THESE
THESE
En vue de l'obtention du
DOCTORAT DE L’UNIVERSITÉ DE TOULOUSE
DOCTORAT DE L’UNIVERSITÉ DE TOULOUSE
Délivré par l'Université Toulouse III - Paul Sabatier Discipline ou spécialité : Ethologie
JURY
Professeur Jean-Louis Deneubourg Professeur Philippe Gaussier Docteur Nicolas Franceschini Professeur Dario Floreano Professeur Richard Fournier
Ecole doctorale : CLESCO Unité de recherche : CRCA
Directeur(s) de Thèse : Dr Guy Theraulaz
Rapporteurs : Professeur Jean-Louis Deneubourg et Professeur Philippe Gaussier
Présentée et soutenue par Simon Garnier Le 19/11/2008
Simon Garnier Tous droits réservés
Remerciements
En tout premier lieu, parce que la tradition l’exige certes, mais surtout parce qu’il le mérite, je tiens à remercier mon directeur de thèse, Guy Theraulaz, pour m’avoir donner l’opportunité de pratiquer le métier de chercheur, métier qui me tient tant à coeur. Au cours de ces années passées à travailler sous ta direction, j’ai pu mesurer à de nombreuses reprises la chance que j’ai eu de débuter ma carrière à tes côtés. Tu t’es toujours efforcé de me fournir le matériel et les ressources nécessaires à la bonne conduite de mes travaux. Tu m’as encouragé à présenter mes résultats dans des conférences nationales et internationales. A travers ton réseau de relations, tu m’as permis d’établir des collaborations multiples dans des domaines aussi divers que l’éthologie, la robotique, l’informatique, la physique ou les mathématiques. Ces collaborations ont enrichi considérablement mon travail quotidien et l’ont rendu plus passionnant que jamais. Enfin, tu as toujours su être présent pour discuter, pour me conseiller et pour me transmettre tes immenses connaissances sur l’organisation des sociétés animales. Mon impulsivité n’a pas dû te rendre la tâche facile tous les jours, et ta patience à mon encontre n’en est que plus méritoire. Pour toutes ces raisons et bien d’autres encore, je t’adresse de nouveau un grand merci.
Au cours de ces années, Christian Jost a été successivement mon professeur, mon directeur de DEA et mon référent pédagogique. Il s’est acquitté de toutes ces tâches avec la ferveur qui caractérise l’excellent enseignant qu’il est. C’est lui qui m’a appris (presque) tout ce que je sais aujourd’hui sur l’analyse statistique et la modélisation en biologie. C’est lui qui m’a mis le pied à l’étrier en m’offrant l’opportunité de réaliser mon premier stage dans le laboratoire lors de mon année de Maîtrise. C’est lui enfin qui m’a transmis l’amour de l’enseignement malgré mes fortes réticences initiales. Pour ta disponibilité, pour ta gentillesse et pour tes inimitables « suisseries », merci beaucoup Christian.
Je tiens à remercier Jean-Louis Deneubourg et Philippe Gaussier d’avoir accepté d’évaluer mon manuscrit. Pour avoir évaluer quelques copies d’examens, j’imagine à quel point la fonction de rapporteur peut être gourmande en temps. Merci d’avoir bien voulu consacrer le vôtre à mon travail de thèse. Je vous associe également aux remerciements que j’adresse à Dario Floreano,
mon jury de thèse.
Ma famille a toujours été d’un grand soutien pour moi a cours de ces longues années d’étude. Avec des moyens financiers restreints, mais surtout avec beaucoup d’amour, mes parents ont toujours fait ce qu’il fallait pour que leurs enfants suivent la voie qu’ils s’étaient choisis. J’en suis arrivé là aujourd’hui parce qu’ils ne m’ont jamais laisser penser que c’était impossible. Autour d’eux, mes frères et soeurs, mes grand-parents, mes oncles, mes tantes et tous mes cousins (et ils sont nombreux) ont entretenu un climat de bonne humeur permanente, dans les moments les plus joyeux comme au coeur des coups durs. Leur verve et leur bagou m’ont appris à me sortir des joutes orales les plus difficiles, leur entêtement et leur force de caractère m’ont appris à ne jamais renoncer. A vous tous qui êtes de ma famille, un grand merci.
Comme ma famille, mes amis prennent une part importante dans la réussite de mon travail. Ils ont enrichi ma vie de leurs expériences et, pour cela, je remercie toutes ces personnes qui ont croisé plus ou moins longtemps mon chemin ces dernières années. Parmi elles, je voudrais adresser un merci spécial et chaleureux à Alexandre, Mehdi et Marie-Hélène qui ont partagé avec moi la passion de l’étude des sociétés animales et m’ont apporté aide, soutien et bonne humeur. Vous avez été les meilleurs collègues possibles pendant ces quatre années et vous resterez mes amis pour un long moment encore. Je tenais également à remercier les « p’tits loups » et les « p’tits monstres » du hand qui m’ont permis de me défouler dans tous les gymnases de la région et avec qui j’ai partagé mes plus beau fous rire. Merci aussi à Nathalie pour tout ce qu’elle donne aux autres, merci à Jeanne pour son éternelle joie de vivre. Merci Lucile, Carole et Elodie. Vous avez partagé ma vie et m’avez offert les moments les plus heureux de mon existence.
La recherche scientifique est un travail d’équipe et rien de ce qui est dans ce manuscrit n’aurait pu voir le jour sans de multiples collaborations, conseils et coups de main. Merci donc à tous les membres de l’équipe EMCC, titulaires et passagers, qui ont toujours su m’apporter leur aide dans mon entreprise. Merci à Vincent, Raphaël, Pablo, Richard, Andrea, Jacques, Laurent, Grégory, Niriaska, Sébastien et Abel pour leur disponibilité et leurs conseils. Merci également à Audrey, il paraît que tu n’y es pas pour rien dans mon arrivée au sein de l’équipe. Merci à Aurélie et à Marjorie, les meilleures stagiaires du monde. Merci à Anne pour les heures de travail qu’elle a consacré à mes travaux, merci à Gérard pour ses connaissances encyclopédiques sur l’élevage des fourmis, merci à Maud pour tous les logiciels qu’elle développe pour nos besoins spécifiques. Je remercie Gilles, Masoud, Fabien, Nikolaus et tous les membres des laboratoires de Roland Siegwart et Alcherio Martinoli pour l’aide qu’ils m’ont apporté dans mon travail de robotique
L’approche multi-échelle et multi-disciplinaire que vous développez est une source de richesse scientifique qu’il est essentiel de préserver, la place privilégiée que vous accordez aux étudiants est un moteur d’innovation rare et précieux.
