Onocerane testifies to dry climatic events during the Quaternary in the Tropics.
Texte intégral
Figure

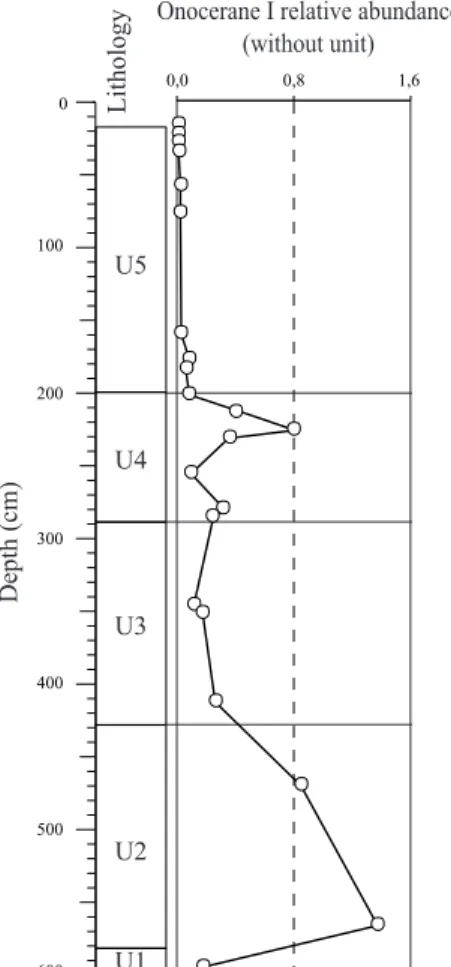
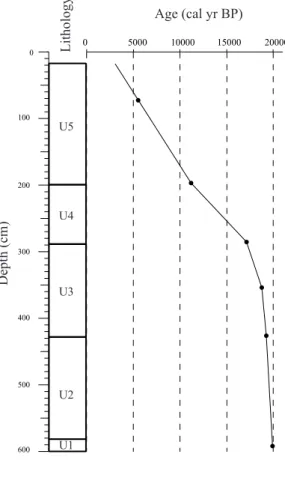
Documents relatifs
In the present research work, we covalently grafted ionenes onto different surfaces using a robust and efficient method based on polydopa- mine coating and
يديهمتلا بناجلا ثحبلاب فيرعتلا 7 ةعبارلا ةساردلا : دمحا يديس كوشتوكل ةسارد ،كوشتوك( 0270 ) اونعب :ن حرتقم يبيردت جمانرب ريثأت
It is therefore reasonable to fit the fluorescence intensity versus time data for linear sweep iDEP in terms of a minimum voltage at which trapping appears and the rate at which
The device utilizes dielectrophoresis (DEP) to simultaneously sense and sort individual cells based on their internal lipid content. Microorganisms such as yeast,
Quantifying the impact of soil texture on plant growth, grain yield and water productivity under AWD and contrasting seedling age: farmers’ rice fields in Tarlac region, dry
variegatum: the tick infestation rate on animals, the tick infection rate, the vector efficacy, the daily inoculation rate, additional pathoge- nicity induced by ticks, the delay of
More preserved left language network in patient with signs of covert command following as compared to patients with MCS-. One patient (8%) with covert signs of
