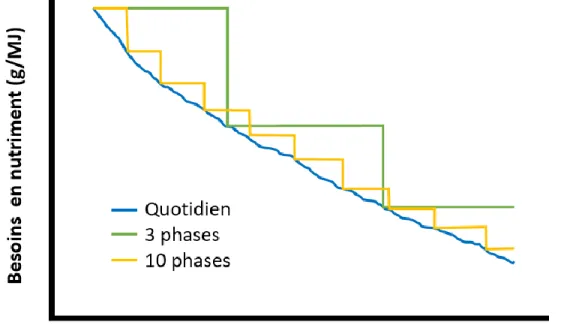
Classiquement, les animaux sont alimentés par phase en fonction de leur poids ou âge compte tenu que leurs besoins diminuent dans le temps lorsqu’exprimés par kg d’aliment. Chez le poulet, une alimentation en 3 phases est généralement pratiquée, par exemple, un aliment de 0-10 jours, un de 11-21 jours et enfin un de 22-35 jours d’âge. Les besoins des oiseaux au début de chaque phase sont utilisés pour formuler les aliments engendrant ainsi de larges excès pour les oiseaux plus âgés vers la fin de la phase. Ainsi, plus on augmente le nombre de phases, plus les excès sont réduits (Figure 1.8).
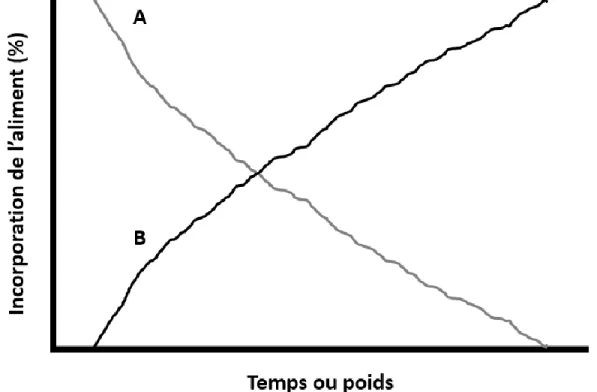
Cependant, la gestion d’un tel nombre d’aliments n’est pas viable économiquement. Une alimentation «multiphase» devient intéressante puisque l’ajustement des nutriments se fait via la combinaison de deux aliments ou pré-mélanges permettant d’atteindre le niveau adéquat des nutriments (Buteri et al.,
Figure 1.8. Concentration en nutriment distribué à des animaux selon un système d’alimentation quotidien, 3 phases ou 10 phases (adaptée de Pomar et al. (2009))