Table des matières
Remerciements v
1 Introduction 1
1.1 Intelligence en essaim . . . 5
1.1.1 Introduction et repères historiques . . . 5
1.1.2 Auto-organisation et propriétés des systèmes d’intelligence en essaim . . . 9
1.1.2.1 Mécanismes . . . 10
1.1.2.2 Propriétés . . . 11
1.1.2.3 Etude des systèmes d’intelligence en essaim . . . 14
1.1.3 Interactions et transferts d’information dans les systèmes d’intelligence en essaim . . . 15
1.1.3.1 Transfert indirect d’information . . . 15
1.1.3.2 Transfert direct d’information . . . 17
1.1.3.3 Direct vs indirect . . . 19
1.1.4 Fonctions assurées par les systèmes d’intelligence en essaim . . . 21
1.1.4.1 Coordination . . . 21
1.1.4.2 Collaboration . . . 22
1.1.4.3 Délibération . . . 23
1.2 Intelligence en essaim dans les systèmes artificiels . . . 23
1.2.1 De l’animal à l’animat . . . 24
1.2.1.1 Intelligence artificielle . . . 24
1.2.1.2 Les Animats . . . 25
1.2.1.3 De l’animat à l’animal . . . 27
1.2.2 Applications de l’intelligence en essaim . . . 32
1.2.2.1 Animation graphique : les Boids et leurs descendants . . . 32
1.2.3 Robotique collective . . . 36 1.2.3.1 Agrégation . . . 37 1.2.3.2 Ségrégation . . . 40 1.2.3.3 Exploration . . . 41 1.2.3.4 Déplacements coordonnés . . . 43 1.2.3.5 Manipulations coopératives . . . 44 1.2.3.6 Allocation de tâches . . . 44
1.2.3.7 Prises de décisions collectives . . . 46
1.3 Décisions collectives dans les systèmes biologiques d’intelligence en essaim . . . . 46
1.3.1 Prises de décisions dans les sociétés animales . . . 46
1.3.1.1 Qui décide dans le groupe ? . . . 47
1.3.1.2 Comment les individus s’accordent-ils sur un choix ? . . . 50
1.3.2 Décisions collectives et intelligence en essaim . . . 51
1.3.2.1 Sélection d’un chemin chez la fourmi . . . 52
1.3.2.2 Les points à retenir de l’exemple des fourmis . . . 53
1.3.2.3 Sensibilité à l’environnement . . . 55
1.3.2.4 Préférences individuelles et choix collectifs . . . 56
1.3.2.5 Modulation des comportements individuels et efficacité des choix collectifs . . . 57
1.3.2.6 Complexité individuelle vs complexité collective . . . 59
1.4 Objectifs . . . 64
2 Implémentation du comportement d’agrégation de la blatte Blattella germa-nica dans un groupe de micro-robots 69 2.1 Introduction . . . 72
2.2 Materials and methods . . . 74
2.2.1 The biological system : first-instar larvae of Blattella germanica . . . 74
2.2.2 The artificial system : micro-robots Alice . . . 74
2.2.3 Experimental set-up . . . 74
2.2.4 The behavioral model . . . 75
2.2.5 Implementation in the Alice robots . . . 78
2.2.5.1 Displacement . . . 78
2.3 Analysis and Comparison to Cockroach behavior . . . 81
2.3.1 Path analysis for an individual robot . . . 81
2.3.1.1 Central zone . . . 81
2.3.1.2 Peripheral zone . . . 84
2.3.1.3 Spontaneous stopping times . . . 84
2.3.2 Interactions among robots . . . 84
2.3.2.1 Probabilities to join and to leave an aggregate . . . 84
2.3.2.2 Calibration of interaction parameters . . . 85
2.3.3 Collective behaviors . . . 86
2.4 Discussion . . . 87
2.5 Conclusion . . . 92
3 Le comportement d’agrégation des blattes comme processus de décision col-lective pour des groupes de robots 95 3.1 Introduction . . . 98
3.2 Self-organized aggregation . . . 100
3.3 Collective choice . . . 101
3.4 Conclusion . . . 107
3.5 Introduction . . . 110
3.6 Material and methods . . . 114
3.6.1 The micro-robots Alice . . . 114
3.6.2 The experimental set-up . . . 115
3.6.3 The behavioral model . . . 115
3.6.4 The experimental parameters . . . 116
3.6.4.1 Identical shelters . . . 116
3.6.4.2 Shelters of different sizes . . . 116
3.6.5 Numerical experiments . . . 117 3.6.6 Data analysis . . . 118 3.6.6.1 Experiments . . . 118 3.6.6.2 Sensitivity analysis . . . 119 3.7 Results . . . 120 3.7.1 Identical shelters . . . 120 3.7.1.1 Experiments . . . 120
3.7.1.2 Simulator validation . . . 121
3.7.1.3 Sensitivity analysis . . . 121
3.7.1.4 Summary of the identical shelters case . . . 124
3.7.2 Shelters with different sizes . . . 124
3.7.2.1 Experiments . . . 124
3.7.2.2 Simulator validation . . . 126
3.7.2.3 Sensitivity analysis . . . 128
3.7.2.4 Summary of the different shelters case . . . 129
3.8 Discussion . . . 130
4 Sensibilité de la fourmi d’Argentine à la géométrie des bifurcations d’un réseau143 4.1 Introduction . . . 146
4.2 Methods . . . 148
4.2.1 Biological material . . . 148
4.2.2 Experimental set-up and protocol . . . 148
4.2.3 Statistical analysis . . . 151
4.3 Results . . . 151
4.3.1 Foodbound trip (unfed ants) . . . 151
4.3.2 Foodbound trip (unfed ants) . . . 152
4.4 Discussion . . . 154
5 Sélection de chemin et efficacité de fourragement dans un réseau de transport chez la fourmi d’Argentine 159 5.1 Introduction . . . 162
5.2 Material and methods . . . 165
5.2.1 Biological material . . . 165
5.2.2 Individual behaviour . . . 165
5.2.3 Collective behaviour . . . 168
5.3 Experimental results . . . 170
5.3.1 Individual ant behaviour . . . 170
5.3.2 Collective behaviour . . . 171
5.4 Model . . . 172
5.4.1 Model description . . . 172
5.5 Discussion . . . 176
6 Sélection de chemin par stigmergie dans un groupe de petits robots autonomes185 6.1 Introduction . . . 188
6.2 Materials and methods . . . 190
6.2.1 Robot Alice . . . 190
6.2.1.1 Base robot . . . 190
6.2.1.2 Trail following add-on . . . 190
6.2.2 Experimental setup . . . 191
6.2.2.1 Maze . . . 191
6.2.2.2 Tracking device . . . 194
6.2.2.3 Pheromone deposit device . . . 194
6.2.3 Behavioral model . . . 195
6.2.4 Simulations . . . 196
6.3 Data analysis and results . . . 196
6.3.1 Parameter estimation . . . 196
6.3.2 Two path maze . . . 198
6.3.2.1 Identical length condition . . . 198
6.3.2.2 Different length condition . . . 199
6.3.3 Network maze . . . 199
6.3.3.1 Three loop maze . . . 200
6.3.3.2 Four loop maze . . . 200
6.4 Discussion . . . 202
6.5 Conclusion . . . 204
7 Discussion et conclusion 207 7.1 Discussion générale . . . 208
7.1.1 Comparaison qualitative des deux processus de décision collective . . . 208
7.1.1.1 Interactions directes vs interactions indirectes . . . 208
7.1.1.2 Sélection de place vs sélection de route . . . 209
7.1.1.3 Sensibilité à la taille du groupe . . . 209
7.1.1.4 Conclusion . . . 211
7.1.2 Implémentation du modèle biologique : objectifs et méthodologie . . . 211
7.1.2.2 L’approche bio-inspirée . . . 213
7.1.2.3 Conclusion . . . 213
7.1.3 Structure de l’environnement, modulation passive du comportement et par-cimonie du traitement cognitif . . . 214
7.1.3.1 Structure de l’environnement et modulation passive du compor-tement . . . 214
7.1.3.2 Structure de l’environnement et parcimonie du traitement cognitif 216 7.2 Perspectives . . . 217
7.2.1 Formation des réseaux de pistes chez la fourmi d’Argentine . . . 217
7.2.2 Détection collective de caractéristiques environnementales . . . 219
7.2.3 Quel avenir pour la robotique en essaim ? . . . 221
7.2.3.1 L’absence d’applications . . . 221
7.2.3.2 Polymorphisme, polyéthisme et modulation des comportements individuels . . . 222
7.2.3.3 Intégration dans les systèmes naturels . . . 223
7.3 Conclusion . . . 224
Bibliographie 227
A Les principes biologiques de l’intelligence en essaim 257
B Transferts d’information et comportements auto-organisés dans les foules et
les essaims 289
C Extraction expérimentale de lois d’interaction chez l’Homme : le cas des
Chapitre 1
Introduction
Dans la nature, les groupes et les sociétés animales sont fréquemment confrontés à des choix dont peut dépendre leur survie. Les singes hamadryas par exemple se dispersent tous les matins pour rechercher de la nourriture. Avant cela, ils décident autour de quel point d’eau ils se retrouve-ront vers le milieu de la journée pour se reposer (Kummer, 1968). Dans la région semi-désertique où ces animaux vivent, les points d’eau sont relativement rares. Le regroupement de l’ensemble des singes autour de l’un d’entre eux facilite sa défense contre d’éventuels compétiteurs, ainsi que la défense du groupe contre ses prédateurs. Tous les matins, ces singes doivent donc prendre une décision importante pour l’avenir du groupe. Pour cela, ils votent... “avec leurs pieds”, c’est à dire en se déplaçant. Pour indiquer leur point d’eau préféré, certains mâles s’éloignent de quelques mètres du groupe dans la direction de ce point d’eau. Les autres mâles “votent” en rejoignant l’un ou l’autre, et des colonnes commencent à se former en direction des points d’eau majori-taires. Les mâles peuvent également attirer l’attention de leurs congénères pour les inciter à les rejoindre dans leur colonne. Pendant plus d’une heure, ces colonnes se développent ou régressent en fonction des changements d’opinions des individus. Finalement, l’une d’entre elles l’emporte et l’ensemble du groupe se met alors en mouvement dans la direction choisie (Sigg & Stolba, 1981).
