Flowering phenology and reproductive fitness along a mountain slope: maladaptive responses to transplantation to a warmer climate in Campanula thyrsoides
Texte intégral
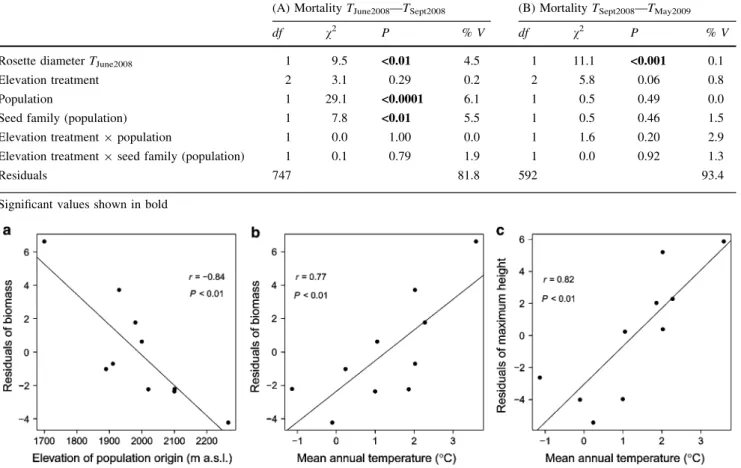
Figure


Documents relatifs
I manipulated density of pool frogs (Rana lessonae) during the aquatic larval and the terrestrial juvenile stages and examined the effect on growth and survival until 1 year of
We study the shortest-path broadcast problem in graphs and digraphs, where a message has to be transmitted from a source node s to all the nodes along shortest paths, in the
A case study based on the comparison of Micro- geomorphology study and numerical simulation of Chui-Sue river watershed suggested that with proper parameters, the numerical model
Conclusions The results of characterization of genetic diversity may reflect different selection criteria for different uses over. centuries of
TABLE 5 | Segmentations of series of mean temperatures during the HA period using piecewise constant models (2 or 1 segment when the 2-segment model was irrelevant): recording
Abstract We investigated long-term (over 100 years) tree-ring width (TRW) variabilities as well as short-term (10 years) variations in net ecosystem productivity (NEP) in response
INRA, Institut de Recherche en Horticulture et Semences (IRHS), UMR 1345 SFR 4207 QUASAV, Beaucouzé, France a ; Université d’Angers, Institut de Recherche en.. Horticulture et
(a) Dynamics of caspase-specific activity rates (pmol L −1 mg protein −1 min −1 ) of Trichodesmium in the New Caledonian lagoon during bloom accumulation and bloom demise,