De manière générale, lorsqu’un groupe d’animaux fait face à un choix, l’ensemble des individus doit adopter de concert la même alternative afin de limiter les risques de séparation. Par exemple, lorsqu’une colonie d’abeilles mellifères essaime vers un nouveau nid, les membres de l’essaim doivent s’accorder à l’avance sur la localisation de leur futur nid parmi les multiples possibilités qui s’offrent dans l’environnement. Sans cet accord préalable, l’essaim encoure le risque de se disperser (Seeley & Buhrman, 1999). Les animaux doivent donc établir un consensus auquel tous les membres du groupe vont se rallier. Le choix par consensus se retrouve également dans les sociétés humaines et s’applique aussi aux systèmes artificiels conçus par l’Homme comme les
robots. Par exemple, lorsque plusieurs robots coopèrent pour transporter une charge importante vers un objectif, il est nécessaire que les robots s’entendent sur la direction à suivre ou la route à prendre pour atteindre cet objectif (Kube & Bonabeau, 2000; Campo et al., 2006). Dans un système naturel ou artificiel, il est donc essentiel que les agents qui le composent disposent de mécanismes leur permettant d’établir un consensus. On distingue principalement deux grandes catégories de mécanismes :
1. Dans la première catégorie, le processus de décision repose sur la centralisation de l’infor-mation acquise par le groupe. Ce regroupement de l’inforl’infor-mation peut avoir deux origines différentes. La centralisation peut être le fait d’un individu unique, un leader, ou d’un petit groupe d’individus qui se chargent de prendre la décision pour l’ensemble du groupe. Dans ce cas, les autres membres du groupe ne participent pas à la décision et se contentent de suivre les directives du leader ou du sous-groupe dirigeant. Mais l’information peut égale-ment être centralisée à travers l’organisation d’un vote. Dans ce cas, les opinions de chaque membre du groupe sont collectées et rassemblées pour être dénombrées. Le consensus est obtenu en suivant une règle établie au préalable, par exemple la règle de la majorité absolue, qui défini l’alternative gagnante en fonction des votes exprimés. L’ensemble des membres du groupe est alors tenu de suivre ce résultat, quelque qu’ait été son vote personnel. 2. Dans la seconde catégorie, l’information n’est jamais centralisée au cours du processus
de décision. Les individus se comportent en suivant leurs propres motivations. Ils ne se conforment pas à la décision d’un leader ni à une opinion majoritaire. Cependant, leur comportement est influencé par celui des congénères avec qui ils interagissent. Plus pré-cisément, ils tendent à adopter le même comportement qu’eux. A travers ce mimétisme comportemental, les individus s’accordent sans concertation sur une alternative à adopter. Ces mécanismes décentralisés permettant à des individus de prendre collectivement une décision s’observent fréquemment dans les colonies d’insectes sociaux. On les retrouve éga-lement chez de nombreuses autres espèces sociales, y compris chez l’Homme.
Le présent mémoire se veut une contribution à la connaissance et à la compréhension de ces mécanismes décentralisés de prise de décision dans les systèmes collectifs. D’un point de vue théorique, ce travail se situe dans le cadre général de l’étude des systèmes d’intelligence en essaim. Ce champ de recherche s’attache à comprendre et à concevoir des mécanismes décentralisés de résolution collective de problèmes, et s’intéresse aussi bien aux systèmes biologiques qu’aux systèmes artificiels (Bonabeau et al., 1999; Bonabeau & Theraulaz, 2000; Eberhart et al., 2001;
Garnier et al., 2007a). Il étudie plus particulièrement les comportements des individus et du groupe auquel ils appartiennent, et cherche avant tout à établir le lien causal qui les unit.
Dans la nature, les mécanismes décentralisés de résolution de problèmes sont à l’origine de certains des comportements collectifs les plus spectaculaires. De l’organisation spontanée du trafic piétonnier chez l’Homme à la construction de nids gigantesques chez les termites et les fourmis, de la répartition adéquate des tâches à accomplir chez les guêpes à la coordination du mouvement dans les vols d’oiseaux et les bancs de poissons, ces mécanismes permettent aux sociétés animales d’organiser de manière efficace l’activité de grands groupes d’individus.
Ils ont également servi de source d’inspiration pour la conception de plusieurs systèmes ar-tificiels. Ils sont par exemple à l’origine d’un système très efficace pour optimiser le routage des marchandises et le transfert des paquets de données dans les réseaux de télécommunication (Do-rigo et al., 1999; Bonabeau et al., 2000). Ils servent également dans l’animation graphique pour générer le mouvement de foules virtuelles et leur donner un aspect réaliste (Reynolds, 1987). Enfin, ils ont contribué au développement d’une nouvelle branche de la robotique qui s’en inspire pour coordonner l’activité de plusieurs robots afin qu’ils accomplissent collectivement une tâche (Sahin & Winfield, 2008).
Parmi tous ces phénomènes collectifs, les mécanismes décentralisés de prise de décision ont été l’objet de très nombreuses études en biologie. Ils ont également servi de sources d’inspira-tion pour concevoir de nouveaux algorithmes d’optimisad’inspira-tion dans le domaine de la recherche opérationnelle et en informatique. Cependant, leur application en robotique reste extrêmement marginale. A travers une approche combinant étroitement éthologie et robotique, expériences et simulations, ce travail se propose donc d’étudier deux processus biologiques permettant à un groupe de prendre une décision et de les implémenter sur une plateforme robotique.
Dans cette introduction, nous aborderons dans un premier temps le concept d’intelligence en essaim (Section 1.1). Nous commencerons par le définir à travers un rapide historique (Section 1.1.1) qui nous permettra d’en comprendre les multiples racines scientifiques. Cette section nous permettra également d’appréhender les questions abordées par ce champ de recherches. Puis nous nous intéresserons plus particulièrement aux processus d’auto-organisation qui sous-tendent le fonctionnement des systèmes d’intelligence en essaim (Section 1.1.2). En particulier, nous passerons en revue les mécanismes (Section 1.1.2.1) et les propriétés (1.1.2.2) de ces processus, ainsi que la méthodologie générale qui permet de les étudier (Section 1.1.2.3). Nous verrons que les processus d’auto-organisation reposent sur de multiples interactions entre les individus
composant le groupe. La nature de ces interactions sera abordée dans la Section 1.1.3. Nous distinguerons deux types d’interactions, les interactions indirectes (Section 1.1.3.1) et directes (Section 1.1.3.2), dont nous comparerons les propriétés (Section 1.1.3.3). Enfin, nous parlerons des fonctions que les systèmes d’intelligence en essaim remplissent pour le groupe (Section 1.1.4). Nous en différencierons trois en particulier : la coordination (Section 1.1.4.1), la collaboration (Section 1.1.4.2) et la délibération (ou prise de décision collective, Section 1.1.4.3).
Dans un second temps, nous passerons en revue les systèmes artificiels d’intelligence en es-saim, et en particulier ce qu’il est convenu d’appeler la robotique en essaim (Section 1.2). Nous commencerons par faire des rappels historiques sur la transition entre l’approche classique de l’intelligence artificielle, et l’approche adaptative (Section 1.2.1). Nous aborderons ces deux no-tions dans les Secno-tions 1.2.1.1 et 1.2.1.2. Nous discuterons également l’intérêt de l’approche adaptative pour la connaissance des systèmes biologiques (Section 1.2.1.3). Nous présenterons ensuite rapidement deux types d’applications de l’intelligence en essaim : l’animation graphique de foules (Section 1.2.2.1) et les algorithmes d’optimisation (Section 1.2.2.2). Nous nous focalise-rons ensuite sur l’application de l’intelligence en essaim dans le domaine de la robotique collective (Section 1.2.3).
Dans la troisième partie de cette introduction, nous aborderons en détails les processus de délibération par auto-organisation dans les sociétés animales (Section 1.3). Nous commencerons par replacer cette question dans un cadre théorique plus général, celui de la prise de décision par consensus. En particulier nous nous poserons les questions suivantes : qui dans un groupe d’ani-maux peut prendre une décision ? (Section 1.3.1.1) Et comment les anid’ani-maux s’accordent-ils sur un choix commun ? (Section 1.3.1.2) A l’intérieur de ce cadre théorique général, nous identifierons la place des processus de délibération par auto-organisation (Section 1.3.2). Puis à travers une série non exhaustive d’exemples, nous expliquerons les mécanismes qui permettent à un groupe d’in-dividus de prendre collectivement une décision en ne se basant que sur une information locale et restreinte de l’activité globale du groupe (Section 1.3.2.1 et 1.3.2.2). Nous évoquerons ensuite la sensibilité de ces mécanismes de délibération aux variations de l’environnement (Section 1.3.2.3) et aux préférences individuelles (Section 1.3.2.4). Nous parlerons également de l’effet des modu-lations du comportement des individus sur l’efficacité du processus de décision (Section 1.3.2.5). Pour finir, nous essaierons de comprendre comment la complexité comportementale de l’individu peut modeler précisément le processus de décision à l’échelle du groupe (Section 1.3.2.6).
Enfin, nous préciserons dans la dernière partie de cette introduction les objectifs de cette thèse ainsi que l’organisation générale du reste du mémoire (Section 1.4).
1.1 Intelligence en essaim
1.1.1 Introduction et repères historiques
L’étude du comportement animal présente à divers égards des aspects aussi intrigants que pas-sionnants. Parmi eux se trouvent les extraordinaires phénomènes collectifs (voir Figure ??) que l’on observe au sein de nombreuses sociétés animales (Camazine et al., 2001; Couzin & Krause, 2003; Sumpter, 2006). Tout le monde a un jour observé les surprenantes chorégraphies exécutées par des vols d’oiseaux ou des bancs de poissons, chaque membre du groupe en parfaite coordina-tion avec ses congénères au sein d’une structure constamment changeante et pourtant toujours cohérente. Moins virevoltant mais tout aussi impressionnant est le spectacle offert chaque année dans la savane africaine du Serengeti par la migration d’un troupeau gigantesque de gnous. Un million et demi de ces animaux, accompagnés par des milliers de zèbres et de gazelles, entament à chaque fin de saison humide un long périple collectif qui les mènera à leurs quartiers de saison sèche. Suivant les zones d’herbes fraiches, le troupeau géant forme un immense front s’étendant parfois sur plusieurs kilomètres (Gueron & A., 1993). Plus proches de nous, dans les rues bondées de nos centres-villes ou dans les couloirs de métro aux heures de pointe, les foules de piétons forment également d’étonnantes structures : les personnes provenant de directions opposées se partagent spontanément la route, créant ainsi des files alternées d’individus se déplaçant dans le même sens (Helbing et al., 2001).
Plus petits, plus discrets dans la nature, les insectes sociaux sont néanmoins maîtres dans l’art de l’organisation des foules, de la construction de structures gigantesques et de la résolution des problèmes à l’échelle de leur société toute entière (Bonabeau et al., 1997; Camazine et al., 2001; Detrain & Deneubourg, 2006). Leurs colonies, qui selon les espèces comptent quelques individus ou des millions, présentent une gamme très variée de comportements collectifs, qui allient l’efficacité à la flexibilité et à la robustesse. Ces animaux à l’apparence simple sont ainsi capables de construire des réseaux de transport et d’échange (Theraulaz et al., 2003; Buhl et al., 2004, 2005) à l’intérieur desquels le traffic des individus et du matériel est régulé (Couzin & Franks, 2003; Dussutour et al., 2004; Burd, 2006; Vittori et al., 2006) ; ils sont également à même de construire des édifices immenses comparés à leur petite taille (Grassé, 1984; Tschinkel, 2003, 2004) ; ils disposent de systèmes de division du travail permettant d’allouer de manière dynamique les différentes tâches à effectuer pour le bon fonctionnement de la colonie (Deneubourg et al., 1987; Gordon, 1996; Bonabeau et al., 1998b; Beshers & Fewell, 2001) ; etc.
ca-Figure 1.1: Exemples de comportements collectifs dans les sociétés animales et humaines. (a) Culture sur agar de la bactérie Bacillus subtilis. Le pattern formé par la colonie dépend à la fois du morphotype de la bactérie et des conditions environnementales (Ben-Jacob et al., 2000). (b) Colonne de chasse chez la fourmi légionnaire Eciton burchelli. Les individus au centre de la colonie se déplacent majoritairement en direction du bivouac de la colonie, alors que les individus sur les côtés se déplacent majoritairement vers le front de chasse. (c) Un banc de poissons formé par l’alignement des individus sur une direction commune de déplacement. (d) Un phénomène similaire est à l’origine de la formation des vols d’oiseaux. (e) Migration d’un troupeau géant de gnous à travers la savane du Serengeti. (f) Déplacement d’une foule de piétons dans une rue commerçante. Les individus se séparent en deux flux de directions opposées (repris depuis Helbing et al., 2005).
pables de former d’étonnantes structures spatio-temporelle (Ben-Jacob et al., 2000). Par exemple, lorsque les conditions environnementales deviennent rudes, en particulier quand les nutriments se font rares, l’amibe Dictyostelium discoideum produit des bouffées d’AMP cyclique qui, relayées de proche en proche par les autres cellules, produit des ondes en spirales. Pour finir, chez de nom-breuses espèces de bactéries, le déplacement coordonné des cellules au cours de la croissance de la colonie conduit à la formation de structures tri-dimensionnelles régulières aux formes multiples.
Depuis longtemps, ces étonnants comportements, ces colossales structures qui émergent au sein des sociétés animales fascinent les hommes. Artistes et naturalistes se sont demandés quelle force permettait à tant d’animaux de se coordonner afin d’atteindre les objectifs du groupe. Le poète belge Maurice Maeterlink par exemple prêtait aux insectes sociaux une forme d’obéissance à une entité, “un maître ignoré” qu’il appelait “l’esprit de la ruche” (Maeterlinck, 1927). Plus récemment, Michael Crichton reprend dans son roman, “La proie”, une idée assez similaire, celle d’une nuée de nanorobots formant une forme de super-organisme capable de prendre des décisions complexes et d’anticiper des événements futurs (Crichton, 2002).
De son côté, la littérature scientifique sur le sujet remonte au moins jusqu’au XIXème siècle. Les premières hypothèses pour expliquer ces phénomènes collectifs faisaient appel à une organi-sation hiérarchique et centralisée (voir par exemple Büchner, 1881; Forel, 1921). Les individus étaient supposés disposer de capacités cognitives suffisantes pour se représenter la globalité de l’état de la société et de l’environnement, pour évaluer la situation de la société par rapport à ses objectifs et pour prendre en conséquence des décisions appropriées (voir par exemple Thorpe, 1963). Dans ce schéma de pensée, la complexité de la structure produite ou du comportement réalisé par le groupe reflétait la complexité cognitive des individus composant le groupe, ou au moins de l’un d’entre eux. En particulier chez les insectes sociaux, la reine était considérée comme le superviseur de l’activité de la colonie, surveillant l’activité des ouvriers et leur donnant des ordres appropriés à chaque situation.
Cependant, une telle approche ne peut pas rendre compte d’un grand nombre des phéno-mènes collectifs observés dans les sociétés animales. Lorsque la taille du groupe est immense (certaines colonies de fourmis comportent plusieurs millions d’individus), lorsque l’espace à cou-vrir dépasse les capacités de communication des animaux (les troupeaux de gnous dans le parc du Serengeti peuvent s’étendre sur plusieurs kilomètres), lorsque la structure produite par le groupe nécessite le travail d’ouvriers sur plusieurs générations (c’est le cas du nid chez certaines espèces de termites) ou lorsque des individus qui ne se perçoivent pas tous doivent changer de
comportement à l’unisson (par exemple lors de changements brusques de direction dans les bancs de poissons), il devient difficile d’imaginer qu’un seul individu, ou même un petit groupe d’in-dividus, puisse mettre en place efficacement une quelconque forme d’organisation à une aussi grande échelle spatio-temporelle. L’extraordinaire quantité d’information à acquérir, à traiter et à restituer pour coordonner efficacement l’activité globale du groupe semble bien souvent hors de portée des capacités cognitives des animaux considérés, être humain compris (Seeley, 2002). La conclusion s’impose alors : une organisation collective, un ordre collectif peut émerger en l’absence de supervision.
Comprendre les mécanismes qui permettent l’émergence de ces propriétés à l’échelle du groupe revient à établir un lien entre deux niveaux d’organisation : d’un côté, le niveau “macroscopique” correspond au comportement du groupe lui-même, comme une entité propre réagissant aux sti-muli environnementaux ; de l’autre côté, le niveau “microscopique” correspond au comportement de chaque individu dans le groupe, en réaction à des stimuli locaux produits par l’environnement proche ou par les individus voisins et qui construisent une perception partielle de la structure ou du comportement du groupe auquel ils appartiennent. La nature du lien qui unit ces deux niveaux d’organisation a été étudié en premier lieu dans les systèmes physico-chimiques. Les tra-vaux d’Ilya Prigogine et de Grégoire Nicolis en particulier (Nicolis & Prigogine, 1977)ont conduit à la création d’une théorie puissante, la théorie de l’auto-organisation, qui permet d’expliquer l’émergence non supervisée (ou autonome) de structures, de régularités, à partir des interactions répétées entre les sous-unités d’un système.
Cette théorie a été transposée au cours des années 1970 à l’étude des systèmes biologiques, et en particulier à l’étude de l’organisation des sociétés d’insectes. Les travaux scientifiques menés depuis ont démontré que des processus d’auto-organisation sont à l’oeuvre dans de nombreux systèmes biologiques. Nous savons aujourd’hui qu’il n’est pas toujours nécessaire de superviser les activités de chaque membre d’un groupe pour que celui-ci s’organise et réponde aux défis de son environnement. C’est en réalité l’activité de chacun des individus combinée à celle de tous les autres selon des principes d’interaction souvent très simples qui fait émerger une organisation fonctionnelle à l’échelle de la société. Nous détaillerons plus loin la nature de ces principes ainsi que leurs propriétés. Lorsque cette organisation permet de résoudre des problèmes, elle peut être assimilée à une forme d’intelligence, qui apparaît seulement lorsque les individus sont en groupe. En référence à certaines espèces d’insectes sociaux (en particulier les guêpes et abeilles), cette intelligence “en groupe” est appelée intelligence “en essaim” (swarm intelligence en anglais).
Aujourd’hui, l’intelligence en essaim est un champ de recherche scientifique, multidisciplinaire, qui s’intéresse aux processus distribués (i.e. non supervisés) d’organisation et de résolution de problèmes présents dans un certain nombre de sociétés animales et dans des systèmes artificiels qui en sont inspirés. Le terme d’intelligence en essaim est apparu au cours de l’année 1989 dans des publications de Beni, Hackwood et Wang portant sur les systèmes de robotique cellulaire. Dans ces automates cellulaires, des agents virtuels représentant des robots se déplaçaient dans des environnements uni- ou bi-dimensionnels en interagissant avec leurs voisins proches. Chaque interaction entraînait une modification du comportement de l’agent selon des règles pré-établies. Selon le jeu de règles utilisé, des structures pouvaient éventuellement se former sans codage expli-cite ni supervision externe. Dans un article plus récent (Beni, 2005), Beni raconte que, lors d’une conférence en 1988 où il présentait ses travaux de robotique cellulaire (Beni, 1988), Alexander Meystel qualifia son système de “sort of swarm” (genre d’essaim). Trouvant l’expression accro-cheuse, il choisit de la conserver dans ses articles. Les robots cellulaires devinrent des essaims de robots, et l’intelligence développée par ses essaims devint l’intelligence en essaim. Par la suite en 1999, Bonabeau, Dorigo et Theraulaz étendirent le champ de l’intelligence en essaim à “toute tentative de conception d’algorithmes ou de dispositifs de résolution distribuée de problèmes ins-pirés par le comportement collectif des colonies d’insectes sociaux et des autres sociétés animales” (Bonabeau et al., 1999). Enfin, il y a quelques mois seulement, lors du lancement de la revue Swarm Intelligence1 (la première consacrée entièrement à l’étude de l’intelligence en essaim), son comité d’édition a ajouté au champ d’investigation l’ensemble des études expérimentales et théoriques portant sur les systèmes collectifs en biologie. Ironiquement, la source d’inspiration de la plupart des travaux dans ce domaine aura été la dernière a intégrer sa définition.
1.1.2 Auto-organisation et propriétés des systèmes d’intelligence en essaim Bien que cela ne soit pas explicitement spécifié dans sa définition, l’intelligence en essaim re-pose en grande partie sur des processus d’auto-organisation. Il existe de nombreuses définitions de l’auto-organisation dans la littérature. Celle que nous choisirons ici décrit l’auto-organisation comme l’émergence spontanée d’une organisation à l’échelle d’un système, à partir des interac-tions locales entre les sous-unités composant ce système, et sans codage explicite de la structure produite au niveau de ces sous-unités (Camazine et al., 2001). Les processus d’auto-organisation sont basés sur un ensemble d’éléments et de mécanismes dynamiques qui assurent l’apparition de la structure et sa stabilisation dans le temps. Dans les sections suivantes, nous décrirons
rapidement ces mécanismes, ainsi que les propriétés collectives générales qui en résultent.
1.1.2.1 Mécanismes
Les processus d’auto-organisation reposent en particulier sur les quatre constituants suivants (voir Figure 1.2) :
1. Rétroaction positive. Il y a rétroaction positive lorsque la probabilité d’apparition d’un élément augmente avec le nombre d’éléments similaires déjà présents. Ce mécanisme d’am-plification conduit à une augmentation non-linéaire de la quantité de ces éléments et sert de moteur à la création de nouvelles structures. Pour illustrer cette notion, prenons l’exemple d’une expérience portant sur l’attention collective et réalisée par Stanley Milgram en 1969 (Milgram & Toch, 1969). Cette expérience consiste à demander à des personnes dans des groupes de taille variable de fixer un point précis situé sur la façade d’un immeuble new-yorkais. Pendant ce temps, on relève le nombre de passants naïfs qui regardent dans cette direction. Les résultats obtenus montrent qu’une seule personne qui semble regarder un point d’intérêt entraîne en moyenne 40% des passants à regarder dans la même direction. Lorsque cinq personnes observent un même point, ce pourcentage double et il atteint près de 90% lorsque le groupe de démonstrateurs est composé de quinze personnes. Dans cette expérience, une rétroaction positive est en jeu : la probabilité que des personnes naïves regardent dans une direction précise augmente avec le nombre de personnes qui regardent déjà dans cette direction. Cette amplification conduit à une propagation rapide du com-portement de quelques individus dans la population.
2. Rétroaction négative. Il y a rétroaction négative lorsque la probabilité d’apparition d’un élément diminue avec le nombre d’éléments similaires déjà présents. Ce mécanisme d’atté-nuation s’oppose à la rétroaction positive dans les sytèmes auto-organisés, et conduit à une stabilisation de la structure collective lorsqu’un équilibre dynamique entre les deux types de rétroactions est atteint. Pour illustrer ce propos, prenons l’exemple du recrutement ali-mentaire chez l’abeille domestique Apis mellifera (Seeley et al., 1991). Chez cette espèce, une ouvrière qui découvre une source de nourriture rentre à sa ruche et entame à l’intérieur une danse de recrutement. Cette danse, permet à l’abeille de communiquer la direction, la distance et la qualité de la source de nourriture découverte aux ouvrières qui se trouvent autour d’elle. Certaines d’entre elles prendront alors leur envol pour aller exploiter cette source. A leur retour, elles entameront éventuellement une danse similaire afin de recruter de nouvelles butineuses. Le recrutement par danse des abeilles est un exemple classique de
rétroaction positive, une abeille en recrutant plusieurs qui à leur tour en recrutent plusieurs autres, etc. En l’absence de toute régulation, ce recrutement conduirait inévitablement au regroupement de toutes les abeilles de la ruche sur la source de nourriture découverte. Dans la nature, cela n’arrive jamais car la rétroaction positive est contrebalancée par un certain nombre de mécanismes réalisant une rétroaction négative : nombre limité d’ouvrières recru-tables pour le fourragement alimentaire, épuisement de la source de nourriture, compétition entre des sources de nourriture, etc. Lorsque la rétroaction positive et la rétroaction néga-tive s’équilibrent, cela conduit à une stabilisation du nombre d’abeilles exploitant la source de nourriture. En général, les boucles de rétroaction négative sont plutôt associées à des facteurs non comportementaux (contraintes spatiales, évaporation du signal, etc.) alors que les boucles de rétroaction positive sont plutôt associées au comportement de l’animal, 3. Fluctuations. Ce sont des hétérogénéités qui modifient le comportement des individus.
Lorsque ces modifications sont soumises à un mécanisme de rétroaction positive, elles peuvent être amplifiées et donner naissance à une structure, qui sera éventuellement sta-bilisée par un mécanisme de rétroaction négative. Ces fluctuations peuvent être des hé-térogénéités environnementales qui vont alors favoriser l’émergence d’une structure à un endroit plutôt qu’à un autre. Mais elles peuvent également être dues à la variabilité des comportements individuels. Dans ce cas, une structure peut être initiée et amplifiée dans un environnement parfaitement homogène.
4. Interactions. Deux individus interagissent lorsqu’au moins l’un d’eux est en mesure d’acqué-rir de l’information détenue par l’autre et que cette information entraîne une modification de l’état de l’individu qui la reçoit. Si cette modification a pour effet d’augmenter (respecti-vement de diminuer) la probabilité d’apparition d’un état particulier, alors la répétition de l’interaction à l’échelle de la population devient le support d’un mécanisme de rétroaction positive (respectivement négative). La multiplication de ce type d’interactions est donc à la base des processus d’auto-organisation.
1.1.2.2 Propriétés
De ces quatre constituants de base découlent plusieurs de propriétés qui caractérisent les processus d’auto-organisation. En particulier, on distingue classiquement les cinq propriétés sui-vantes :
1. Ce sont des systèmes dynamiques. La mise en place d’une organisation ainsi que son main-tien dans le temps sont le fruit des interactions permanentes entre les individus qui
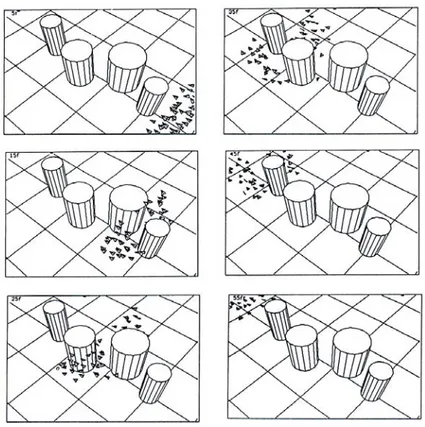
com-Figure 1.2: Illustration des constituants de l’auto-organisation à travers l’exemple du recrute-ment alirecrute-mentaire chez la fourmi. Au début de l’expérience, les fourmis découvrent par hasard une source de nourriture. Le nombre de fourmis à cette source est alors très faible et fluctue de manière aléatoire (flèche verte). Lorsqu’elles retournent à leur nid, ces fourmis déposent derrière elles une piste chimique qui leur permet d’interagir avec les autres ouvrières. En particulier, cette piste stimule les autres ouvrières à rechercher de la nourriture et les guide vers la source découverte. Ces fourmis recrutées vont à leur tour déposer une trace chimique par-dessus la pre-mière piste à leur retour vers le nid. Elles amplifient ainsi le pouvoir attracteur de la piste. Cette rétroaction positive (flèche rouge) enclenche une croissance exponentielle du nombre de fourmis présentes à la source. Enfin, l’évaporation de la piste chimique, sa saturation, l’encombrement à la source ou sur la piste vont limiter le nombre maximal de fourmis qui pourront de rendre à la source. Cette rétroaction négative (flèche bleue) va contrebalancer l’effet de la rétroaction positive, et le nombre de fourmis à la source va se stabiliser en fin d’expérience.
posent le groupe. En conséquence, l’état du système évolue au cours du temps et/ou en réponse à des perturbations. Le système peut éventuellement atteindre un état stable, mais celui-ci ne sera pas figé. Au contraire, il s’agira d’un équilibre dynamique qui résultera de la compétition ininterrompue entre les rétroactions positives et négatives.
2. Ce sont des systèmes redondants. A l’intérieur du groupe, plusieurs individus (voire tous) peuvent accomplir une même tâche. En conséquence, l’échec d’un individu dans la réalisa-tion d’une tâche ou la dispariréalisa-tion de cet individu du système peut être compensée par les autres membres du groupe. Cette redondance augmente la robustesse générale du système aux perturbations et aux variations de l’environnement.
3. Ces systèmes présentent des propriétés émergentes. Le comportement du groupe ne peut être résumé à la simple addition des contributions de chaque membre du groupe. Il est le résultat d’une combinaison non linéaire de ces contributions, grâce aux interactions entre les membres du groupe qui supportent des processus de rétroaction positive. La conséquence est l’apparition à l’échelle du groupe de nouvelles propriétés qui ne sont pas directement déductibles du comportement isolé de chaque individu.
4. Le comportement de ces systèmes présente des bifurcations. Lorsque certains paramètres du système évoluent, son comportement peut opérer une transition d’un état stable vers un autre. Il s’agit en fait d’un changement qualitatif du comportement du groupe. Par exemple, lors de la construction de leur nid, les guêpes sociales Polistinae doivent prendre en charge trois activités différentes : récolter de la pulpe de bois, récolter de l’eau et construire les cellules du nid. Lorsque la taille de la colonie est faible, chaque guêpe peut prendre en charge chacune de ces activités indifféremment. Mais si la taille de la colonie dépasse un certaine valeur seuil, alors il apparaît des spécialistes qui prennent en charge exclusivement l’une de ces tâches (Karsai & Wenzel, 1998; Theraulaz et al., 1998). La taille de la colonie est donc un paramètre susceptible de provoquer un changement qualitatif (i.e., une bifurcation) dans l’organisation de la colonie.
5. Enfin, ces systèmes peuvent être multi-stables. Pour des paramètres donnés, le système peut atteindre plusieurs états stables différents, Cela signifie que qu’un groupe donné peut présenter plusieurs comportements collectifs différents, en fonction des conditions initiales ou de l’historique des fluctuations aléatoires.
Figure 1.3: Les quatres étapes de l’étude des processus d’auto-organisation.
1.1.2.3 Etude des systèmes d’intelligence en essaim
Pour comprendre l’émergence d’un phénomène d’auto-organisation, il faut avant tout établir un lien causal entre le comportement des individus et celui du groupe auquel ils appartiennent. Une approche en quatre étapes permet l’établissement de ce lien (voir Figure 1.3).
1. Dans la première étape, l’étude porte sur la structure et la dynamique du groupe (i.e., l’évolution de la structure au cours du temps). Cette étape permet de caractériser, d’une part, l’état d’organisation du groupe, et d’autre part, la ou les fonctions remplies par cette organisation particulière. Dans le premier cas, il s’agit d’identifier la présence d’élé-ments récurrents (on parle également de patterns), non aléatoires, et qui sont le signe d’une organisation sous-jacente. Dans le second cas, il s’agit de comprendre quel défi de l’environ-nement est solutionné par cette organisation. Il est également fréquent d’étudier l’impact de différentes conditions environnementales sur la structure et la dynamique du groupe. 2. Dans une deuxième étape, l’étude s’attache à identifier dans le comportement des individus
composant le groupe et dans leurs interactions des mécanismes susceptibles d’expliquer la dynamique observée à l’échelle du groupe. Il s’agit dans cette étape de caractériser finement les comportements des individus en fonction des conditions environnementales et sociales qu’ils perçoivent. En particulier, l’influence des congénères sur le comportement individuel doit faire l’objet d’une analyse précise. Par exemple, le degré de mimétisme comportemental des individus aura un impact très important sur la capacité du groupe à
opérer des choix collectifs. En général, le comportement des individus est décrit dans cette étape par des processus stochastiques, comme par exemple des probabilités de transition entre deux comportements donnés. Ce type de description permet à la fois de capturer le changement de comportement et la variabilité qui y est associé.
3. La troisième étape consiste à rassembler dans un cadre cohérent les observations obte-nues au cours de l’étape précédente. Il s’agit de bâtir un modèle contenant l’ensemble des comportements individuels susceptibles d’expliquer l’apparition d’une organisation fonc-tionnelle (car elle répond à un problème) à l’échelle du groupe. Ce modèle peut être basé sur des observations qualitatives des comportements individuels. Dans la mesure du pos-sible, il est cependant préférable de disposer d’une mesure quantitative de la valeur de chaque paramètre du modèle.
4. Enfin, la quatrième étape consiste à formaliser le modèle dans un langage permettant de l’implémenter sur une plateforme de simulation. Ce langage est en général celui des équa-tions mathématiques ou des algorithmes informatiques. La simulation du modèle permet d’observer la structure et la dynamique collective théoriques obtenues à partir des éléments individuels décrits lors de l’étape numéro 2. La comparaison de ces observations théoriques aux observations réalisées lors de l’étape numéro 1 permet de confirmer ou d’infirmer la validité du modèle proposé dans l’étape numéro 3. La simulation du modèle est donc une étape cruciale pour établir le lien entre le niveau des individus et celui du groupe. Elle per-met en outre de faire des prédictions sur de nouvelles conditions qui pourront être ensuite testées expérimentalement.
1.1.3 Interactions et transferts d’information dans les systèmes d’intelligence en essaim
Les propriétés d’auto-organisation d’un système reposent principalement sur les interactions entre ses composants. Classiquement, les interactions au sein des systèmes auto-organisés sont divisées en deux catégories distinctes, qui représentent deux manières différentes pour les éléments d’un système ou les individus d’un groupe d’échanger de l’information.
1.1.3.1 Transfert indirect d’information
Le transfert indirect d’information est basé sur une modification par un individu de l’envi-ronnement dans lequel il vit. Cette modification peut en retour modifier le comportement de ce même individu ou de ses congénères lorsqu’ils la perçoivent. Ce type d’interactions reposant sur
la formation d’une trace dans l’environnement est qualifié de stigmergique (Theraulaz & Bona-beau, 1999). Le concept de stigmergie a été introduit pour la première fois à la fin des années 1950 par le biologiste Pierre-Paul Grassé pour rendre compte de la coordination des activités lors de la construction du nid chez les termites (Grassé, 1959). Grassé montra que chaque acte de construction réalisé par les ouvriers était déterminé par la structure locale de l’environnement qui est elle-même modifiée par l’acte de construction. A travers cette boucle, les traces laissées par les actions présentes de chaque individu influencent le comportement futur des individus qui les percevront.
La construction du nid chez les guêpes sociales Polistinae est un bon exemple de compor-tement basé sur un transfert stigmergique de l’information (voir Figure 1.5a). Chez ces guêpes, la grande majorité des nids est constituée de rayons de cellules hexagonales construites à par-tir de fibres végétales et de sécrétions orales assemblées par mastication (Wenzel, 1991). Afin de déterminer l’emplacement d’une nouvelle cellule, ces insectes utilisent la configuration locale des cellules qui les entourent et qu’ils perçoivent grâce à leurs antennes (Karsai & Theraulaz, 1995). En fonction de cette configuration, la probabilité de construire une cellule varie fortement. Par exemple, elle est élevée lorsque le site potentiel de construction est entouré de trois murs adjacents. Elle est au contraire très faible lorsqu’il n’existe qu’un ou deux murs adjacents. Les constructions passées et leur résultat “stocké” dans l’environnement ont donc une influence sur les actes de construction suivants. Ils permettent ainsi une coordination précise des activités indi-viduelles. Dans ce cas c’est l’architecture qui contrôle indirectement de déroulement des activités bâtisseuses.
Le transfert stigmergique d’information n’est pas uniquement porté par des modifications qualitatives des structures présentes dans l’environnement. Toute trace durable déposée dans l’environnement et qui peut être détéectée peut suffire à établir une interaction stigmergique entre des individus. Ainsi, de nombreuses espèces de fourmis communiquent entre elles au moyen d’un signal chimique, une phéromone, qu’elles déposent dans l’environnement afin de marquer, par exemple, le chemin qu’elles ont emprunté pour rejoindre une source de nourriture depuis leur nid (Hölldobler & Wilson, 1990). Ce signal chimique a la particularité d’être attractif pour les fourmis passant à proximité. La piste chimique déposée par une première fourmi va alors augmenter la probabilité que d’autres ouvrières empruntent le même chemin, et éventuellement renforcent le marquage de la piste par leurs propres dépôts chimiques, augmentant d’autant plus son attractivité. On retrouve donc ici aussi des interactions stigmergiques qui vont induire un feedback positif. A long terme, le renforcement de cette trace phéromonale par les dépôts
successifs des fourmis peut conduire à l’établissement d’un trafic important d’ouvrières entre leur nid et une source de nourriture, comme en par exemple les gigantesques colonnes de chasse des fourmis légionnaires qui peuvent parfois s’étendre sur plusieurs centaines de mètres (Franks & Fletcher, 1983; Franks, 1989; Hölldobler & Wilson, 1990).
Enfin, des interactions stigmergiques peuvent également avoir lieu dans des environnements sans dimension physique. Dans certaines communautés internet, par exemple sur le site Digg.com, l’exposition d’une nouvelle sur la page d’entrée du site est directement reliée à sa popularité (Wu & Huberman, 2007). Chaque membre de cette communauté peut attribuer un point à chaque nouvelle qu’il juge intéressante. Plus une nouvelle récolte de points dans un laps de temps donné, plus elle est mise en avant sur la page d’entrée du site. Cette mise en avant augmente la probabilité que la nouvelle atteigne d’autres membres naïfs de la communauté. Ces membres peuvent à leur tour, à la manière des dépôts chimiques chez les fourmis, renforcer la nouvelle en lui attribuant un nouveau point. Des interactions stigmergiques similaires sont également utilisées dans les réseaux sociaux pour assurer la promotion de certains produits (Leskovec, 2006).
1.1.3.2 Transfert direct d’information
Contrairement aux interactions stigmergiques, les transferts directs d’information ne se font pas par l’intermédiaire d’une trace durable laissée dans l’environnement. Dans le cas d’interac-tions directes, les individus tirent leurs informad’interac-tions de l’observation du comportement de leurs congénères voisins (voir Figure 1.5b). L’information peut circuler sous plusieurs formes (visuelle, tactile, auditive), mais elle ne perdure pas dans l’environnement.
De nombreux comportements auto-organisés reposent sur des interactions directes entre les membres d’un groupe. Probablement l’exemple le plus parlant est celui des bancs de poissons et des vols d’oiseaux dans lesquels des milliers, voire même des millions, d’individus se déplacent de manière cohérente, changeant de direction parfois brusquement mais toujours de manière synchrone (Partridge, 1982; Aoki, 1982; Parrish et al., 2002; Ballerini et al., 2008). Cette cohésion presque parfaite est le résultat d’un fort mimétisme comportemental couplé à un transfert direct d’information : chaque poisson, chaque oiseau adopte dans ces groupes la direction et la vitesse de déplacement de ses voisins. De proche en proche, chaque individu s’aligne sur ses voisins qui eux-mêmes s’alignent sur les leurs : le groupe adopte ainsi une direction commune de déplacement sans concertation de ses membres ni supervision par l’un d’entre eux. Comme l’information “direction de déplacement” n’est pas imprimée dans l’environnement, toute variation importante dans le déplacement de quelques individus en réponse à des facteurs externes au groupe va se
Figure 1.4: A gauche, de 1 à 8, une séquence d’images montrant l’attaque d’un prédateur sur un banc de poissons. A droite, de a à h, un banc de poissons simulés (n=1000) répondant à l’attaque d’un prédateur simulé.
propager rapidement à l’ensemble du groupe et entraîner un brusque changement de direction ou de configuration du groupe. Cela arrive par exemple lorsque qu’un banc de poissons est attaqué par un prédateur. Les individus qui le perçoivent et tentent de le fuir déclenchent alors des vagues de mouvements de fuite qui parcourent l’ensemble du groupe.
Chez l’homme, on retrouve un phénomène similaire lorsque, à la fin d’un concert par exemple, la foule qui applaudie au départ de manière désordonnée adopte peu à peu un battement ryth-mique (Néda et al., 2000; Strogatz, 2003). Grâce à un signal acoustique, chaque spectateur communique à ses voisins son propre rythme de battement et perçoit celui des personnes qui l’entourent. Comme chez oiseaux ou les poissons, les individus ajustent leur rythme sur celui de leurs voisins et la foule finit par adopter une fréquence d’applaudissement consensuelle. Toujours sur le même principe, certaines espèces de lucioles sont capables de synchroniser leurs émissions lumineuses en adaptant leur fréquence propre d’émission sur celle de leur voisins (Buck & Buck, 1976).
Figure1.5: Deux exemples d’interactions au sein de sociétés animales. (a) Lors de la construction de leur nid, les guêpes Polistes interagissent indirectement à travers la configuration des cellules du nid en cosntruction. (b) Chez l’abeille mellifère, une ouvrière qui a découvert une source de nourriture informe directement ces congénères à travers une danse qui leur indique la direction et la distance à laquelle se trouve la source.
Enfin, des interactions directes à courte distance sont à l’origine de l’organisation particulière du trafic dans les colonnes de chasse des fourmis légionnaires (Couzin & Franks, 2003). Lorsque deux fourmis se rencontrent sur ces pistes, elles se perçoivent à courte distance et amorcent une manoeuvre d’évitement. Cependant, chaque fourmi ne s’écarte pas de sa trajectoire de la même façon, selon qu’elle provient du front de chasse ou du bivouac de la colonie. En particulier, les fourmis provenant du bivouac tournent de manière plus prononcée que les fourmis provenant du front. A chaque interaction, les premières ont une probabilité plus importante de se déporter vers le bord de la piste. La répétition de ce type d’interaction directe à l’échelle de la colonie conduit à une organisation du trafic selon trois flux distincts de fourmis (voir Figure ??b) : un flux central composé majoritairement de fourmis provenant du front de chasse, flanqué de deux flux latéraux composés majoritairement de fourmis provenant du bivouac (Franks, 1985).
1.1.3.3 Direct vs indirect
Les interactions directes et indirectes (ou stigmergiques) peuvent cohabiter au sein des so-ciétés animales. Cependant les spécificités de ces deux formes de transferts d’information leur confèrent des propriétés différentes à l’intérieur des processus d’auto-organisation. Lors de trans-ferts indirects d’information, la trace laissée par les individus confère une forme de stabilité temporelle à la solution trouvée par le groupe. Une fois que cette dernière est établie, elle reste “imprimée” dans l’environnement pendant une durée qui peut parfois être très longue. Certaines espèces de fourmis des bois (Rosengren & Sundström, 1987; Chauvin, 1962), de fourmis cou-peuses de feuilles (Weber, 1972; Shepherd, 1982; Vasconcelos, 1990) et de fourmis granivores (Hölldobler, 1976; Lopez et al., 1994; Detrain et al., 2000; Azcárate & Peco, 2003) forment des
pistes qui peuvent durer plusieurs semaines à plusieurs mois, et peuvent même résister jusqu’à la fin de la période d’hibernation. De plus, les ouvrières nettoient la piste des débris et de la vé-gétation qui s’y trouvent, et peuvent dans certains cas construire des murs ou un tunnel autour (Shepherd, 1982; Kenne & Dejean, 1999; Anderson & Mcshea, 2001).
Dans ces exemples, les pistes conduisent à des ressources persistantes dans l’environnement et leur stabilité maintient ainsi la mémoire du chemin vers ces ressources. Les interactions indirectes sont donc le plus souvent associées à des solutions qui présentent une certaine robustesse aux perturbations. Cependant, cette robustesse implique parfois une moindre flexibilité. Par exemple, une fois que la piste vers une ressource est établie, il est difficile pour certaines espèces de fourmis d’abandonner ensuite cette piste pour un chemin plus court ou une ressource de meilleure qualité (Beckers et al., 1990). Pour être plus précis, il existe une forme de compromis entre la robustesse et la flexibilité de la structure produite par le processus d’auto-organisation. Et la valeur de ce compromis est régulée par la stabilité de la trace laissée par les individus. Par exemple, les piliers, les murs et les plafonds construits par les termites sont des traces très stables qui assurent la pérennité du nid pour plusieurs générations, même si les dimensions ou l’organisation de certaines structures comme la chambre royale peuvent être adaptées au cours de la vie d’une colonie (Bruinsma, 1979). Au contraire, les traces virtuelles déposées dans les communautés Internet sont plus volatiles, l’intérêt des utilisateurs pour une information diminuant rapidement dans le temps. Cela assure le renouvellement quotidien des sujets traités sur le réseau.
Les interactions directes permettent plutôt de créer des structures hautement réactives aux perturbations. Comme nous l’avons évoqué un peu plus haut, les bancs de poissons ou les vols d’oiseaux peuvent modifier leur déplacement de manière brusque lorsqu’un prédateur se présente. La propagation de l’information de proche en proche permet à celle-ci de traverser à grande vitesse l’ensemble du groupe qui peut en conséquence réagir très rapidement. Cependant, cette grande flexibilité peut également avoir des conséquences négatives. Par exemple, une fluctuation aléatoire dans le comportement des individus peut soudainement être amplifiée et créer ainsi des déplacements inutiles et coûteux. Un compromis entre la flexibilité et la stabilité du groupe existe donc également dans le cas des processus d’auto-organisation basés sur des interactions directes. La valeur de ce compromis est en général régulée par le nombre de congénères que l’individu prend en compte pour ajuster son comportement. Plus ce nombre est élevé, moins les fluctuations aléatoires locales auront d’impact sur la structure du groupe.
Figure1.6: Exemple de coordination dans les systèmes d’intelligence en essaim. Dans ce banc de poisson, les individus coordonnent leur déplacement afin de se diriger dans une même direction.
1.1.4 Fonctions assurées par les systèmes d’intelligence en essaim
Dans un contexte biologique, les processus d’auto-organisation remplissent souvent des fonc-tions qui permettent au groupe ou à la société animale d’appréhender les divers éléments de son environnement. Il est possible de regrouper ces fonctions au sein de trois grandes catégories, non exclusives l’une de l’autre et souvent combinées au sein d’un même comportement collectif. Ces trois catégories sont la coordination, la collaboration et la délibération.
1.1.4.1 Coordination
La coordination correspond à l’organisation dans l’espace et dans le temps des tâches requises pour résoudre un problème (Figure 1.6). Elle affecte la séquence temporelle et/ou la répartition spatiale des activités des individus. Par exemple, dans les essaims d’abeilles (Janson et al., 2005) et de criquets (Buhl et al., 2006b), dans les vols d’oiseaux (Ballerini et al., 2008)ou les bancs de poissons (Aoki, 1982), les individus alignent leur déplacement sur celui de leurs voisins. Ce comportement génère une synchronisation des changements individuels de direction (coordina-tion temporelle) et l’adop(coordina-tion d’une direc(coordina-tion commune de déplacement (coordina(coordina-tion spatiale). D’une manière générale, la coordination de l’activité des individus est la conséquence princi-pale des processus d’auto-organisation qui sont au coeur des systèmes d’intelligence en essaim. Les mécanismes de rétroaction positive, en favorisant la réalisation d’une tâche à un endroit donné ou en propageant un comportement dans la population, conduisent nécessairement à une organisation spatio-temporelle des activités des individus.
Figure 1.7: Exemple de collaboration dans les systèmes d’intelligence en essaim. Lors de la construction du nid chez la fourmi tisserande Oecophylla longinoda, un premier groupe de fourmis assure la mise en place des feuilles, pendant qu’un second groupe les connecte à l’aide la soie produite par les larves.
1.1.4.2 Collaboration
La collaboration correspond à la répartition adéquate des activités entre des individus ou des groupes d’individus spécialisés dans la réalisation de l’une de ces activités (Figure 1.7). On parle également de division du travail. Cette répartition des tâches peut s’établir sur la base de différences morphologiques inter-individuelles : des individus aux capacités physiques différentes vont accomplir des tâches différentes. Un exemple bien connu est l’existence de castes physiques chez certaines espèces d’insectes sociaux. Chez les fourmis Atta coupeuses de feuilles par exemple, la taille des ouvrières est fortement corrélée aux tâches qu’elles effectuent (Hölldobler & Wilson, 1990). Seules les ouvrières dont la tête dépasse 1,6 millimètres sont capables de couper les feuilles qui sont utilisées pour faire pousser le champignon qui sert de nourriture principale à la colonie. Cependant, la tête de ces ouvrières est trop grosse pour se glisser dans les galeries où pousse ce champignon. Seules les ouvrières dont la tête à une largeur d’environ 0,5 millimètres en sont capables et prennent en charge la culture du champignon.
La spécialisation des individus peut également être le fruit de leur expérience (Deneubourg
et al., 1987; Bonabeau et al., 1998b). Des individus régulièrement confrontés à un stimulus
par-ticulier déclenchant la réalisation d’une tâche particulière peuvent devenir plus sensibles à la présence de ce stimulus. Une stimulation moins importante suffira alors à déclencher la réalisa-tion de la tâche. Tant que l’intensité du stimulus reste suffisamment faible, seuls ces individus “spécialistes” accompliront la tâche associée. Ce mécanisme basé sur l’expérience des individus semble être à l’origine de l’organisation du travail chez plusieurs espèces d’insectes sociaux, et en particulier chez certaines guêpes sociales (Gadagkar & Joshi, 1983, 1984) et certaines espèces de fourmis (Ravary et al., 2007).
Figure1.8: Exemple de délibération dans les systèmes d’intelligence en essaim. Les fourmis ont accès depuis leur nid à une source de nourriture de l’autre côté de ce pont dont les deux branches son de longueur identique. Elles sélectionnent cependant l’une des branches sur laquelle elles établissent la majorité de leur trafic.
1.1.4.3 Délibération
La délibération correspond à la sélection d’une alternative parmi plusieurs possibilité offerte au système (Figure 1.8). On parle également de choix collectif ou de décision collective. Par exemple, l’abeille mellifère Apis mellifera est capable de sélectionner la parcelle florale la plus productive grâce au recrutement d’ouvrières inactives (Seeley et al., 1991). La fourmi Lasius
niger est elle aussi capable de choisir parmi plusieurs sources de nourritures celle qui est la plus
riche, et parmi plusieurs chemins conduisant à une source, celui qui est le plus court (Beckers
et al., 1990, 1992b).
Cette fonction faisant l’objet du présent travail, elle sera traitée en détails dans la section 1.3 de ce mémoire.
1.2 Intelligence en essaim dans les systèmes artificiels
La simplicité des principes d’organisation à l’oeuvre au sein des sociétés animales couplée à l’importante gamme de problèmes qu’ils sont capables de résoudre efficacement ont attiré depuis maintenant plus de 30 ans l’attention d’une communauté grandissante de chercheurs et d’ingénieurs en recherche opérationnelle, en informatique ou encore en robotique. Dans cette section, nous allons plus particulièrement nous intéresser à l’application de l’intelligence en essaim dans les systèmes multi-robots. Avant cela, il nous semble nécessaire de faire quelques rappels historiques qui permettront de mieux positionner la robotique en essaim dans le champ plus large